
Regulator of G Protein Signaling Proteins Control Growth, Development and Cellulase Production in Neurospora crassa

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media, Strains and Genetic Procedures
2.2. Growth and Developmental Phenotypic Assays
2.3. Assessment of Gene Expression, Growth, Supernatant Protein, Cell Mass Protein and Cellulase Activity Measurements with Avicel as the Carbon Source
2.4. Statistical Analysis
3. Results
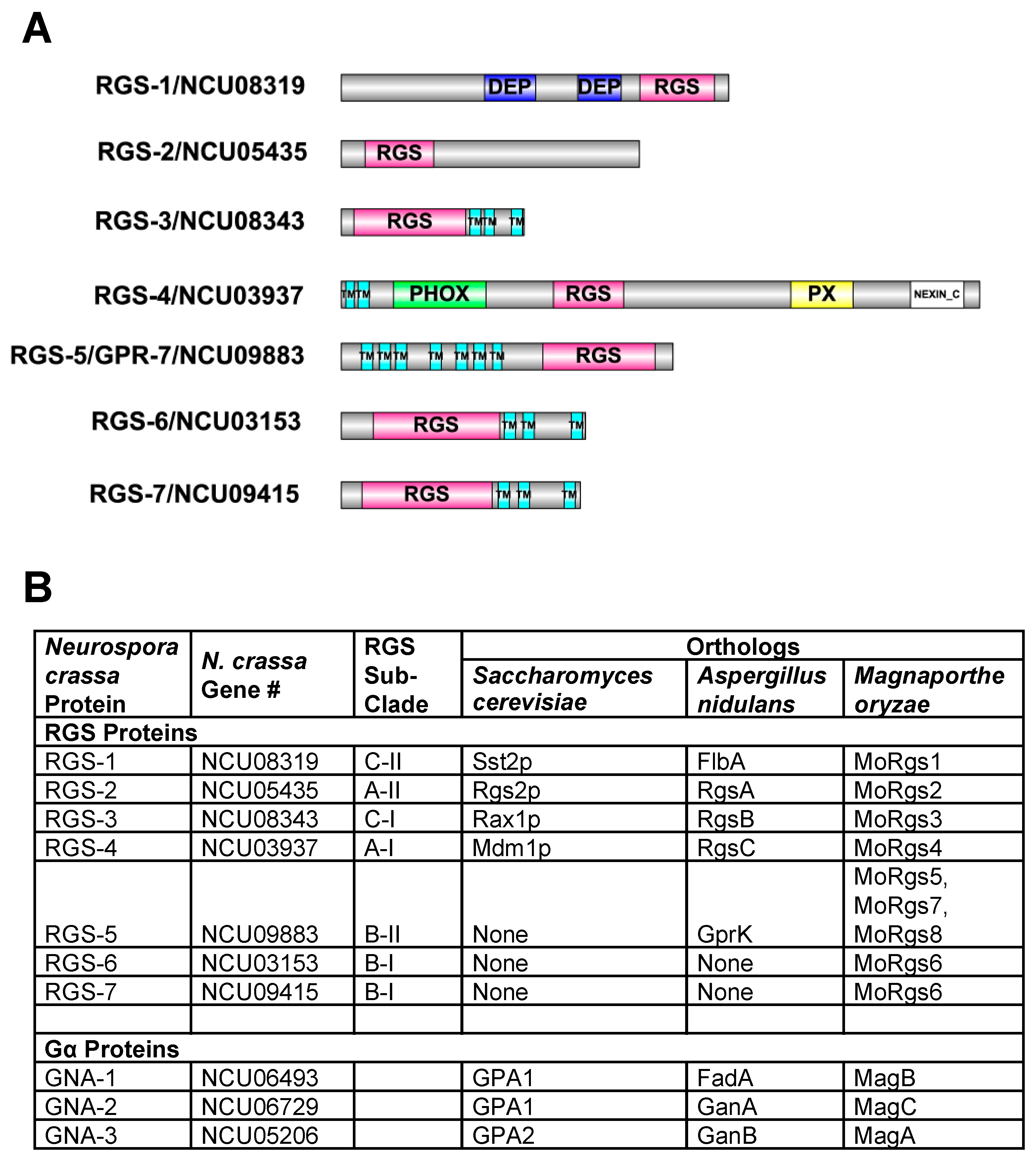
3.1. The N. crassa Genome Contains Seven Predicted RGS Genes
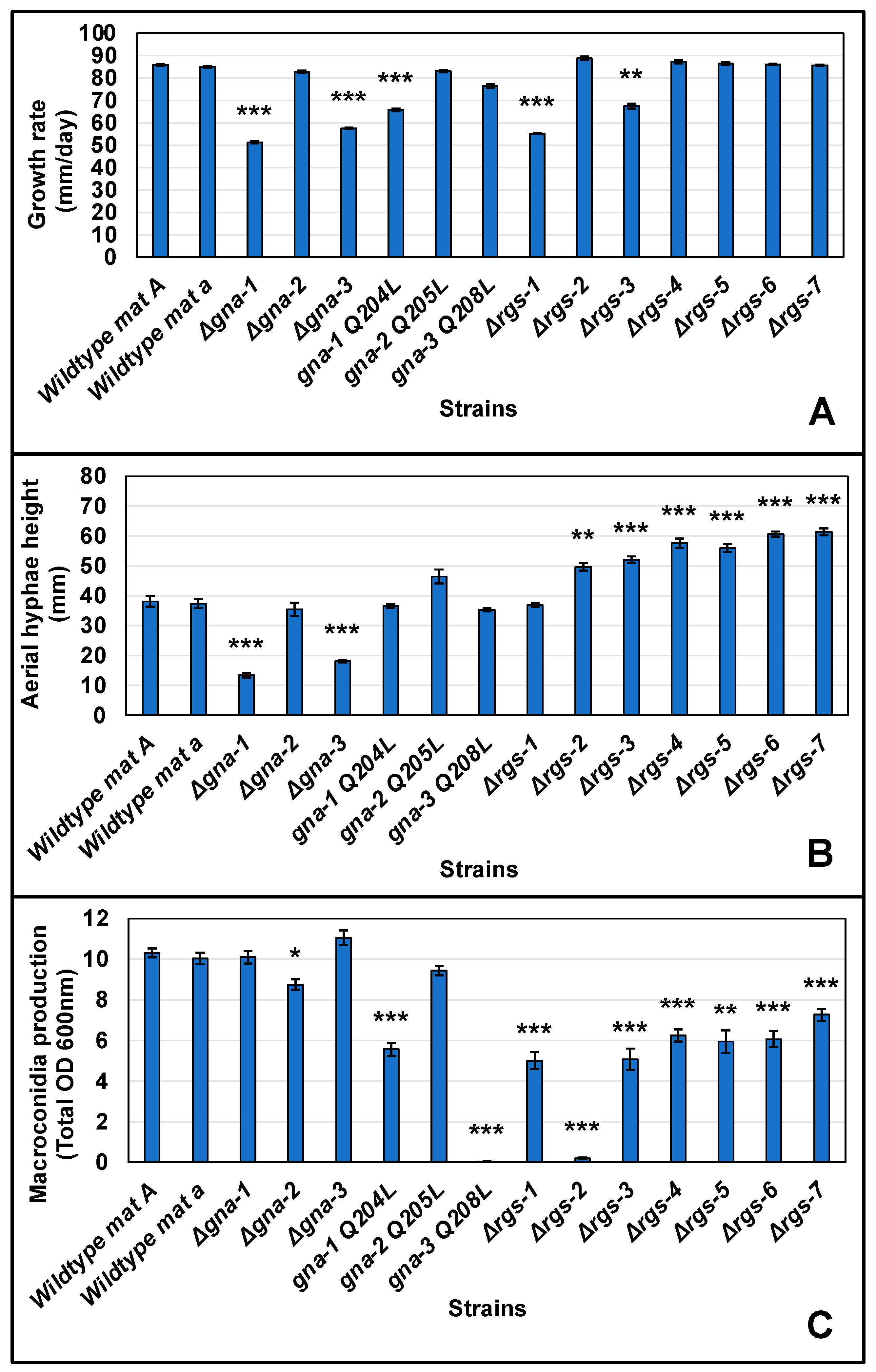
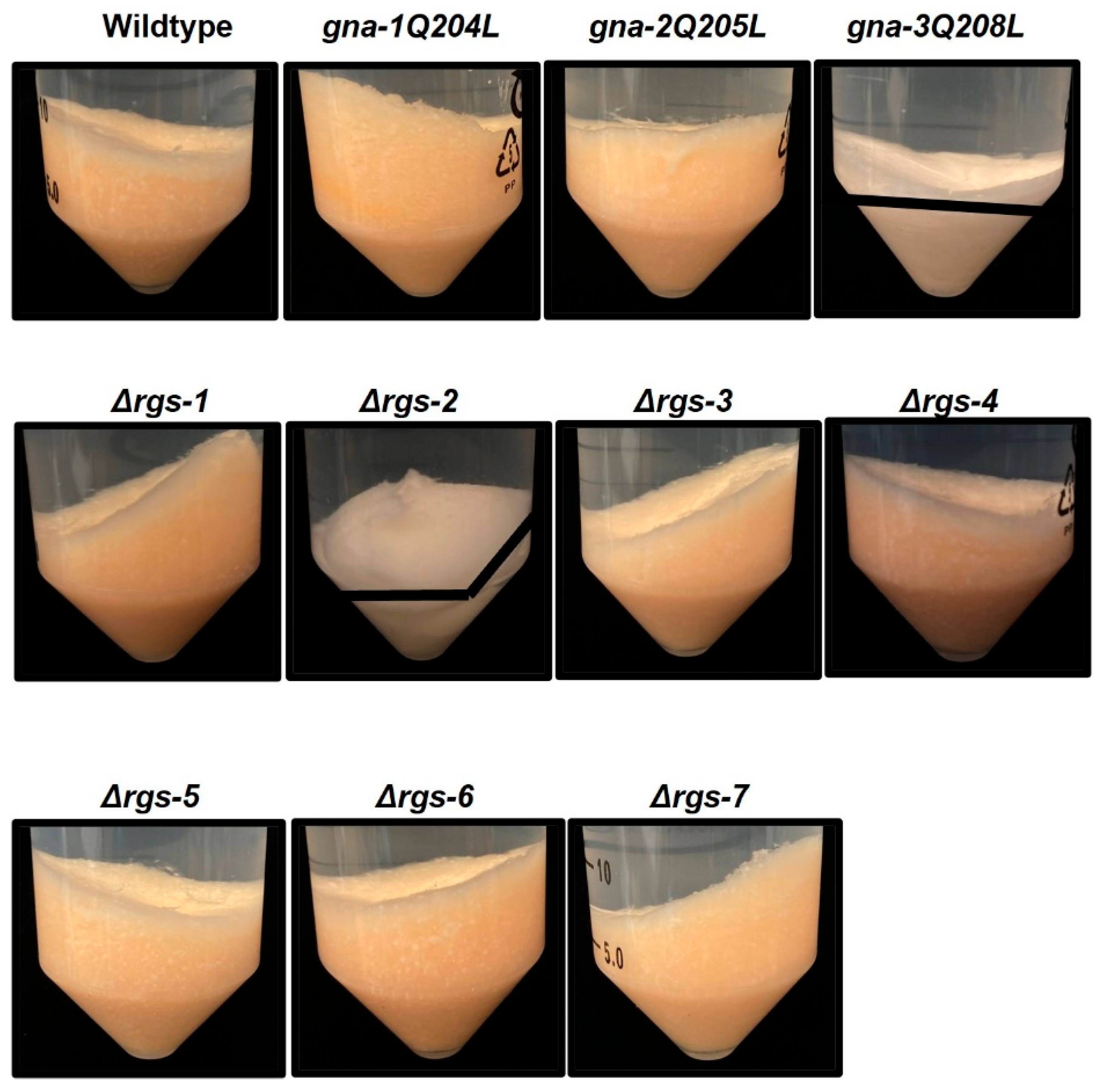
3.2. Assessment of Quantitative Phenotypes in Strains with Mutations in RGS or Gα Subunit Genes during Growth and Asexual Development
3.3. rgs-1 and rgs-2 Exhibit Epistatic Interactions with gna-1 or gna-3 during the Sexual Cycle
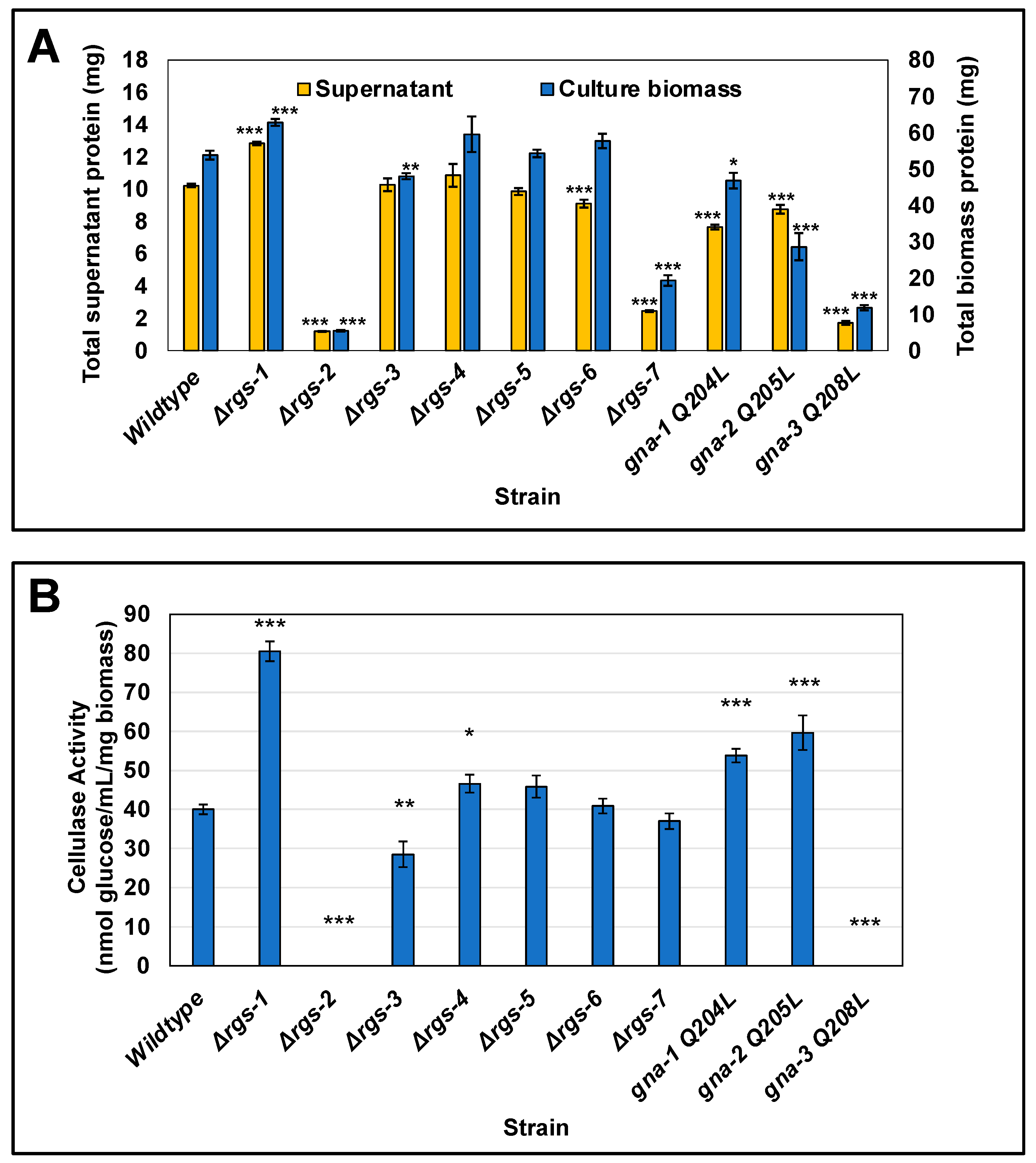
3.4. Five of the RGS Gene Deletion Mutants Have Phenotypes during Growth on Cellulose and/or in Extracellular Cellulase Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borkovich, K.A.; Alex, L.A.; Yarden, O.; Freitag, M.; Turner, G.E.; Read, N.D.; Seiler, S.; Bell-Pedersen, D.; Paietta, J.; Plesofsky, N.; et al. Lessons from the genome sequence of Neurospora crassa: Tracing the path from genomic blueprint to multicellular organism. Microbiol. Mol. Biol. Rev. 2004, 68, 1–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selker, E.U. Neurospora. Curr. Biol. 2011, 21, R139–R140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, R.H.; Perkins, D.D. Timeline: Neurospora: A model of model microbes. Nat. Rev. Genet. 2002, 3, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.D. Septum formation in Aspergillus nidulans. Curr. Opin. Microbiol. 2001, 4, 736–739. [Google Scholar] [CrossRef]
- Fleissner, A.; Serrano, A. The Art of Networking: Vegetative Hyphal Fusion in Filamentous Ascomycete Fungi. In The Mycota: Growth, Differentiation and Sexuality; Wendland, J., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1, pp. 133–153. [Google Scholar]
- Springer, M.L. Genetic control of fungal differentiation: The three sporulation pathways of Neurospora crassa. Bioessays 1993, 15, 365–374. [Google Scholar] [CrossRef]
- Raju, N.B. Neurospora as a model fungus for studies in cytogenetics and sexual biology at Stanford. J. Biosci. 2009, 34, 139–159. [Google Scholar] [CrossRef]
- Barratt, R.W.; Garnjobst, L. Genetics of a Colonial Microconidiating Mutant Strain of Neurospora crassa. Genetics 1949, 34, 351–369. [Google Scholar] [CrossRef]
- Li, L.; Wright, S.J.; Krystofova, S.; Park, G.; Borkovich, K.A. Heterotrimeric G protein signaling in filamentous fungi. Annu. Rev. Microbiol. 2007, 61, 423–452. [Google Scholar] [CrossRef]
- Turner, G.E.; Borkovich, K.A. Identification of a G protein alpha subunit from Neurospora crassa that is a member of the Gi family. J. Biol. Chem. 1993, 268, 14805–14811. [Google Scholar] [CrossRef]
- Ivey, F.D.; Hodge, P.N.; Turner, G.E.; Borkovich, K.A. The G alpha i homologue gna-1 controls multiple differentiation pathways in Neurospora crassa. Mol. Biol. Cell 1996, 7, 1283–1297. [Google Scholar] [CrossRef]
- Baasiri, R.A.; Lu, X.; Rowley, P.S.; Turner, G.E.; Borkovich, K.A. Overlapping functions for two G protein alpha subunits in Neurospora crassa. Genetics 1997, 147, 137–145. [Google Scholar] [CrossRef]
- Kays, A.M.; Rowley, P.S.; Baasiri, R.A.; Borkovich, K.A. Regulation of conidiation and adenylyl cyclase levels by the Galpha protein GNA-3 in Neurospora crassa. Mol. Cell Biol. 2000, 20, 7693–7705. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Poole, S.I.; Borkovich, K.A. A G-protein beta subunit required for sexual and vegetative development and maintenance of normal G alpha protein levels in Neurospora crassa. Eukaryot. Cell 2002, 1, 378–390. [Google Scholar] [CrossRef] [Green Version]
- Muller, F.; Kruger, D.; Sattlegger, E.; Hoffmann, B.; Ballario, P.; Kanaan, M.; Barthelmess, I.B. The cpc-2 gene of Neurospora crassa encodes a protein entirely composed of WD-repeat segments that is involved in general amino acid control and female fertility. Mol. Gen. Genet. MGG 1995, 248, 162–173. [Google Scholar] [CrossRef]
- Garud, A.; Carrillo, A.J.; Collier, L.A.; Ghosh, A.; Kim, J.D.; Lopez-Lopez, B.; Ouyang, S.; Borkovich, K.A. Genetic relationships between the RACK1 homolog cpc-2 and heterotrimeric G protein subunit genes in Neurospora crassa. PLoS ONE 2019, 14, e0223334. [Google Scholar] [CrossRef] [Green Version]
- Krystofova, S.; Borkovich, K.A. The heterotrimeric G-protein subunits GNG-1 and GNB-1 form a Gbetagamma dimer required for normal female fertility, asexual development, and galpha protein levels in Neurospora crassa. Eukaryot. Cell 2005, 4, 365–378. [Google Scholar] [CrossRef] [Green Version]
- Ivey, F.D.; Yang, Q.; Borkovich, K.A. Positive regulation of adenylyl cyclase activity by a galphai homolog in Neurospora crassa. Fungal Genet. Biol. 1999, 26, 48–61. [Google Scholar] [CrossRef]
- Yang, Q.; Borkovich, K.A. Mutational activation of a Galphai causes uncontrolled proliferation of aerial hyphae and increased sensitivity to heat and oxidative stress in Neurospora crassa. Genetics 1999, 151, 107–117. [Google Scholar] [CrossRef]
- Kays, A.M.; Borkovich, K.A. Severe impairment of growth and differentiation in a Neurospora crassa mutant lacking all heterotrimeric G alpha proteins. Genetics 2004, 166, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- Collier, L.A.; Ghosh, A.; Borkovich, K.A. Heterotrimeric G-Protein Signaling Is Required for Cellulose Degradation in Neurospora crassa. Mbio 2020, 11, e02419-20. [Google Scholar] [CrossRef]
- Alqinyah, M.; Hooks, S.B. Regulating the regulators: Epigenetic, transcriptional, and post-translational regulation of RGS proteins. Cell Signal. 2018, 42, 77–87. [Google Scholar] [CrossRef]
- Ross, E.M.; Wilkie, T.M. GTPase-activating proteins for heterotrimeric G proteins: Regulators of G protein signaling (RGS) and RGS-like proteins. Annu. Rev. Biochem. 2000, 69, 795–827. [Google Scholar] [CrossRef] [Green Version]
- Koelle, M.R. A new family of G-protein regulators—The RGS proteins. Curr. Opin. Cell Biol. 1997, 9, 143–147. [Google Scholar] [CrossRef]
- Watson, N.; Linder, M.E.; Druey, K.M.; Kehrl, J.H.; Blumer, K.J. RGS family members: GTPase-activating proteins for heterotrimeric G-protein alpha-subunits. Nature 1996, 383, 172–175. [Google Scholar] [CrossRef]
- Wang, Y.; Geng, Z.; Jiang, D.; Long, F.; Zhao, Y.; Su, H.; Zhang, K.Q.; Yang, J. Characterizations and functions of regulator of G protein signaling (RGS) in fungi. Appl. Microbiol. Biotechnol. 2013, 97, 7977–7987. [Google Scholar] [CrossRef]
- Jung, M.G.; Kim, S.S.; Yu, J.H.; Shin, K.S. Characterization of gprK Encoding a Putative Hybrid G-Protein-Coupled Receptor in Aspergillus fumigatus. PLoS ONE 2016, 11, e0161312. [Google Scholar] [CrossRef] [Green Version]
- Igbalajobi, O.A.; Yu, J.H.; Shin, K.S. Characterization of the rax1 gene encoding a putative regulator of G protein signaling in Aspergillus fumigatus. Biochem. Biophys. Res. Commun. 2017, 487, 426–432. [Google Scholar] [CrossRef]
- Kim, Y.; Heo, I.B.; Yu, J.H.; Shin, K.S. Characteristics of a Regulator of G-Protein Signaling (RGS) rgsC in Aspergillus fumigatus. Front. Microbiol. 2017, 8, 2058. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, M.W.; Jun, S.C.; Choi, Y.H.; Yu, J.H.; Shin, K.S. RgsD negatively controls development, toxigenesis, stress response, and virulence in Aspergillus fumigatus. Sci. Rep. 2019, 9, 811. [Google Scholar] [CrossRef] [Green Version]
- Xie, R.; Yang, K.; Tumukunde, E.; Guo, Z.; Zhang, B.; Liu, Y.; Zhuang, Z.; Yuan, J.; Wang, S. Regulator of G Protein Signaling Contributes to the Development and Aflatoxin Biosynthesis in Aspergillus flavus through the Regulation of Galpha Activity. Appl. Environ. Microbiol. 2022, 88, e0024422. [Google Scholar] [CrossRef]
- Bosnjak, N.; Smith, K.M.; Asaria, I.; Lahola-Chomiak, A.; Kishore, N.; Todd, A.T.; Freitag, M.; Nargang, F.E. Involvement of a G Protein Regulatory Circuit in Alternative Oxidase Production in Neurospora crassa. G3 Genes Genomes Genet. 2019, 9, 3453–3465. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.J. Distribution of lysine pathways among fungi: Evolutionary implications. Am. Nat. 1964, 98, 435–446. [Google Scholar] [CrossRef]
- Davis, R.H.; de Serres, F.J. Genetic and microbiological research techniques for Neurospora crassa. Methods Enzymol. 1970, 17, 79–143. [Google Scholar]
- Kuck, U.; Hoff, B. Application of the nourseothricin acetyltransferase gene (nat1) as dominant marker for the transformation of filamentous fungi. Fungal Genet. Newsl. 2006, 53, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Hays, S.; Selker, E.U. Making the selectable marker bar tighter and more economical. Fungal Genet. Rep. 2000, 47, 25. [Google Scholar] [CrossRef] [Green Version]
- Metzenberg, R.L.; Jacobson, D.J.; Bertrand, H. Making the selective agent for the bar plasmids, phoshoinothricin (glufosinate) affordable for routine use. Fungal Genet. Rep. 2000, 47, 13. [Google Scholar] [CrossRef] [Green Version]
- Colot, H.V.; Park, G.; Turner, G.E.; Ringelberg, C.; Crew, C.M.; Litvinkova, L.; Weiss, R.L.; Borkovich, K.A.; Dunlap, J.C. A high-throughput gene knockout procedure for Neurospora reveals functions for multiple transcription factors. Proc. Natl. Acad. Sci. USA 2006, 103, 10352–10357. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, S.; Beecher, C.N.; Wang, K.; Larive, C.K.; Borkovich, K.A. Metabolic Impacts of Using Nitrogen and Copper-Regulated Promoters to Regulate Gene Expression in Neurospora crassa. G3 Genes Genomes Genet. 2015, 5, 1899–1908. [Google Scholar] [CrossRef] [Green Version]
- Freitag, M.; Hickey, P.C.; Raju, N.B.; Selker, E.U.; Read, N.D. GFP as a tool to analyze the organization, dynamics and function of nuclei and microtubules in Neurospora crassa. Fungal Genet. Biol. 2004, 41, 897–910. [Google Scholar] [CrossRef]
- Westergaard, M.; Mitchell, H.K.; Neurospora, V. A synthetic medium favoring sexual reproduction. Am. J. Bot. 1947, 34, 573–577. [Google Scholar] [CrossRef]
- Coradetti, S.T.; Craig, J.P.; Xiong, Y.; Shock, T.; Tian, C.; Glass, N.L. Conserved and essential transcription factors for cellulase gene expression in ascomycete fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 7397–7402. [Google Scholar] [CrossRef]
- Grubbs, F. Procedures for detecting outlying observations in samples. Tecnometrics 1969, 11, 1–21. [Google Scholar] [CrossRef]
- Student. The probable error of a mean. Biometrika 1908, 6, 1–25. [Google Scholar] [CrossRef]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. (Use R); Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar] [CrossRef]
- Soundararajan, M.; Willard, F.S.; Kimple, A.J.; Turnbull, A.P.; Ball, L.J.; Schoch, G.A.; Gileadi, C.; Fedorov, O.Y.; Dowler, E.F.; Higman, V.A.; et al. Structural diversity in the RGS domain and its interaction with heterotrimeric G protein alpha-subunits. Proc. Natl. Acad. Sci. USA 2008, 105, 6457–6462. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, I.E.; Pacentine, I.V.; Lim, A.; Guerrero, N.; Krystofova, S.; Li, L.; Michkov, A.V.; Servin, J.A.; Ahrendt, S.R.; Carrillo, A.J.; et al. Global Analysis of Predicted G Protein-Coupled Receptor Genes in the Filamentous Fungus, Neurospora crassa. G3 Genes Genomes Genet. 2015, 5, 2729–2743. [Google Scholar] [CrossRef] [Green Version]
- Consonni, S.V.; Maurice, M.M.; Bos, J.L. DEP domains: Structurally similar but functionally different. Nat. Rev. Mol. Cell Biol 2014, 15, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.W.; Lemmon, M.A. All phox homology (PX) domains from Saccharomyces cerevisiae specifically recognize phosphatidylinositol 3-phosphate. J. Biol. Chem. 2001, 276, 44179–44184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosomi, A.; Kawanishi, Y.Y.; Tanaka, N.; Takegawa, K. PXA domain-containing protein Pxa1 is required for normal vacuole function and morphology in Schizosaccharomyces pombe. Biosci. Biotechnol. Biochem. 2008, 72, 548–556. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, R.D.; Loci, D.; Houghton, F.; Karlsson, L.; Gleeson, P.A. A large family of endosome-localized proteins related to sorting nexin 1. Biochem. J. 2001, 358, 7–16. [Google Scholar] [CrossRef]
- Wright, S.J.; Inchausti, R.; Eaton, C.J.; Krystofova, S.; Borkovich, K.A. RIC8 is a guanine-nucleotide exchange factor for Galpha subunits that regulates growth and development in Neurospora crassa. Genetics 2011, 189, 165–176. [Google Scholar] [CrossRef]
- Ivey, F.D.; Kays, A.M.; Borkovich, K.A. Shared and independent roles for a Galpha(i) protein and adenylyl cyclase in regulating development and stress responses in Neurospora crassa. Eukaryot. Cell 2002, 1, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Glass, N.L.; Schmoll, M.; Cate, J.H.; Coradetti, S. Plant cell wall deconstruction by ascomycete fungi. Annu. Rev. Microbiol. 2013, 67, 477–498. [Google Scholar] [CrossRef]
- Tian, C.; Beeson, W.T.; Iavarone, A.T.; Sun, J.; Marletta, M.A.; Cate, J.H.; Glass, N.L. Systems analysis of plant cell wall degradation by the model filamentous fungus Neurospora crassa. Proc. Natl. Acad. Sci. USA 2009, 106, 22157–22162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.H.; Wieser, J.; Adams, T.H. The Aspergillus FlbA RGS domain protein antagonizes G protein signaling to block proliferation and allow development. EMBO J. 1996, 15, 5184–5190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tang, W.; Liu, K.; Huang, Q.; Zhang, X.; Yan, X.; Chen, Y.; Wang, J.; Qi, Z.; Wang, Z.; et al. Eight RGS and RGS-like proteins orchestrate growth, differentiation, and pathogenicity of Magnaporthe oryzae. PLoS Pathog. 2011, 7, e1002450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafuente, M.J.; Gancedo, C. Disruption and basic functional analysis of six novel ORFs of chromosome XV from Saccharomyces cerevisiae. Yeast 1999, 15, 935–943. [Google Scholar] [CrossRef]
- Han, K.H.; Seo, J.A.; Yu, J.H. Regulators of G-protein signalling in Aspergillus nidulans: RgsA downregulates stress response and stimulates asexual sporulation through attenuation of GanB (Galpha) signalling. Mol. Microbiol. 2004, 53, 529–540. [Google Scholar] [CrossRef]
- Rosen, S.; Yu, J.H.; Adams, T.H. The Aspergillus nidulans sfaD gene encodes a G protein beta subunit that is required for normal growth and repression of sporulation. EMBO J. 1999, 18, 5592–5600. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Borkovich, K.A. A pheromone receptor gene, pre-1, is essential for mating type-specific directional growth and fusion of trichogynes and female fertility in Neurospora crassa. Mol. Microbiol. 2004, 52, 1781–1798. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relevant Genotype | Strain Name | Detailed Genotype | NCU Number | Source |
|---|---|---|---|---|
| Wild type | 74-OR23-1A | Wild type, mat A | NA 2 | FGSC2489 1 |
| Wild type | OR8-1a | Wild type mat a | NA | FGSC4200 |
| ∆gna-1 | 3b10 | ∆gna-1:hph, mat a | NCU06493 | Ivey et al., 1999 [18] |
| ∆gna-2 | gna-2 a | ∆gna-2:hph, mat a | NCU06729 | FGSC12377 |
| ∆gna-3 | 31c2 | ∆gna-3:hph, mat A | NCU05206 | Kays et al., 2000 [13] |
| gna-1* | ∆1gna-1* | ∆gna-1:hph, gna-1Q204L:his-3+, mat A | NA | Collier et al., 2020 [21] |
| gna-2* | G2-7 | pccg-1::gna-2Q205L::his-3+, mat A | NA | Collier et al., 2020 [21] |
| gna-3* | gna3Q208L | pccg-1::gna-3Q208L::his-3+, mat A | NA | Collier et al., 2020 [21] |
| ∆rgs-1 | rgs-1-2a | ∆rgs-1::hph, mat a | NCU08319 | This Study |
| ∆rgs-2 | rgs-2-7a | ∆rgs-2::hph, mat a | NCU05435 | This Study |
| ∆rgs-3 | rgs-3-2a | ∆rgs-3::hph, mat a | NCU08343 | This Study |
| ∆rgs-4 | rgs-4-7A | ∆rgs-4::hph, mat A | NCU03937 | This Study |
| ∆rgs-5 | rgs-5-3 | ∆rgs-5::hph, mat a | NCU09883 | This Study |
| ∆rgs-6 | rgs-6-a | ∆rgs-6::hph, mat a | NCU03153 | FGSC14041 |
| ∆rgs-7 | rgs-7-A | ∆rgs-7::hph, mat A | NCU09415 | FGSC15515 |
| ∆rgs-1, ∆mus-52 | rgs1m7 | ∆rgs-1::hph, ∆mus-52::nat, mat a | NA | This Study |
| ∆rgs-2, ∆mus-52 | rgs2m2 | ∆rgs-2::hph, ∆mus-52::nat, mat a | NA | This Study |
| ∆rgs-3, ∆mus-52 | rgs3m1 | ∆rgs-3::hph, ∆mus-52::nat, mat a | NA | This Study |
| ∆rgs-4, ∆mus-52 | rgs4m5 | ∆rgs-4::hph, ∆mus-52::nat, mat a | NA | This Study |
| ∆rgs-5, ∆mus-52 | rgs5m4 | ∆rgs-5::hph, ∆mus-52::nat, mat A | NA | This Study |
| ∆rgs-6, ∆mus-52 | rgs6m1 | ∆rgs-6::hph, ∆mus-52::nat, mat a | NA | This Study |
| ∆rgs-6, ∆mus-52 | rgs6m5 | ∆rgs-6::hph, ∆mus-52::nat, mat a | NA | This Study |
| ∆rgs-7, ∆mus-52 | rgs7m1 | ∆rgs-7::hph, ∆mus-52::nat, mat a | NA | This Study |
| ∆rgs-1, rgs-1+ | rgs1m7-c2 | ∆rgs-1::hph, ∆mus-52::nat, pccg-1::rgs-1+::pan-2, mat a | NA | This Study |
| ∆rgs-2, rgs-2+ | rgs2m2-c2 | ∆rgs-2::hph, ∆mus-52::nat, pccg-1::rgs-2+::pan-2, mat a | NA | This Study |
| ∆rgs-3, rgs-3+ | rgs3m1-c5 | ∆rgs-3::hph, ∆mus-52::nat, pccg-1::rgs-3+::pan-2, mat a | NA | This Study |
| ∆rgs-4, rgs-4+ | rgs4m4-c1 | ∆rgs-4::hph, ∆mus-52::nat, pccg-1::rgs-4+::pan-2, mat a | NA | This Study |
| ∆rgs-5, rgs-5+ | rgs5m4-c8 | ∆rgs-5::hph, ∆mus-52::nat, pccg-1::rgs-5+::pan-2, mat A | NA | This Study |
| ∆rgs-6, rgs-6+ | rgs6m1-c3 | ∆rgs-6::hph, ∆mus-52::nat, pccg-1::rgs-6+::pan-2, mat a | NA | This Study |
| ∆rgs-7, rgs-7+ | rgs7m1-c1 | ∆rgs-7::hph, ∆mus-52::nat, pccg-1::rgs-7+::pan-2, mat a | NA | This Study |
| Strain Name | Relevant Genotype 1 | Protoperithecia | Perithecia | Ascospores |
|---|---|---|---|---|
| 74-OR23-1A | Wild type, mat A | Normal | Normal | Normal |
| OR8-1a | Wild type, mat a | Normal | Normal | Normal |
| 3b10 | ∆gna-1 | Normal | Abnormal | Not Formed |
| gna-2 a | ∆gna-2 | Normal | Normal | Normal |
| 31c2 | ∆gna-3 | Reduced | Reduced | Normal |
| ∆1gna-1* | gna-1Q204L | Reduced | Reduced | Reduced |
| G2-7 | gna-2Q205L | Normal | Normal | Normal |
| gna3Q208L | gna-3Q208L | Not Formed | Not Formed | Not Formed |
| rgs-1-2a | ∆rgs-1 | Not Formed | Not Formed | Not Formed |
| rgs-2-7a | ∆rgs-2 | Normal | Reduced | Reduced |
| rgs-3-2a | ∆rgs-3 | Normal | Normal | Normal |
| rgs-4-7A | ∆rgs-4 | Normal | Normal | Normal |
| rgs-5-3 | ∆rgs-5 | Normal | Normal | Normal |
| rgs-6-a | ∆rgs-6 | Normal | Normal | Normal |
| rgs-7-A | ∆rgs-7 | Normal | Normal | Normal |
| Function | RGS Protein Regulation Type/Possible Gα Partner(s) Strongly Supported Partners Are Underlined. —; No Effect due to Mutation of RGS | ||||||
|---|---|---|---|---|---|---|---|
| RGS-1 | RGS-2 | RGS-3 | RGS-4 | RGS-5 | RGS-6 | RGS-7 | |
| Growth Rate | Positive/GNA-1 | — | Positive/GNA-1 | — | — | — | — |
| Aerial Hyphae Height | — | Negative | Negative | Negative | Negative | Negative | Negative |
| Macroconidia Abundance | Positive/GNA-1 | Positive/GNA-3 | Positive/GNA-1 | Positive/GNA-1 | Positive/GNA-1 | Positive/GNA-1 | Positive/GNA-1 |
| Sexual Development | Positive/GNA-3 | Positive/GNA-1 | — | — | — | — | — |
| Avicel Utilization (2 days) | — | Positive/GNA-3 | — | Positive/GNA-3 | Positive/GNA-3 | Positive/GNA-3 | Positive/GNA-3 |
| Culture Biomass in Avicel (3 days) | Negative | Positive/GNA-3 | Positive/GNA-3 | — | — | — | Positive/GNA-3 |
| Glucose Release Cellulase Activity | Negative/GNA-1 GNA-2 | Positive/GNA-3 | Positive/GNA-3 | Negative/GNA-1 GNA-2 | — | — | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabrera, I.E.; Oza, Y.; Carrillo, A.J.; Collier, L.A.; Wright, S.J.; Li, L.; Borkovich, K.A. Regulator of G Protein Signaling Proteins Control Growth, Development and Cellulase Production in Neurospora crassa. J. Fungi 2022, 8, 1076. https://doi.org/10.3390/jof8101076
Cabrera IE, Oza Y, Carrillo AJ, Collier LA, Wright SJ, Li L, Borkovich KA. Regulator of G Protein Signaling Proteins Control Growth, Development and Cellulase Production in Neurospora crassa. Journal of Fungi. 2022; 8(10):1076. https://doi.org/10.3390/jof8101076
Chicago/Turabian StyleCabrera, Ilva E., Yagna Oza, Alexander J. Carrillo, Logan A. Collier, Sara J. Wright, Liande Li, and Katherine A. Borkovich. 2022. "Regulator of G Protein Signaling Proteins Control Growth, Development and Cellulase Production in Neurospora crassa" Journal of Fungi 8, no. 10: 1076. https://doi.org/10.3390/jof8101076