

Histoplasma capsulatum Activates Hematopoietic Stem Cells and Their Progenitors through a Mechanism Dependent on TLR2, TLR4, and Dectin-1


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Considerations
2.2. Isolation, Purification, and Maintenance of Bone Marrow-Derived HSPCs
2.3. Determination of the TLR2, TLR4, and Dectin-1 Expression on HSPC
2.4. Histoplasma Capsulatum Yeasts
2.5. Culture of HSPC with H. capsulatum Yeasts
2.6. Phagocytosis Assay
2.7. Microbicidal Activity of HSPCs against H. capsulatum Yeasts
2.8. Determination of the Expression of Cytokines and Inflammatory Mediators by Real-Time Quantitative PCR (qPCR)
2.9. Apoptosis Assay
2.10. Cell Proliferation Assay
2.11. Statistical Analysis
3. Results
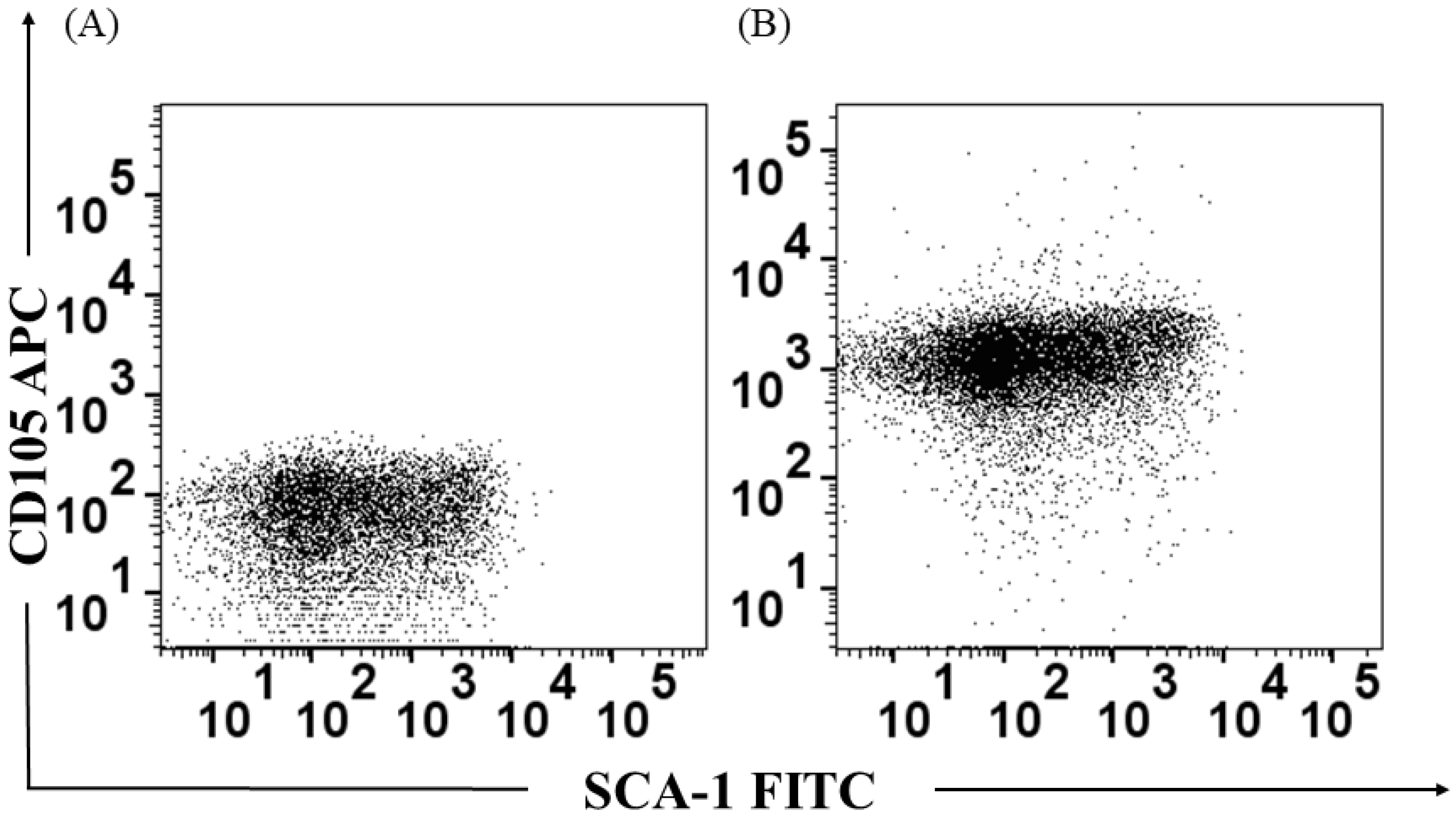
3.1. Identification of the Bone Marrow-Derived HSPC Population
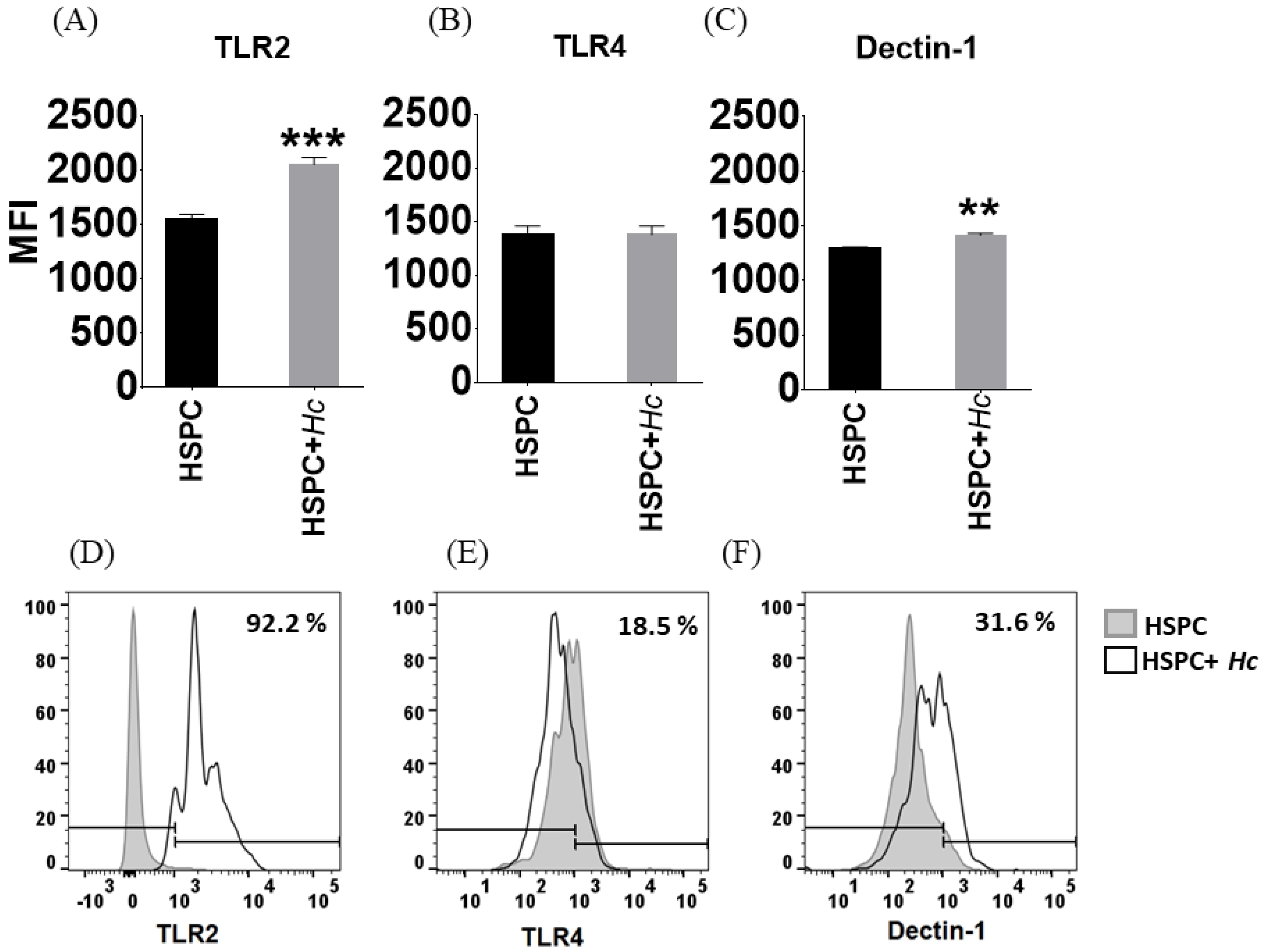
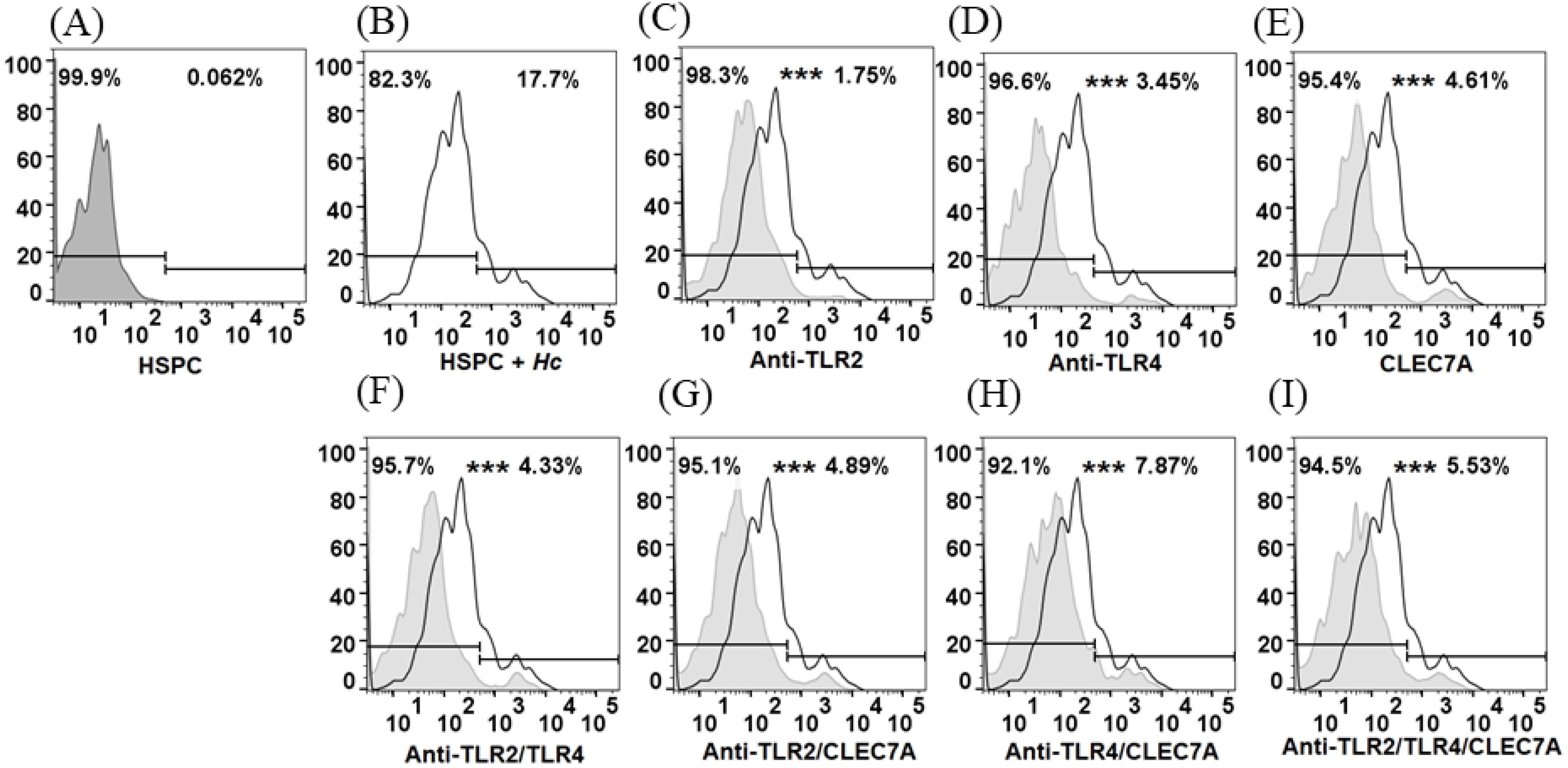
3.2. Histoplasma Capsulatum induces Increased Expression of TLR2 and Dectin-1 in HSPC
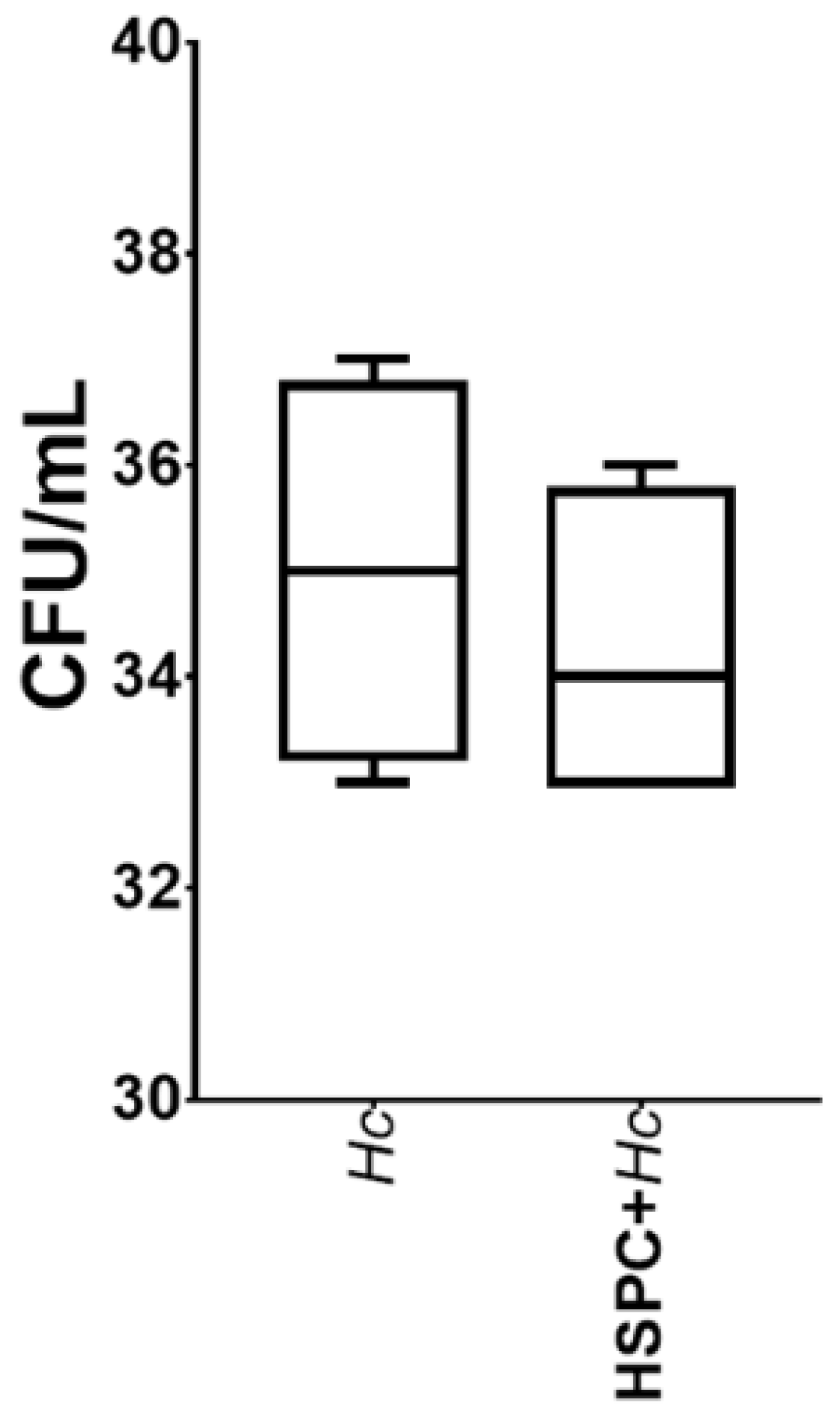
3.3. HSPCs Are Able to Phagocytose H. capsulatum Yeasts but Do Not Exert a Fungicidal Effect
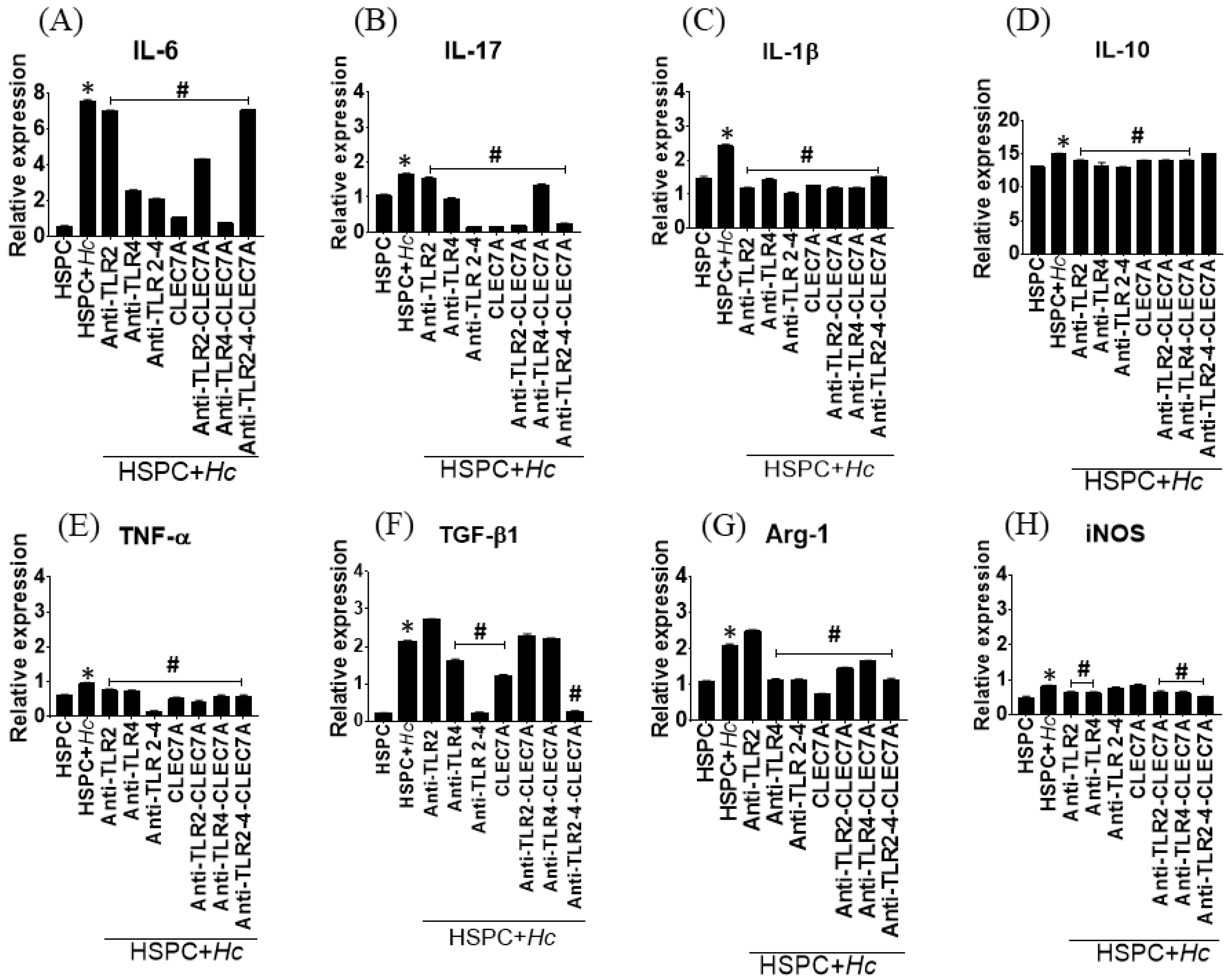
3.4. Histoplasma Capsulatum induces the Expression of Cytokines and Inflammatory Mediators in HSPCs
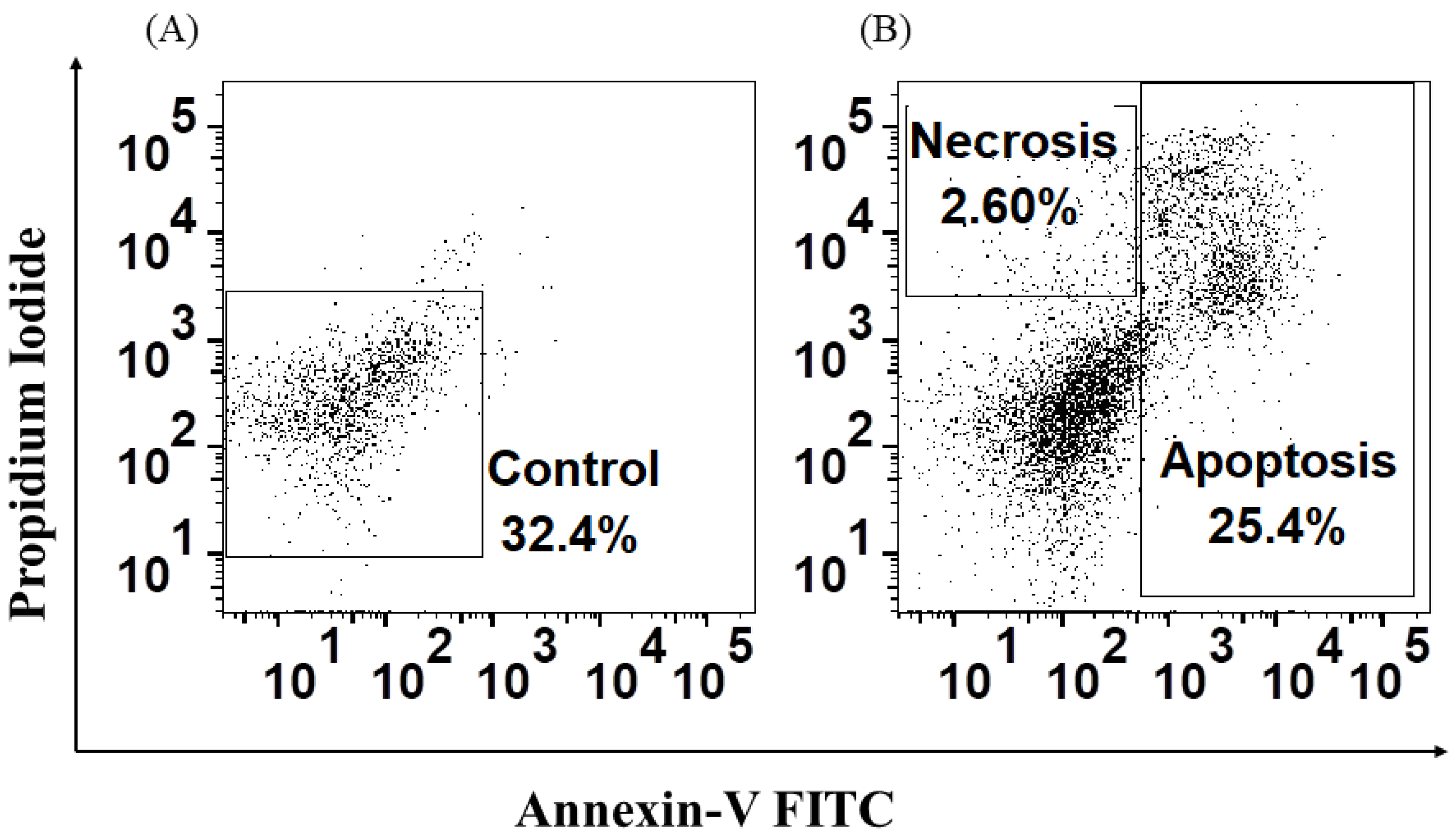
3.5. H. capsulatum Yeasts Induce Apoptosis and Necrosis and Affect Proliferation in HSPCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rossi, L.; Challen, G.A.; Sirin, O.; Lin, K.K.-Y.; Goodell, M.A. Hematopoietic stem cell characterization and isolation. Methods Mol. Biol. 2011, 750, 47–59. [Google Scholar]
- Dzierzak, E.; Bigas, A. Blood Development: Hematopoietic Stem Cell Dependence and Independence. Cell Stem Cell 2018, 22, 639–651. [Google Scholar]
- Domen, J. Hematopoietic Stem Cells: Isolation, Characterization and Applications. In Biomedical Sciences: Encyclopedia of Tissue Engineering and Regenerative Medicine; Academic Press: Cambridge, MA, USA, 2019; pp. 15–24. [Google Scholar]
- Yáñez, A.; Goodridge, H.S.; Gozalbo, D.; Gil, M.L. TLRs control hematopoiesis during infection. Eur. J. Immunol. 2013, 43, 2526–2533. [Google Scholar]
- Sallustio, F.; Curci, C.; Stasi, A.; De Palma, G.; Divella, C.; Gramignoli, R.; Castellano, G.; Gallone, A.; Gesualdo, L. Role of Toll-Like Receptors in Actuating Stem/Progenitor Cell Repair Mechanisms: Different Functions in Different Cells. Stem Cells Int. 2019, 2019, 6795845. [Google Scholar]
- Megías, J.; Martínez, A.; Yáñez, A.; Goodridge, H.S.; Gozalbo, D.; Gil, M.L. TLR2, TLR4 and Dectin-1 signalling in hematopoietic stem and progenitor cells determines the antifungal phenotype of the macrophages they produce. Microbes Infect. 2016, 18, 354–363. [Google Scholar]
- Nagai, Y.; Garrett, K.P.; Ohta, S.; Bahrun, U.; Kouro, T.; Akira, S.; Takatsu, K.; Kincade, P.W. Toll-like receptors on hematopoietic progenitor cells stimulate innate immune system replenishment. Immunity 2006, 24, 801–812. [Google Scholar]
- Kobayashi, H.; Suda, T.; Takubo, K. How hematopoietic stem/progenitors and their niche sense and respond to infectious stress. Exp. Hematol. 2016, 44, 92–100. [Google Scholar]
- Sepúlveda, V.E.; Márquez, R.; Turissini, D.A.; Goldman, W.E.; Matute, D.R. Genome Sequences Reveal Cryptic Speciation in the Human Pathogen Histoplasma capsulatum. mBio 2017, 8, e01339-17. [Google Scholar]
- Horwath, M.C.; Fecher, R.A.; Deepe, G.S., Jr. Histoplasma capsulatum, lung infection and immunity. Future Microbiol. 2015, 10, 967–975. [Google Scholar]
- Deepe, G.S. Histoplasma capsulatum. In Principles and Practice of Infectious Diseases; Mandell, G.L., Bennett, J.E., Dolin, R., Blaser, M.J., Eds.; Elsevier: Philadelphia, PA, USA, 2015; pp. 2949–2962. [Google Scholar]
- Yáñez, A.; Murciano, C.; O’Connor, J.E.; Gozalbo, D.; Gil, M.L. Candida albicans triggers proliferation and differentiation of hematopoietic stem and progenitor cells by a MyD88-dependent signaling. Microbes Infect. 2009, 11, 531–535. [Google Scholar]
- Megías, J.; Maneu, V.; Salvador, P.; Gozalbo, D.; Gil, M.L. Candida albicans stimulates in vivo differentiation of haematopoietic stem and progenitor cells towards macrophages by a TLR2-dependent signalling. Cell Microbiol. 2013, 15, 1143–1153. [Google Scholar]
- Rodriguez-Echeverri, C.; Puerta-Arias, J.D.; González, Á. Paracoccidioides brasiliensis activates mesenchymal stem cells through TLR2, TLR4, and Dectin-1. Med. Mycol. 2021, 59, 149–157. [Google Scholar]
- Gómez, L.F.; Arango, M.; McEwen, J.G.; Gómez, O.M.; Zuluaga, A.; Peláez, C.A.; Acevedo, J.M.; Taylor, M.L.; Jiménez, M.D.P. Molecular epidemiology of Colombian Histoplasma capsulatum isolates obtained from human and chicken manure samples. Heliyon 2019, 5, e02084. [Google Scholar]
- Costa, M.H.; de Soure, A.M.; Cabral, J.M.; Ferreira, F.C.; da Silva, C.L. Hematopoietic Niche—Exploring Biomimetic Cues to Improve the Functionality of Hematopoietic Stem/Progenitor Cells. Biotechnol. J. 2018, 13, 1700088. [Google Scholar] [CrossRef]
- Di Paolo, N.C.; Doronin, K.; Baldwin, L.K.; Papayannopoulou, T.; Shayakhmetov, D.M. The Transcription Factor IRF3 Triggers “Defensive Suicide” Necrosis in Response to Viral and Bacterial Pathogens. Cell Rep. 2013, 3, 1840–1846. [Google Scholar]
- Aravalli, R.N.; Hu, S.; Woods, J.P.; Lokensgard, J.R. Histoplasma capsulatum yeast phase-specific protein Yps3p induces Toll-like receptor 2 signaling. J. Neuroinflammation 2008, 5, 30. [Google Scholar]
- Höft, M.A.; Duvenage, L.; Hoving, J.C. Key thermally dimorphic fungal pathogens: Shaping host immunity. Open Biol. 2022, 12, 210219. [Google Scholar]
- Carvalho-Gontijo, R.; Moreira, D.R.; Resende, M.; Costa-Silva, M.F.; Peruhype-Magalhães, V.; Ribeiro, C.M.F.; Ribeiro, D.D.; Silvestre, R.; Cordeiro-Da-Silva, A.; Martins-Filho, O.A.; et al. Infection of hematopoietic stem cells by Leishmania infantum increases erythropoiesis and alters the phenotypic and functional profiles of progeny. Cell Immunol. 2018, 326, 77–85. [Google Scholar]
- Yáñez, A.; Flores, A.; Murciano, C.; O’Connor, J.E.; Gozalbo, D.; Gil, M.L. Signalling through TLR2/MyD88 induces differentiation of murine bone marrow stem and progenitor cells to functional phagocytes in response to Candida albicans. Cell Microbiol. 2010, 12, 114–128. [Google Scholar]
- Ray, S.C.; Rappleye, C.A. Flying under the radar: Histoplasma capsulatum avoidance of innate immune recognition. Semin. Cell Dev. Biol. 2019, 89, 91–98. [Google Scholar]
- Luther, K.; Torosantucci, A.; Brakhage, A.A.; Heesemann, J.; Ebel, F. Phagocytosis of Aspergillus fumigatus conidia by murine macrophages involves recognition by the dectin-1 beta-glucan receptor and Toll-like receptor 2. Cell Microbiol. 2007, 9, 368–381. [Google Scholar]
- Figueiredo, R.T.; Fernandez, P.L.; Dutra, F.F.; González, Y.; Lopes, L.C.; Bittencourt, V.C.B.; Sassaki, G.L.; Barreto-Bergter, E.; Bozza, M.T. TLR4 recognizes Pseudallescheria boydii conidia and purified rhamnomannans. J. Biol. Chem. 2010, 285, 40714–40723. [Google Scholar]
- Meier, A.; Kirschning, C.J.; Nikolaus, T.; Wagner, H.; Heesemann, J.; Ebel, F. Toll-like receptor (TLR) 2 and TLR4 are essential for Aspergillus-induced activation of murine macrophages. Cell Microbiol. 2003, 5, 561–570. [Google Scholar]
- Huang, J.-H.; Lin, C.-Y.; Wu, S.-Y.; Chen, W.-Y.; Chu, C.-L.; Brown, G.D.; Chuu, C.-P.; Wu-Hsieh, B.A.-Y. CR3 and Dectin-1 Collaborate in Macrophage Cytokine Response through Association on Lipid Rafts and Activation of Syk-JNK-AP-1 Pathway. PLoS Pathog. 2015, 11, e1004985. [Google Scholar]
- Smith, J.G.; Magee, D.M.; Williams, D.M.; Graybill, J.R. Tumor necrosis factor-alpha plays a role in host defense against Histoplasma capsulatum. J. Infect. Dis. 1990, 162, 1349–1353. [Google Scholar]
- Deepe, G.S., Jr.; Gibbons, R.S. Protective and memory immunity to Histoplasma capsulatum in the absence of IL-10. J. Immunol. 2003, 171, 5353–5362. [Google Scholar]
- Mittal, J.; Ponce, M.G.; Gendlina, I.; Nosanchuk, J.D. Histoplasma Capsulatum: Mechanisms for Pathogenesis. Curr. Top. Microbiol. Immunol. 2019, 422, 157–191. [Google Scholar]
- Hung, C.Y.; Xue, J.; Cole, G.T. Virulence mechanisms of Coccidioides. Ann. N. Y. Acad. Sci. 2007, 1111, 225–235. [Google Scholar]
- Diep, A.L.; Hoyer, K.K. Host Response to Coccidioides Infection: Fungal Immunity. Front. Cell. Infect. Microbiol. 2020, 10, 581101. [Google Scholar]
- Martínez, A.; Bono, C.; Gozalbo, D.; Goodridge, H.S.; Gil, M.L.; Yáñez, A. TLR2 and Dectin-1 Signaling in Mouse Hematopoietic Stem and Progenitor Cells Impacts the Ability of the Antigen Presenting Cells They Produce to Activate CD4 T Cells. Cells 2020, 9, 1317. [Google Scholar]
- Bono, C.; Martínez, A.; Megías, J.; Gozalbo, D.; Yáñez, A.; Gil, M.L. Dectin-1 Stimulation of Hematopoietic Stem and Progenitor Cells Occurs in Vivo and Promotes Differentiation toward Trained Macrophages via an Indirect Cell-Autonomous Mechanism. mBio 2020, 11, e00781-20. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| GAPDH | CATGGCCTTCCGTGTTCCTA | GCGGCACGTCAGATCCA |
| IL-6 | CAACCACGGCCTTCCCTACTTC | TCTCATTTCCACGATTTCCCAGAG |
| IL-17 | CCAAACACTGAGGCCAAGGACTTC | GGTGACGTGGAACGGTTGAGGTAG |
| IL-1β | CTTCAAATCTCGCAGCAGCACATC | TCCACGGGAAAGACACAGGTAGC |
| IL-10 | TGGGTTGCCAAGCCTTATCGG | CTCACCCAGGGAATTCAAATGCTC |
| TNF-α | GACAAGGCTGCCCCGACTACG | CTTGGGGCAGGGGCTCTTGAC |
| TGFβ | TACTGCCGCTTCTGCTCCCACTCC | TCGATGCGCTTCCGTTTCACCAG |
| Arg-1 | CCTTGGCTTGCTTCGGAACTCA | CTTGGGAGGAGAAGGCGTTTGC |
| iNOS | GCCGCATGAGCTTGGTGTTTG | GCAGCCGGGAGTAGCCTGTGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Echeverri, C.; Gómez, B.L.; González, Á. Histoplasma capsulatum Activates Hematopoietic Stem Cells and Their Progenitors through a Mechanism Dependent on TLR2, TLR4, and Dectin-1. J. Fungi 2022, 8, 1108. https://doi.org/10.3390/jof8101108
Rodríguez-Echeverri C, Gómez BL, González Á. Histoplasma capsulatum Activates Hematopoietic Stem Cells and Their Progenitors through a Mechanism Dependent on TLR2, TLR4, and Dectin-1. Journal of Fungi. 2022; 8(10):1108. https://doi.org/10.3390/jof8101108
Chicago/Turabian StyleRodríguez-Echeverri, Carolina, Beatriz L. Gómez, and Ángel González. 2022. "Histoplasma capsulatum Activates Hematopoietic Stem Cells and Their Progenitors through a Mechanism Dependent on TLR2, TLR4, and Dectin-1" Journal of Fungi 8, no. 10: 1108. https://doi.org/10.3390/jof8101108