
Voriconazole Treatment Induces a Conserved Sterol/Pleiotropic Drug Resistance Regulatory Network, including an Alternative Ergosterol Biosynthesis Pathway, in the Clinically Important FSSC Species, Fusarium keratoplasticum

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolate and Culture Conditions
2.2. Total RNA Extraction and Sequencing
2.3. De Novo Transcriptome Assembly and Analysis of Differentially Expressed Genes (DEGs)
2.4. Search for Homologs of F. keratoplasticum Genes in Other Fungi
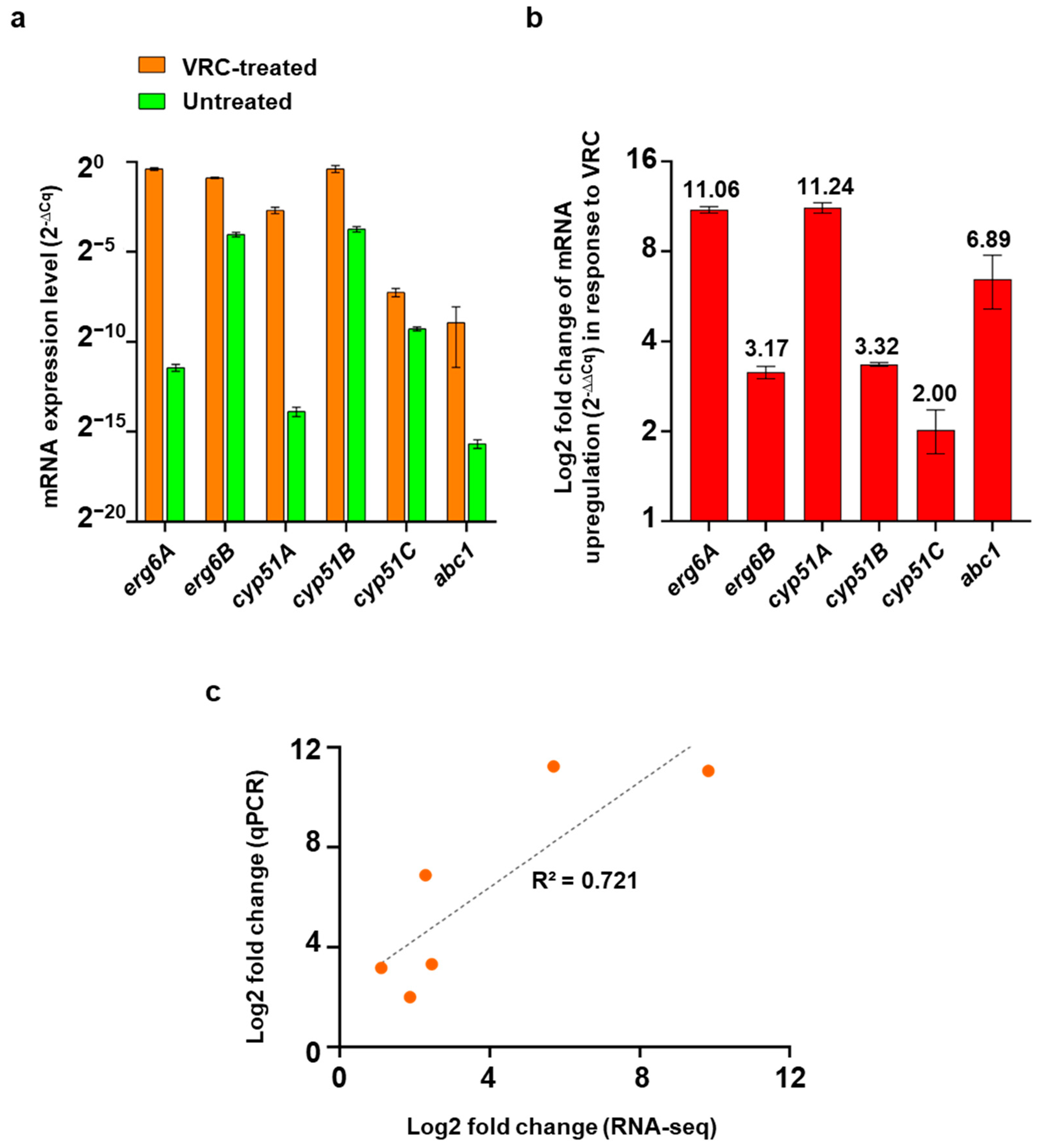
2.5. Validation of RNA-Seq DEGs Using qPCR
3. Results
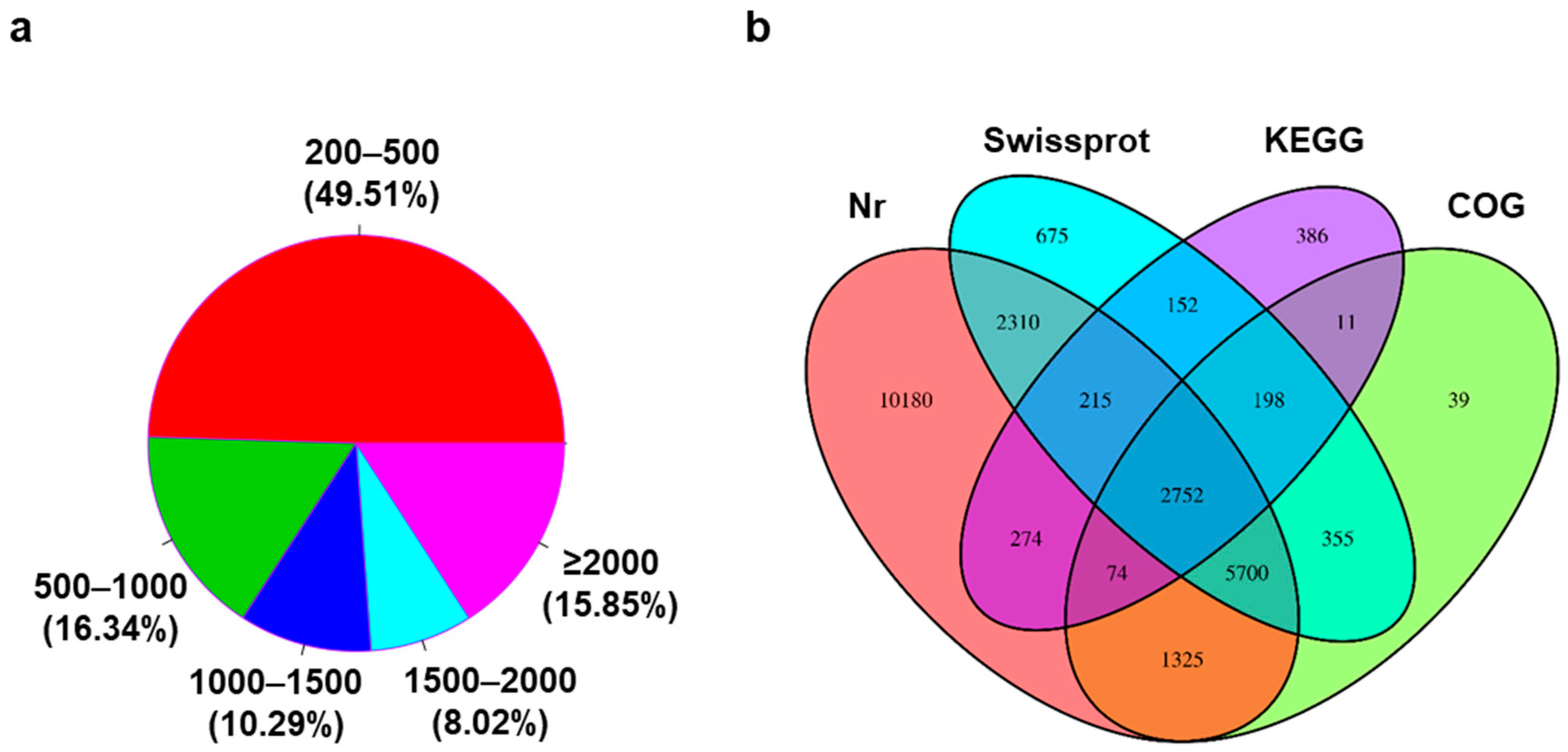
3.1. De Novo Transcriptome Assembly and Annotation of Transcripts
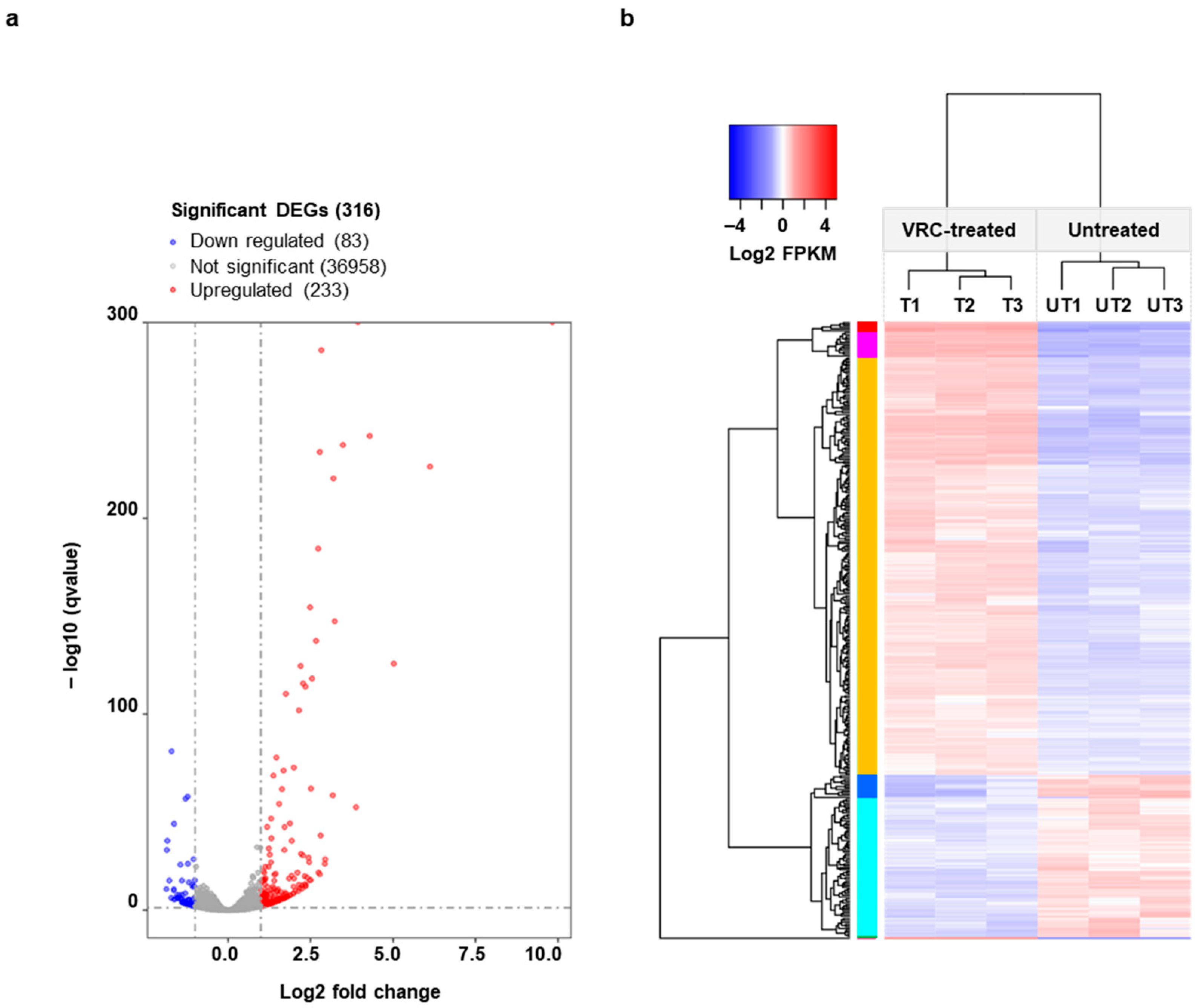
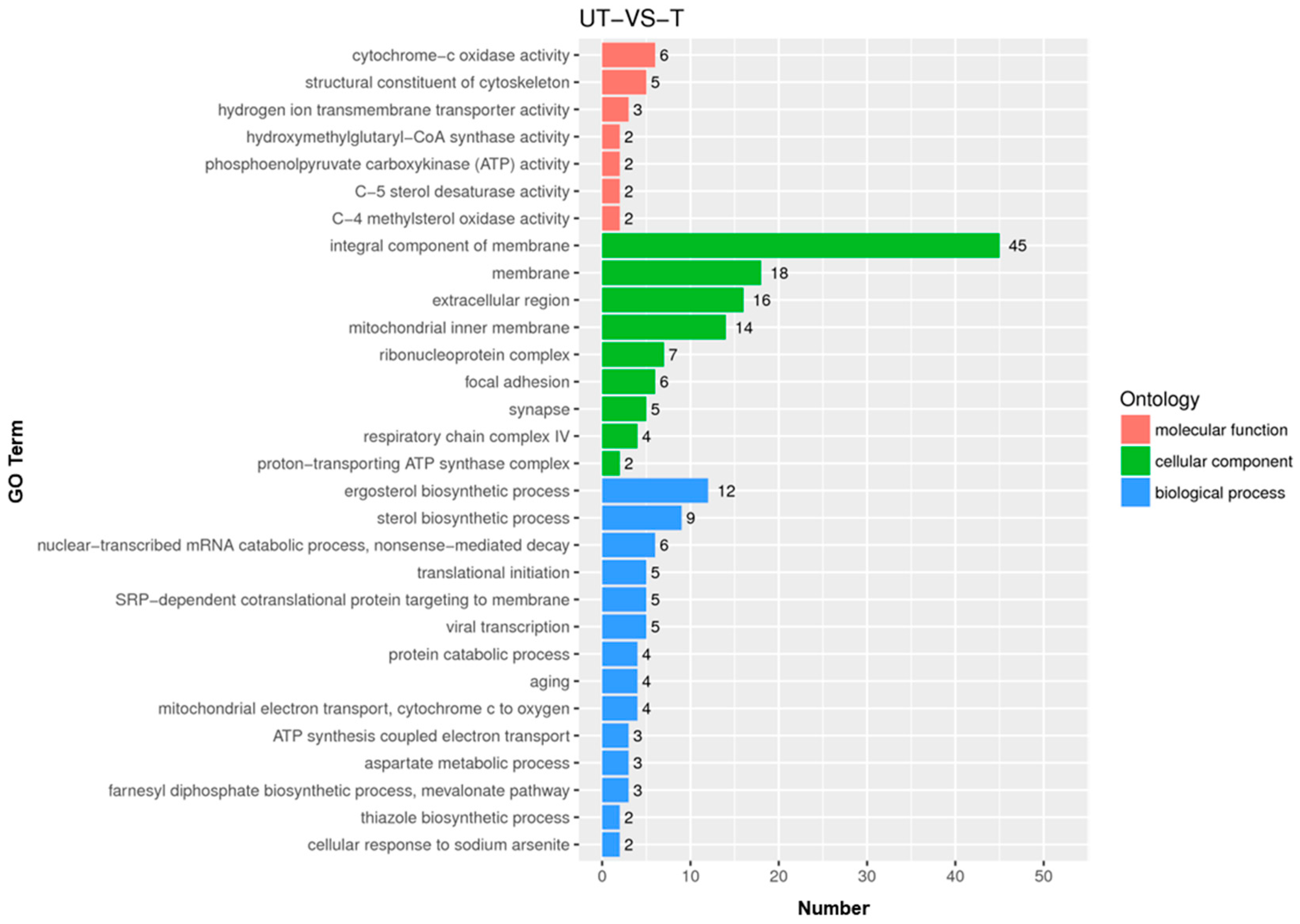
3.2. Genes Differentially Expressed in the Presence of VRC
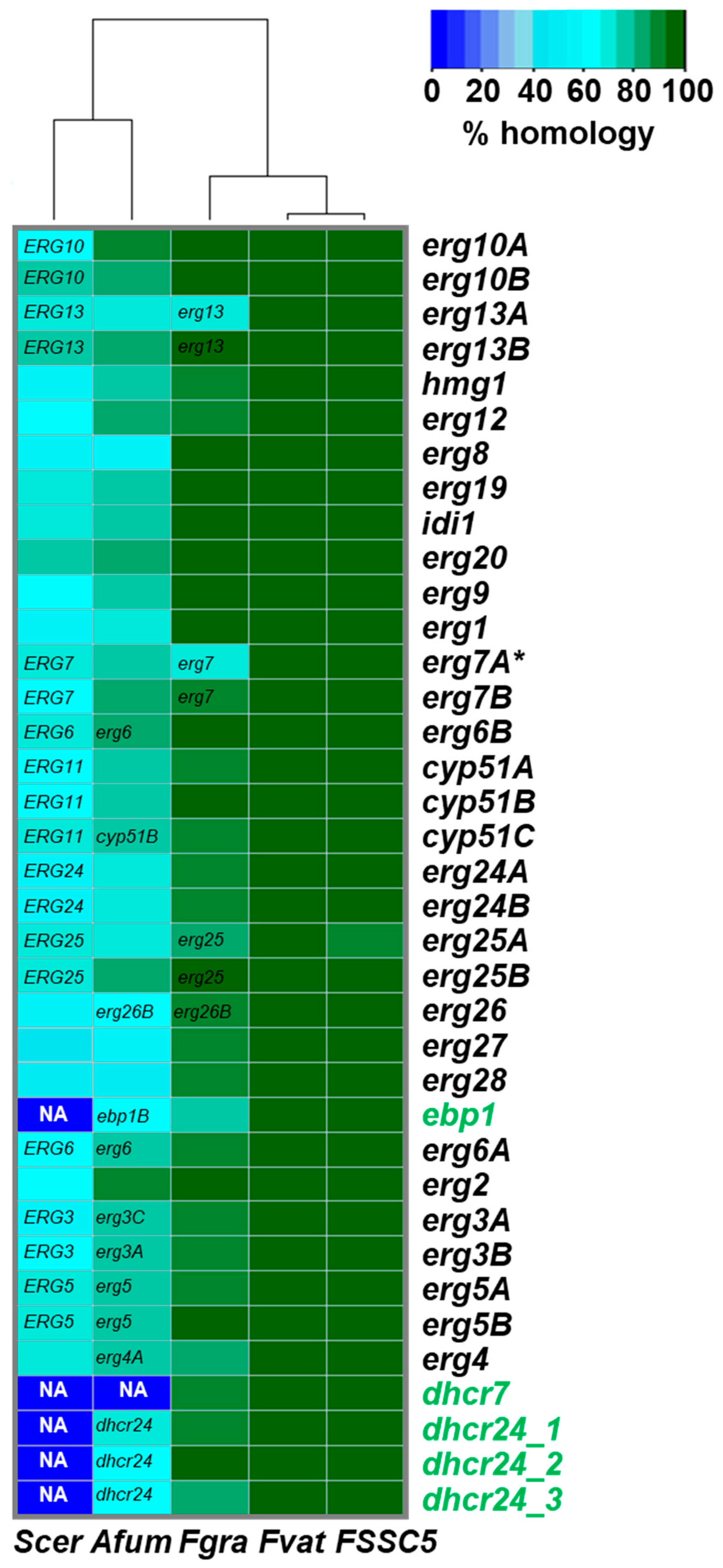
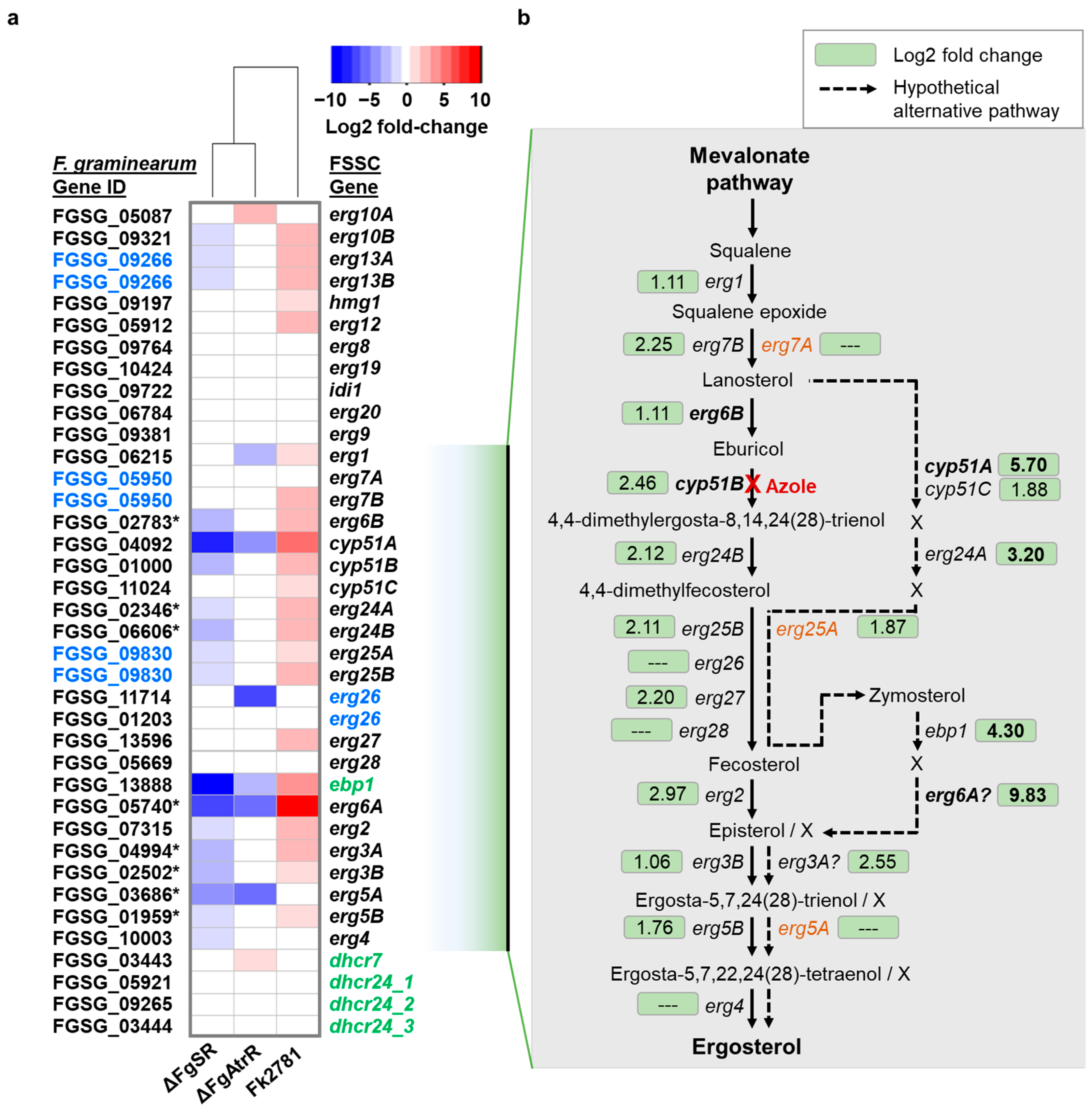
3.3. Inventory of All F. keratoplasticum Sterol Biosynthesis Gene Transcripts and Their Orthologs in F. solani, F. vanettenii, F. graminearum, A. fumigatus and S. cerevisiae
3.4. Identification of F. keratoplasticum ABC Transporters and Transcription Factors Differentially Expressed in VRC Treated Cells
3.5. Transcriptional Response of F. keratoplasticum Cells to VRC Exposure Suggests the Presence of an Alternative Route in the Ergosterol Biosynthesis Pathway
3.6. Differential Expression of ABC Transporters and Transcription Factors in Response to VRC
3.7. Correlation between Gene Expression Levels Measured by qPCR and RNA-Seq
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Coleman, J.J. The Fusarium solani species complex: Ubiquitous pathogens of agricultural importance. Mol. Plant Pathol. 2016, 17, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Gardiner, D.M. Transcriptomics of cereal–Fusarium graminearum interactions: What we have learned so far. Mol. Plant Pathol. 2018, 19, 764–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbaniak, C.; Massa, G.; Hummerick, M.; Khodadad, C.; Schuerger, A.; Venkateswaran, K. Draft genome sequences of two Fusarium oxysporum isolates cultured from infected Zinnia hybrida plants grown on the International Space Station. Genome Announc. 2018, 6, e00326-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nucci, M.; Anaissie, E. Fusarium infections in immunocompromised patients. Clin. Microbiol. Rev. 2007, 20, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Horn, D.L.; Freifeld, A.G.; Schuster, M.G.; Azie, N.E.; Franks, B.; Kauffman, C.A. Treatment and outcomes of invasive fusariosis: Review of 65 cases from the PATH Alliance registry. Mycoses 2014, 57, 652–658. [Google Scholar] [CrossRef] [Green Version]
- Nucci, M.; Anaissie, E. Cutaneous infection by Fusarium species in healthy and immunocompromised hosts: Implications for diagnosis and management. Clin. Infect. Dis. 2002, 35, 909–920. [Google Scholar] [CrossRef] [Green Version]
- Espinel-Ingroff, A.; Colombo, A.L.; Cordoba, S.; Dufresne, P.J.; Fuller, J.; Ghannoum, M.; Gonzalez, G.M.; Guarro, J.; Kidd, S.E.; Meis, J.F.; et al. International evaluation of MIC distributions and epidemiological cutoff value (ECV) definitions for Fusarium species identified by molecular methods for the CLSI broth microdilution method. Antimicrob. Agents Chemother. 2016, 60, 1079–1084. [Google Scholar] [CrossRef] [Green Version]
- Blaize, M.; Normand, A.C.; Imbert, S.; Al-Hatmi, A.M.S.; Chryssanthou, E.; Cassaing, S.; Schuttler, C.; Hasseine, L.; Mahinc, C.; Costa, D.; et al. Antifungal susceptibility of 182 Fusarium species isolates from 20 European centers: Comparison between EUCAST and gradient concentration strip methods. Antimicrob. Agents Chemother. 2021, 65, e0149521. [Google Scholar] [CrossRef]
- Stempel, J.M.; Hammond, S.P.; Sutton, D.A.; Weiser, L.M.; Marty, F.M. Invasive fusariosis in the voriconazole era: Single-center 13-year experience. Open Forum Infect. Dis. 2015, 2, ofv099. [Google Scholar] [CrossRef] [Green Version]
- Lortholary, O.; Obenga, G.; Biswas, P.; Caillot, D.; Chachaty, E.; Bienvenu, A.L.; Cornet, M.; Greene, J.; Herbrecht, R.; Lacroix, C.; et al. International retrospective analysis of 73 cases of invasive fusariosis treated with voriconazole. Antimicrob. Agents Chemother. 2010, 54, 4446–4450. [Google Scholar] [CrossRef]
- Hoenigl, M.; Salmanton-García, J.; Walsh, T.J.; Nucci, M.; Neoh, C.F.; Jenks, J.D.; Lackner, M.; Sprute, R.; Al-Hatmi, A.M.S.; Bassetti, M.; et al. Global guideline for the diagnosis and management of rare mould infections: An initiative of the European Confederation of Medical Mycology in cooperation with the International Society for Human and Animal Mycology and the American Society for Microbiology. Lancet Infect. Dis. 2021, 21, e246–e257. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Endo, T.; Hashimoto, D.; Saga, T.; Ara, T.; Ogasawara, R.; Yasumoto, A.; Ibata, M.; Takahata, M.; Shigematsu, A.; et al. Disseminated fusariosis emerged from prolonged local genital infection after cord blood transplantation. J. Infect. Chemother. 2018, 24, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.M.; Silveira, A.R.; Betania, C.A.; Macedo, D.P.; Neves, R.P. Disseminated fusariosis secondary to neuroblastoma with fatal outcome. Mycopathologia 2013, 176, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Esnakula, A.K.; Summers, I.; Naab, T.J. Fatal disseminated Fusarium infection in a human immunodeficiency virus positive patient. Case Rep. Infect. Dis. 2013, 2013, 379320. [Google Scholar] [CrossRef] [Green Version]
- Nucci, M.; Anaissie, E.J.; Queiroz-Telles, F.; Martins, C.A.; Trabasso, P.; Solza, C.; Mangini, C.; Simoes, B.P.; Colombo, A.L.; Vaz, J. Outcome predictors of 84 patients with hematologic malignancies and Fusarium infection. Cancer 2003, 98, 315–319. [Google Scholar] [CrossRef]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Bruhl, C.A.; Imfeld, G.; Knabel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmuller, A.; et al. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef]
- Meis, J.F.; Chowdhary, A.; Rhodes, J.L.; Fisher, M.C.; Verweij, P.E. Clinical implications of globally emerging azole resistance in Aspergillus fumigatus. Philos. Trans. R Soc. Lond. B Biol. Sci. 2016, 371, 20150460. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Emergence of azole-resistant Aspergillus fumigatus strains due to agricultural azole use creates an increasing threat to human health. PLoS Pathog. 2013, 9, e1003633. [Google Scholar] [CrossRef]
- Cao, D.; Wang, F.; Yu, S.; Dong, S.; Wu, R.; Cui, N.; Ren, J.; Xu, T.; Wang, S.; Wang, M.; et al. Prevalence of azole-resistant Aspergillus fumigatus is highly associated with azole fungicide residues in the fields. Environ. Sci. Technol. 2021, 55, 3041–3049. [Google Scholar] [CrossRef]
- Zhang, J.; van den Heuvel, J.; Debets, A.J.M.; Verweij, P.E.; Melchers, W.J.G.; Zwaan, B.J.; Schoustra, S.E. Evolution of cross-resistance to medical triazoles in Aspergillus fumigatus through selection pressure of environmental fungicides. Proc. Biol. Sci. 2017, 284, 20170635. [Google Scholar] [CrossRef]
- Kang, S.E.; Sumabat, L.G.; Melie, T.; Mangum, B.; Momany, M.; Brewer, M.T. Evidence for the agricultural origin of resistance to multiple antimicrobials in Aspergillus fumigatus, a fungal pathogen of humans. G3 2022, 12, jkab427. [Google Scholar] [CrossRef] [PubMed]
- Snelders, E.; Huis In ’t Veld, R.A.; Rijs, A.J.; Kema, G.H.; Melchers, W.J.; Verweij, P.E. Possible environmental origin of resistance of Aspergillus fumigatus to medical triazoles. Appl. Environ. Microbiol. 2009, 75, 4053–4057. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Sharma, C.; Hagen, F.; Meis, J.F. Exploring azole antifungal drug resistance in Aspergillus fumigatus with special reference to resistance mechanisms. Future Microbiol. 2014, 9, 697–711. [Google Scholar] [CrossRef] [PubMed]
- Otu, A.; Osaigbovo, I.; Orefuwa, E.; Ebenso, B.; Ojumu, T. Collaborative One Health approaches can mitigate increasing azole-resistant Aspergillus fumigatus in Africa. Lancet Microbe 2021, 2, e490–e491. [Google Scholar] [CrossRef]
- James, J.E.; Lamping, E.; Santhanam, J.; Milne, T.J.; Abd Razak, M.F.; Zakaria, L.; Cannon, R.D. A 23 bp cyp51A promoter deletion associated with voriconazole resistance in clinical and environmental isolates of Neocosmospora keratoplastica. Front. Microbiol. 2020, 11, 272. [Google Scholar] [CrossRef]
- Tava, V.; Prigitano, A.; Cortesi, P.; Esposto, M.C.; Pasquali, M. Fusarium musae from diseased bananas and human patients: Susceptibility to fungicides used in clinical and agricultural settings. J. Fungi 2021, 7, 784. [Google Scholar] [CrossRef] [PubMed]
- Bard, M.; Lees, N.D.; Turi, T.; Craft, D.; Cofrin, L.; Barbuch, R.; Koegel, C.; Loper, J.C. Sterol synthesis and viability of Erg11 (cytochrome P450 lanosterol demethylase) mutations in Saccharomyces cerevisiae and Candida albicans. Lipids 1993, 28, 963–967. [Google Scholar] [CrossRef]
- Ghannoum, M.A.; Rice, L.B. Antifungal agents: Mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [CrossRef] [Green Version]
- Bien, C.M.; Espenshade, P.J. Sterol regulatory element binding proteins in fungi: Hypoxic transcription factors linked to pathogenesis. Eukaryot. Cell 2010, 9, 352–359. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.S.; Goldstein, J.L. The SREBP pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997, 89, 331–340. [Google Scholar] [CrossRef]
- Yang, H.; Tong, J.; Lee, C.W.; Ha, S.; Eom, S.H.; Im, Y.J. Structural mechanism of ergosterol regulation by fungal sterol transcription factor Upc2. Nat. Commun. 2015, 6, 6129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacPherson, S.; Akache, B.; Weber, S.; De Deken, X.; Raymond, M.; Turcotte, B. Candida albicans zinc cluster protein Upc2p confers resistance to antifungal drugs and is an activator of ergosterol biosynthetic genes. Antimicrob. Agents Chemother. 2005, 49, 1745–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Jian, Y.; Chen, Y.; Kistler, H.C.; He, P.; Ma, Z.; Yin, Y. A phosphorylated transcription factor regulates sterol biosynthesis in Fusarium graminearum. Nat. Commun. 2019, 10, 1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Stamnes, M.; Thomas, G.H.; Liu, H.; Hagiwara, D.; Gomi, K.; Filler, S.G.; Moye-Rowley, W.S. AtrR is an essential determinant of azole resistance in Aspergillus fumigatus. mBio 2019, 10, e02563-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, D.; Miura, D.; Shimizu, K.; Paul, S.; Ohba, A.; Gonoi, T.; Watanabe, A.; Kamei, K.; Shintani, T.; Moye-Rowley, W.S.; et al. A novel Zn2-Cys6 transcription factor AtrR plays a key role in an azole resistance mechanism of Aspergillus fumigatus by co-regulating cyp51A and cdr1B expressions. PLoS Pathog. 2017, 13, e1006096. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Sun, H.; Li, J.; Ju, C.; Huang, J. The transcription factor FgAtrR regulates asexual and sexual development, virulence, and DON production and contributes to intrinsic resistance to azole fungicides in Fusarium graminearum. Biology 2022, 11, 326. [Google Scholar] [CrossRef]
- Mellado, E.; Diaz-Guerra, T.M.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Identification of two different 14-alpha sterol demethylase-related genes (CYP51A and CYP51B) in Aspergillus fumigatus and other Aspergillus species. J. Clin. Microbiol. 2001, 39, 2431–2438. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Urban, M.; Parker, J.E.; Brewer, H.C.; Kelly, S.L.; Hammond-Kosack, K.E.; Fraaije, B.A.; Liu, X.; Cools, H.J. Characterization of the sterol 14alpha-demethylases of Fusarium graminearum identifies a novel genus-specific CYP51 function. New Phytol. 2013, 198, 821–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucio, J.; Gonzalez-Jimenez, I.; Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Pelaez, T.; Alcazar-Fuoli, L.; Mellado, E. Point mutations in the 14-alpha sterol demethylase Cyp51A or Cyp51C could contribute to azole resistance in Aspergillus flavus. Genes 2020, 11, 1217. [Google Scholar] [CrossRef]
- Warrilow, A.G.; Melo, N.; Martel, C.M.; Parker, J.E.; Nes, W.D.; Kelly, S.L.; Kelly, D.E. Expression, purification, and characterization of Aspergillus fumigatus sterol 14-alpha demethylase (Cyp51) isoenzymes A and B. Antimicrob. Agents Chemother. 2010, 54, 4225–4234. [Google Scholar] [CrossRef]
- Bean, T.P.; Cools, H.J.; Lucas, J.A.; Hawkins, N.D.; Ward, J.L.; Shaw, M.W.; Fraaije, B.A. Sterol content analysis suggests altered eburicol 14alpha-demethylase (Cyp51) activity in isolates of Mycosphaerella graminicola adapted to azole fungicides. FEMS Microbiol. Lett. 2009, 296, 266–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcazar-Fuoli, L.; Mellado, E.; Garcia-Effron, G.; Lopez, J.F.; Grimalt, J.O.; Cuenca-Estrella, J.M.; Rodriguez-Tudela, J.L. Ergosterol biosynthesis pathway in Aspergillus fumigatus. Steroids 2008, 73, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Alcazar-Fuoli, L.; Mellado, E. Ergosterol biosynthesis in Aspergillus fumigatus: Its relevance as an antifungal target and role in antifungal drug resistance. Front. Microbiol. 2012, 3, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Esquivel, B.D.; White, T.C. Overexpression or deletion of ergosterol biosynthesis genes alters doubling time, response to stress agents, and drug susceptibility in Saccharomyces cerevisiae. mBio 2018, 9, e01291-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Diekema, D.; Moye-Rowley, W.S. Contributions of both ATP-binding cassette transporter and Cyp51A proteins are essential for azole resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2017, 61, e02748-16. [Google Scholar] [CrossRef] [Green Version]
- James, J.E.; Lamping, E.; Santhanam, J.; Cannon, R.D. PDR transporter ABC1 is involved in the innate azole resistance of the human fungal pathogen Fusarium keratoplasticum. Front. Microbiol. 2021, 12, 673206. [Google Scholar] [CrossRef]
- James, J.E.; Santhanam, J.; Zakaria, L.; Mamat Rusli, N.; Abu Bakar, M.; Suetrong, S.; Sakayaroj, J.; Abdul Razak, M.F.; Lamping, E.; Cannon, R.D. Morphology, phenotype, and molecular identification of clinical and environmental Fusarium solani species complex isolates from Malaysia. J. Fungi 2022, 8, 845. [Google Scholar] [CrossRef]
- Andrew, S. FastQC: A Quality Control Tool for High Throughput Sequence Data, 0.11.4; Babraham Institute: Cambridge, UK, 2015.
- Martin, M. Cutadapt: Removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J. TransDecoder: Find Coding Regions Within Transcripts, 3.0.0. 2016.
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. gplots: Various R Programming Tools for Plotting Data, 3.1.3; R package version: Vienna, Austria, 2022.
- R-core-team. R: A Language and Environment for Statistical Computing, 4.2.0; R Foundation for Statistical Computing: Vienna, Austria, 2022.
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesny, F.; Miyauchi, S.; Thiergart, T.; Pickel, B.; Atanasova, L.; Karlsson, M.; Huttel, B.; Barry, K.W.; Haridas, S.; Chen, C.; et al. Genetic determinants of endophytism in the Arabidopsis root mycobiome. Nat. Commun. 2021, 12, 7227. [Google Scholar] [CrossRef]
- Coleman, J.J.; Rounsley, S.D.; Rodriguez-Carres, M.; Kuo, A.; Wasmann, C.C.; Grimwood, J.; Schmutz, J.; Taga, M.; White, G.J.; Zhou, S.; et al. The genome of Nectria haematococca: Contribution of supernumerary chromosomes to gene expansion. PLoS Genet. 2009, 5, e1000618. [Google Scholar] [CrossRef] [Green Version]
- Cuomo, C.A.; Guldener, U.; Xu, J.R.; Trail, F.; Turgeon, B.G.; Di Pietro, A.; Walton, J.D.; Ma, L.J.; Baker, S.E.; Rep, M.; et al. The Fusarium graminearum genome reveals a link between localized polymorphism and pathogen specialization. Science 2007, 317, 1400–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nierman, W.C.; Pain, A.; Anderson, M.J.; Wortman, J.R.; Kim, H.S.; Arroyo, J.; Berriman, M.; Abe, K.; Archer, D.B.; Bermejo, C.; et al. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 2005, 438, 1151–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 genes. Science 1996, 274, 546–547. [Google Scholar] [CrossRef] [Green Version]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef]
- Zhang, T.; Yuan, D.; Xie, J.; Lei, Y.; Li, J.; Fang, G.; Tian, L.; Liu, J.; Cui, Y.; Zhang, M.; et al. Evolution of the cholesterol biosynthesis pathway in animals. Mol. Biol. Evol. 2019, 36, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Silve, S.; Dupuy, P.H.; Labit-Lebouteiller, C.; Kaghad, M.; Chalon, P.; Rahier, A.; Taton, M.; Lupker, J.; Shire, D.; Loison, G. Emopamil-binding protein, a mammalian protein that binds a series of structurally diverse neuroprotective agents, exhibits delta8-delta7 sterol isomerase activity in yeast. J. Biol. Chem. 1996, 271, 22434–22440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, S.H.; Lee, J.N.; Fitzky, B.U.; Seong, J.; Paik, Y.K. Cholesterol biosynthesis from lanosterol. Molecular cloning, tissue distribution, expression, chromosomal localization, and regulation of rat 7-dehydrocholesterol reductase, a Smith-Lemli-Opitz syndrome-related protein. J. Biol. Chem. 1999, 274, 14624–14631. [Google Scholar] [CrossRef] [Green Version]
- Becher, R.; Weihmann, F.; Deising, H.B.; Wirsel, S.G. Development of a novel multiplex DNA microarray for Fusarium graminearum and analysis of azole fungicide responses. BMC Genom. 2011, 12, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamping, E.; Baret, P.V.; Holmes, A.R.; Monk, B.C.; Goffeau, A.; Cannon, R.D. Fungal PDR transporters: Phylogeny, topology, motifs and function. Fungal Genet. Biol. 2010, 47, 127–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, J.J.; White, G.J.; Rodriguez-Carres, M.; Vanetten, H.D. An ABC transporter and a cytochrome P450 of Nectria haematococca MPVI are virulence factors on pea and are the major tolerance mechanisms to the phytoalexin pisatin. Mol. Plant Microbe Interact. 2011, 24, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou Ammar, G.; Tryono, R.; Doll, K.; Karlovsky, P.; Deising, H.B.; Wirsel, S.G. Identification of ABC transporter genes of Fusarium graminearum with roles in azole tolerance and/or virulence. PLoS ONE 2013, 8, e79042. [Google Scholar] [CrossRef] [Green Version]
- Hellin, P.; King, R.; Urban, M.; Hammond-Kosack, K.E.; Legreve, A. The adaptation of Fusarium culmorum to DMI fungicides is mediated by major transcriptome modifications in response to azole fungicide, including the overexpression of a PDR transporter (FcABC1). Front. Microbiol. 2018, 9, 1385. [Google Scholar] [CrossRef]
- Fraczek, M.G.; Bromley, M.; Buied, A.; Moore, C.B.; Rajendran, R.; Rautemaa, R.; Ramage, G.; Denning, D.W.; Bowyer, P. The cdr1B efflux transporter is associated with non-cyp51a-mediated itraconazole resistance in Aspergillus fumigatus. J. Antimicrob. Chemother. 2013, 68, 1486–1496. [Google Scholar] [CrossRef]
- Willger, S.D.; Puttikamonkul, S.; Kim, K.H.; Burritt, J.B.; Grahl, N.; Metzler, L.J.; Barbuch, R.; Bard, M.; Lawrence, C.B.; Cramer, R.A., Jr. A sterol-regulatory element binding protein is required for cell polarity, hypoxia adaptation, azole drug resistance, and virulence in Aspergillus fumigatus. PLoS Pathog. 2008, 4, e1000200. [Google Scholar] [CrossRef]
- Todd, B.L.; Stewart, E.V.; Burg, J.S.; Hughes, A.L.; Espenshade, P.J. Sterol regulatory element binding protein is a principal regulator of anaerobic gene expression in fission yeast. Mol. Cell. Biol. 2006, 26, 2817–2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Alonso, L.; Romero, A.M.; Martinez-Pastor, M.T.; Puig, S. Iron regulatory mechanisms in Saccharomyces cerevisiae. Front. Microbiol. 2020, 11, 582830. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Moebius, F.F.; Striessnig, J.; Glossmann, H. The mysteries of sigma receptors: New family members reveal a role in cholesterol synthesis. Trends Pharmacol. Sci. 1997, 18, 67–70. [Google Scholar] [CrossRef]
- Sonawane, P.D.; Pollier, J.; Panda, S.; Szymanski, J.; Massalha, H.; Yona, M.; Unger, T.; Malitsky, S.; Arendt, P.; Pauwels, L.; et al. Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. Nat. Plants 2016, 3, 16205. [Google Scholar] [CrossRef]
- Long, N.; Xu, X.; Zeng, Q.; Sang, H.; Lu, L. Erg4A and Erg4B are required for conidiation and azole resistance via regulation of ergosterol biosynthesis in Aspergillus fumigatus. Appl. Environ. Microbiol. 2017, 83, e02924-16. [Google Scholar] [CrossRef] [Green Version]
- Souza, C.M.; Schwabe, T.M.; Pichler, H.; Ploier, B.; Leitner, E.; Guan, X.L.; Wenk, M.R.; Riezman, I.; Riezman, H. A stable yeast strain efficiently producing cholesterol instead of ergosterol is functional for tryptophan uptake, but not weak organic acid resistance. Metab. Eng. 2011, 13, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Weete, J.D.; Abril, M.; Blackwell, M. Phylogenetic distribution of fungal sterols. PLoS ONE 2010, 5, e10899. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F. keratoplasticum 2781 | JGI Protein ID (% Identity/% Homology) 1, 2 | |||||
|---|---|---|---|---|---|---|
| Gene | Fusarium solani FSSC 5 | F. vanettenii 77-13-4 | F. graminearum PH-1 | A. fumigatus Af293 | S. cerevisiae S288C | |
| ABC transporters | ||||||
| ABCB-1 | DN33072 | 511443 (100/100) | 96330 (98/99) | 6063 (80/88) | 3538 (54/72) | 3617 (28/47) |
| ABCB-2 | DN3265 | 543122 (100/100) | 92751 (99/100) | 7949 (75/85) | 8558 (47/66) | 6078 (28/48) |
| ABCC-yor1 | DN5825 | 393338 (100/100) | 63546 (98/100) | 8483 (85/92) | 5371 (59/74) | 2864 (37/56) |
| ABCG-abc1 | DN16847 | 537064 (100/100) | 63187 (98/99) | 3418 (81/89) | 1300 (60/73) | 1445 (46/64) |
| ABCG-abc3 | DN36936 | 417245 (100/100) | 35467 (99/100) | 9970 (86/91) | 1300 (63/76) | 5806 (49/67) |
| ABCG-H1 | DN21770 | 439905 (100/100) | 35868 (99/100) | 9973 (84/91) | 6736 (64/78) | 6299 (37/56) |
| ABCG-F | DN29261 | 57426 (99/99) | 68948 (99/99) | 12739 (81/90) | 4105 (56/72) | 5566 (38/58) |
| Transcription factors | ||||||
| atrR | DN25544 | 393104 (100/100) | 96223 (98/99) | 7871 (84/90) | 1901 (63/73) | NA |
| moc3 | DN12705 | 489853 (98/99) | 1518 (97/98) | 8308 (66/76) | NA | NA |
| atF | DN14162 | 502182 (96/98) | 84321 (100/100) | 9383 (36/55) | NA | NA |
| SR | DN37099 | 384730 (100/100) | 103092 (99/100) | 1375 (87/93) | NA | NA |
| yap5 | DN16099 | 42136 (99/100) | 74330 (97/98) | 1660 (73/82) | 1198 (46/59) | NA |
| SREBP1 | DN13837 | 554245 (100/100) | 67823 (99/100) | 5412 (84/91) | NA | NA |
| Gene ID | Gene Description 1 | Gene | Log2 Fold Change 2, 3, 4 | Mean FPKM Values | |
|---|---|---|---|---|---|
| VRC-Treated (n = 3) | Untreated (n = 3) | ||||
| Ergosterol biosynthesis 5 | |||||
| DN33281 | Acetyl CoA thiolase | erg10A | −0.41 | 99 | 119 |
| DN17604 | Acetyl CoA thiolase | erg10B | 2.49 | 1165 | 180 |
| DN37027 | HMG CoA synthase | erg13A | 2.44 | 209 | 25 |
| DN8815 | HMG CoA synthase | erg13B | 2.78 | 1199 | 151 |
| DN33169 | HMG CoA reductase | hmg1 | 1.64 | 208 | 59 |
| DN18060 | Mevalonate kinase | erg12 | 2.28 | 110 | 20 |
| DN33074 | Phosphomevalonate kinase | erg8 | * −0.10 | 30 | 29 |
| DN29440 | Mevalonate pyrophosphate kinase | erg19 | * −0.03 | 90 | 83 |
| DN29378 | Isopentenyl pyrophosphate isomerase | idi1 | * −0.16 | 171 | 172 |
| DN10425 | Geranyl pyrophosphate synthase | erg20 | * 0.17 | 20 | 16 |
| DN17806 | Squalene synthase | erg9 | 0.88 | 343 | 166 |
| DN18090 | Squalene epoxidase | erg1 | 1.11 | 78 | 31 |
| DN26925 | Lanosterol synthase | erg7A | * 0.08 | 2 | 0 |
| DN37088 | Lanosterol synthase | erg7B | 2.25 | 59 | 11 |
| DN33059 | Sterol C-24 methyltransferase | erg6B | 1.11 | 1694 | 207 |
| DN25638 | Eburicol 14-α demethylase | cyp51A | 5.70 | 2982 | 2 |
| DN21545 | Eburicol 14-α demethylase | cyp51B | 2.46 | 899 | 104 |
| DN8317 | Claimed not to encode sterol 14-α demethylase 6 | cyp51C | 1.88 | 20 | 5 |
| DN29591 | Sterol C-14 reductase | erg24A | 3.20 | 160 | 15 |
| DN32963 | Sterol C-14 reductase | erg24B | 2.12 | 290 | 58 |
| DN8081 | Sterol C-4 methyl oxidase | erg25A | 1.87 | 2 | 0 |
| DN17697 | Sterol C-4 methyl oxidase | erg25B | 2.11 | 1543 | 248 |
| DN29406 | Sterol C-3 dehydrogenase | erg26 | 0.35 | 82 | 58 |
| DN3112 | 3-keto sterol reductase | erg27 | 2.20 | 149 | 28 |
| DN22157 | Sterol C-4 demethylase | erg28 | * 0.06 | 68 | 58 |
| DN33250 | Sterol C-8 isomerase | ebp1 | 4.30 | 152 | 6 |
| DN32910 | Sterol C-24 methyltransferase | erg6A | 9.83 | 4186 | 3 |
| DN25482 | Sterol C-8 isomerase | erg2 | 2.97 | 1039 | 109 |
| DN15167 | Sterol C-5 desaturase | erg3A | 2.55 | 364 | 52 |
| DN37320 | Sterol C-5 desaturase | erg3B | 1.06 | 103 | 24 |
| DN16841 | Sterol C-22 desaturase | erg5A | * 0.30 | 0 | 0 |
| DN10857 | Sterol C-22 desaturase | erg5B | 1.76 | 575 | 151 |
| DN21765 | Sterol C-24 reductase | erg4 | 0.98 | 170 | 77 |
| DN12846 | C-7 sterol reductase | dhcr7 | NA | 0 | 0 |
| DN10241 | C-24 sterol reductase | dhcr24-1 | NA | 9 | 8 |
| DN18330 | C-24 sterol reductase | dhcr24-2 | NA | 27 | 29 |
| DN14443 | C-24 sterol reductase | dhcr24-3 | NA | 1 | 1 |
| ABC transporters 7 | |||||
| DN3265 | ABCB family | - | 1.38 | 4 | 1.4 |
| DN33072 | ABCB family | - | 1.25 | 20 | 5.4 |
| DN16847 | ABCG family (PDR transporter; cluster B) | abc1 | 2.29 | 1 | 0.05 |
| DN36936 | ABCG family (PDR transporter; cluster B) | abc3 | 1.48 | 21 | 3.3 |
| DN21770 | ABCG family (PDR transporter; cluster H1) | - | 1.24 | 17 | 6.4 |
| DN29261 | ABCG family (PDR transporter; cluster F) | - | –1.86 | 3 | 11.9 |
| Transcription factors | |||||
| DN25544 | Zn2-Cys6 transcription factor 8 | atrR | 2.15 | 34 | 7 |
| DN12705 | Transcriptional regulatory protein | moc3 | 1.32 | 5 | 2 |
| DN14162 | Terreic acid cluster-specific transcription factor | atF | 1.05 | 3 | 1 |
| DN37099 | Ergosterol biosynthesis regulator in fusaria 9 | SR | 0.13 | 33 | 27 |
| DN16099 | AP-1-like transcription factor | yap5 | –1.08 | 0 | 2 |
| DN13837 | Sterol regulatory element-binding protein 1 | SREBP1 | –1.16 | 4 | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, J.E.; Santhanam, J.; Cannon, R.D.; Lamping, E. Voriconazole Treatment Induces a Conserved Sterol/Pleiotropic Drug Resistance Regulatory Network, including an Alternative Ergosterol Biosynthesis Pathway, in the Clinically Important FSSC Species, Fusarium keratoplasticum. J. Fungi 2022, 8, 1070. https://doi.org/10.3390/jof8101070
James JE, Santhanam J, Cannon RD, Lamping E. Voriconazole Treatment Induces a Conserved Sterol/Pleiotropic Drug Resistance Regulatory Network, including an Alternative Ergosterol Biosynthesis Pathway, in the Clinically Important FSSC Species, Fusarium keratoplasticum. Journal of Fungi. 2022; 8(10):1070. https://doi.org/10.3390/jof8101070
Chicago/Turabian StyleJames, Jasper E., Jacinta Santhanam, Richard D. Cannon, and Erwin Lamping. 2022. "Voriconazole Treatment Induces a Conserved Sterol/Pleiotropic Drug Resistance Regulatory Network, including an Alternative Ergosterol Biosynthesis Pathway, in the Clinically Important FSSC Species, Fusarium keratoplasticum" Journal of Fungi 8, no. 10: 1070. https://doi.org/10.3390/jof8101070