
The Effects of Sex and Strain on Pneumocystis murina Fungal Burdens in Mice
 , , and
, , and {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
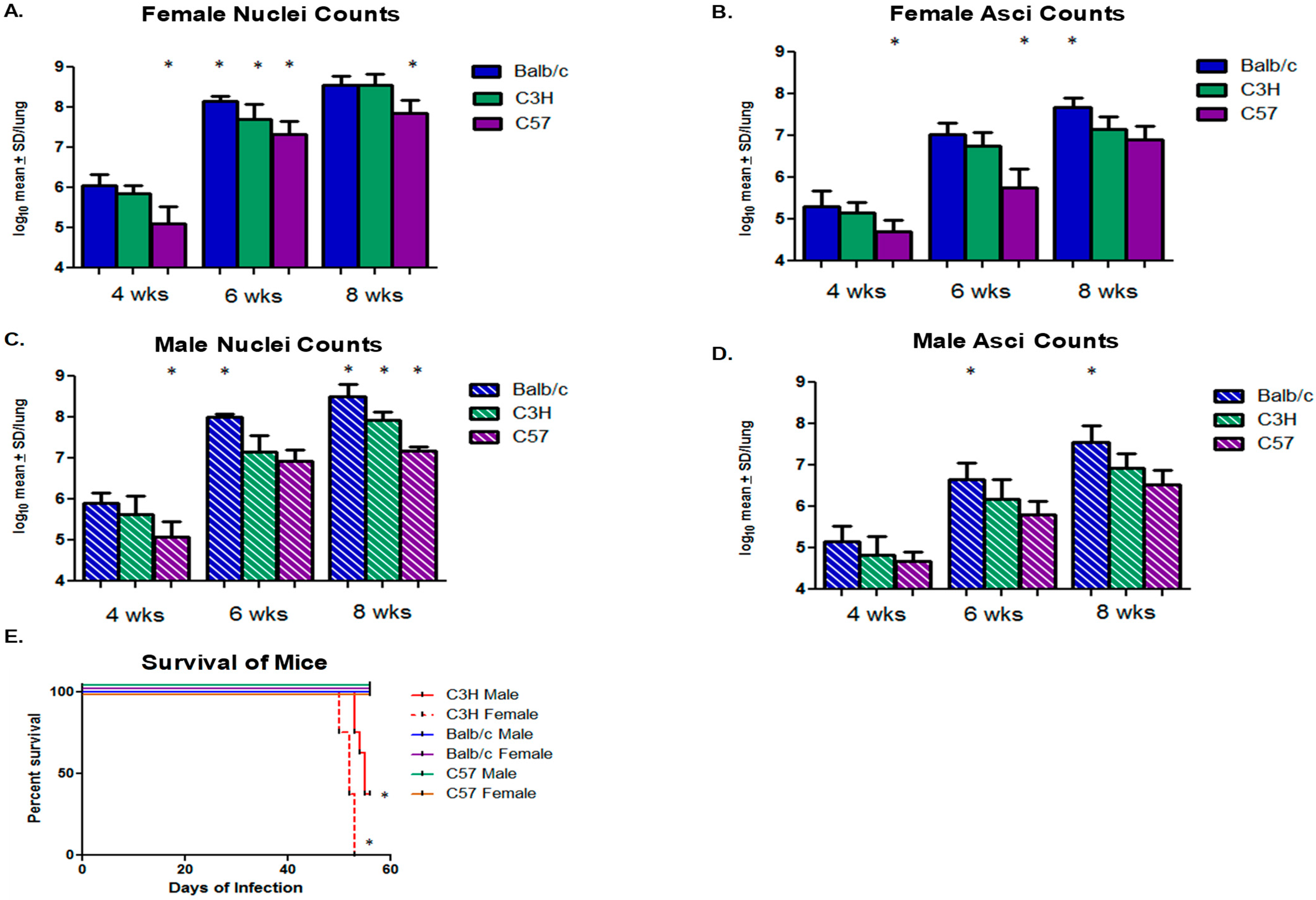
2.1. Design
2.2. Evaluation of Organism Burden
2.3. Microscopic Enumeration
2.4. Survival Curve
3. Results and Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Soldin, O.P.; Mattison, D.R. Sex differences in pharmacokinetics and pharmacodynamics. Clin. Pharmacokinet. 2009, 48, 143–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beery, A.K.; Zucker, I. Sex bias in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2011, 35, 565–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clayton, J.A.; Collins, F.S. Policy: NIH to balance sex in cell and animal studies. Nature 2014, 509, 282–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaillon, S.; Berthenet, K.; Garlanda, C. Sexual Dimorphism in Innate Immunity. Clin. Rev. Allergy Immunol. 2019, 56, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.R.; Kunz, C.; Dietze-Schwonberg, K.; Lorenz, B.; von Stebut, E. Myeloid cells do not contribute to gender-dependent differences in disease outcome in murine cutaneous leishmaniasis. Cell. Immunol. 2016, 308, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Dkhil, M.A.; Al-Shaebi, E.M.; Lubbad, M.Y.; Al-Quraishy, S. Impact of sex differences in brain response to infection with Plasmodium berghei. Parasitol. Res. 2016, 115, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Bastian, L.; Bennett, C.L.; Adams, J.; Waskin, H.; Divine, G.; Edlin, B.R. Differences between men and women with HIV-related Pneumocystis carinii pneumonia: Experience from 3070 cases in New York City in 1987. J. Acquir. Immune Defic. Syndr. 1993, 6, 617–623. [Google Scholar] [PubMed]
- Liu, Y.; Su, L.; Jiang, S.J.; Qu, H. Risk factors for mortality from Pneumocystis carinii pneumonia (PCP) in non-HIV patients: A meta-analysis. Oncotarget 2017, 8, 59729–59739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walzer, P.D.; Foy, J.; Steele, P.; White, M. Treatment of experimental pneumocystosis: Review of 7 years of experience and development of a new system for classifying antimicrobial drugs. Antimicrob. Agents Chemother. 1992, 36, 1943–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFadden, D.C.; Powles, M.A.; Pittarelli, L.A.; Schmatz, D.M. Establishment of Pneumocystis carinii in various mouse strains using natural transmission to initiate infection. J. Protozool. 1991, 38, 126S–127S. [Google Scholar] [PubMed]
- Cushion, M.T.; Walzer, P.D.; Ashbaugh, A.; Rebholz, S.; Brubaker, R.; Eynde, J.J.V.; Mayence, A.; Huang, T.L. In vitro selection and in vivo efficacy of piperazine- and alkanediamide-linked bisbenzamidines against Pneumocystis pneumonia in mice. Antimicrob. Agents Chemother. 2006, 50, 2337–2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cushion, M.T.; Linke, M.J.; Ashbaugh, A.; Sesterhenn, T.; Collins, M.S.; Lynch, K.; Brubaker, R.; Walzer, P.D. Echinocandin treatment of Pneumocystis pneumonia in rodent models depletes cysts leaving trophic burdens that cannot transmit the infection. PLoS ONE 2010, 5, e8524. [Google Scholar] [CrossRef] [PubMed]
- Cushion, M.T.; Ruffolo, J.J.; Walzer, P.D. Analysis of the developmental stages of Pneumocystis carinii, in vitro. Lab. Investig. 1988, 58, 324–331. [Google Scholar] [PubMed]
- Cushion, M.T.; Linke, M.J.; Collins, M.; Keely, S.P.; Stringer, J.R. The minimum number of Pneumocystis carinii f. sp. carinii organisms required to establish infections is very low. J. Eukaryot. Microbiol. 1999, 46, 111S. [Google Scholar] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macioce-Tisdale, N.L.; Ashbaugh, A.; Hendrix, K.; Collins, M.S.; Porollo, A.; Cushion, M.T. The Effects of Sex and Strain on Pneumocystis murina Fungal Burdens in Mice. J. Fungi 2022, 8, 1101. https://doi.org/10.3390/jof8101101
Macioce-Tisdale NL, Ashbaugh A, Hendrix K, Collins MS, Porollo A, Cushion MT. The Effects of Sex and Strain on Pneumocystis murina Fungal Burdens in Mice. Journal of Fungi. 2022; 8(10):1101. https://doi.org/10.3390/jof8101101
Chicago/Turabian StyleMacioce-Tisdale, Nikeya L., Alan Ashbaugh, Keeley Hendrix, Margaret S. Collins, Aleksey Porollo, and Melanie T. Cushion. 2022. "The Effects of Sex and Strain on Pneumocystis murina Fungal Burdens in Mice" Journal of Fungi 8, no. 10: 1101. https://doi.org/10.3390/jof8101101