
Dry-Season Soil and Co-Cultivated Host Plants Enhanced Propagation of Arbuscular Mycorrhizal Fungal Spores from Sand Dune Vegetation in Trap Culture

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Collection and Nutrient Property Analyses
2.2. Quantification of AMF Spores
2.3. Analysis of Relative Fungal Abundance in Coastal Sand Dunesoil
2.4. Investigation of AMF Species in Coastal Sand Dunesoil
2.5. Experimental Design for Trap Culture in the Greenhouse
2.6. Measurement of Fungal and AMF Colonization in Roots
2.7. Analysis of AMF Communities by T-RFLP
2.8. Statistical Analyses
3. Results
3.1. Soil from the Coastal Sand Dunevegetation Was Infertile
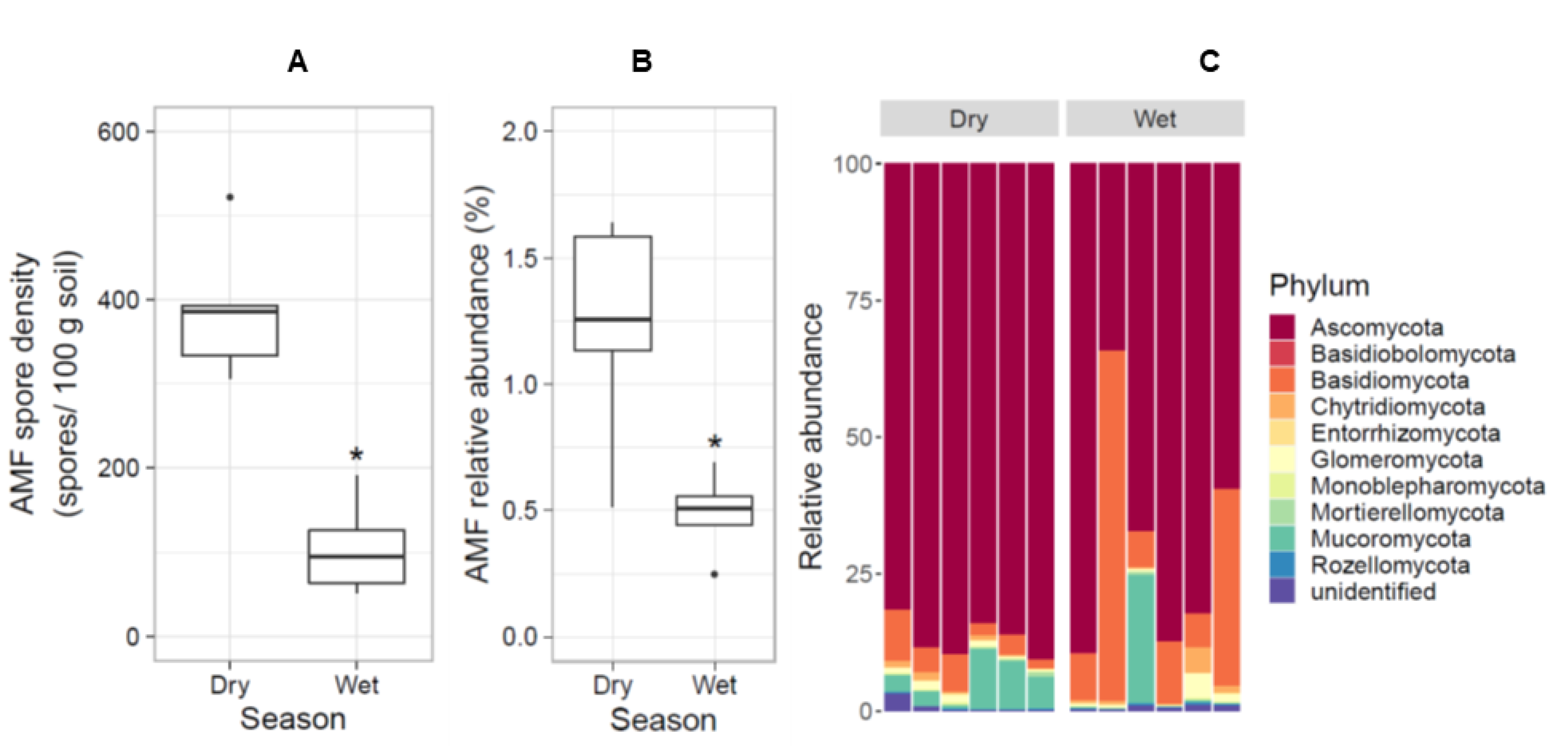
3.2. AMF Was More Abundant in the Dry Season
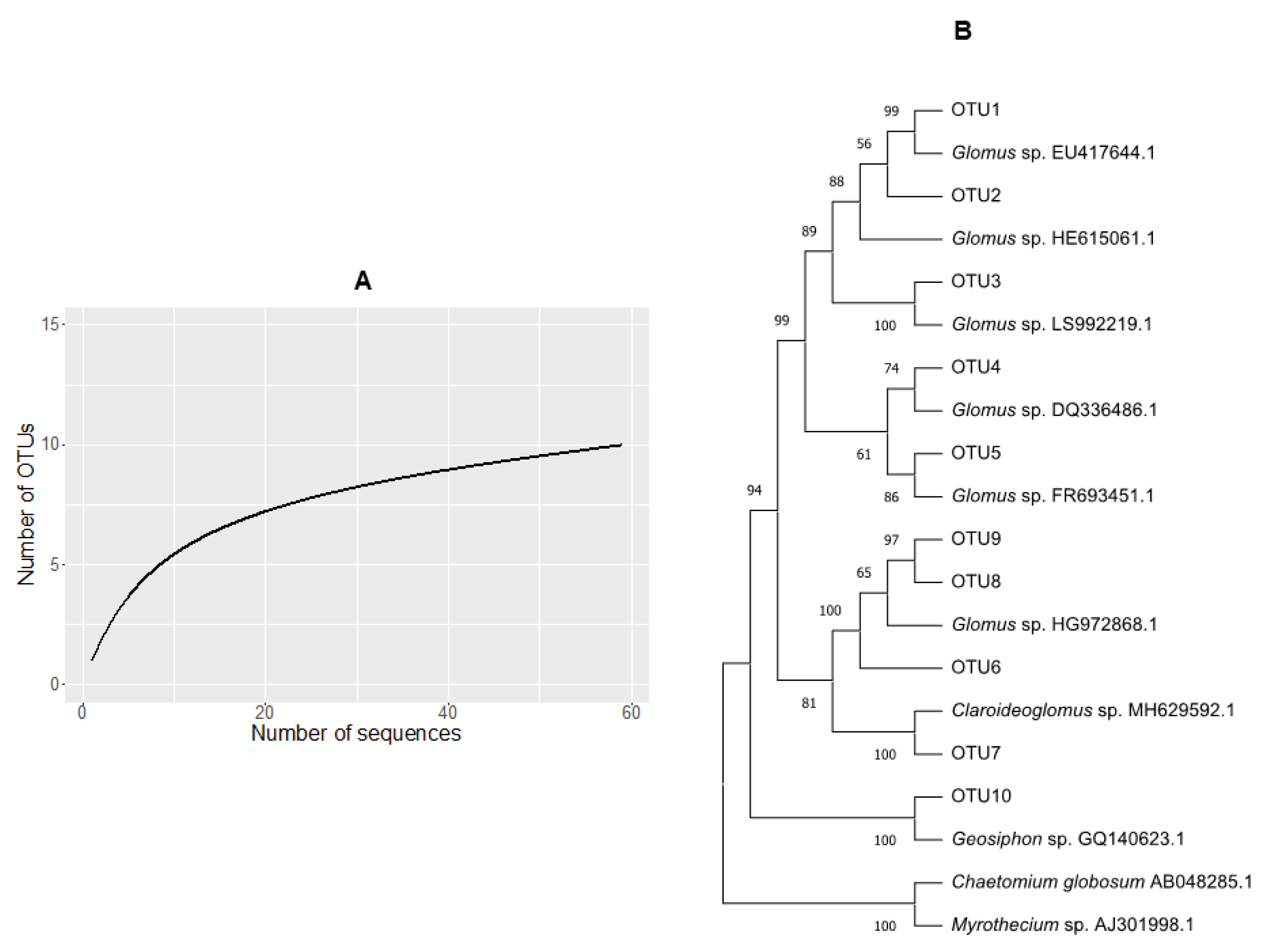
3.3. Glomus Was the Dominant AMF Genus in the Coastal Sand Dune in Dry Season
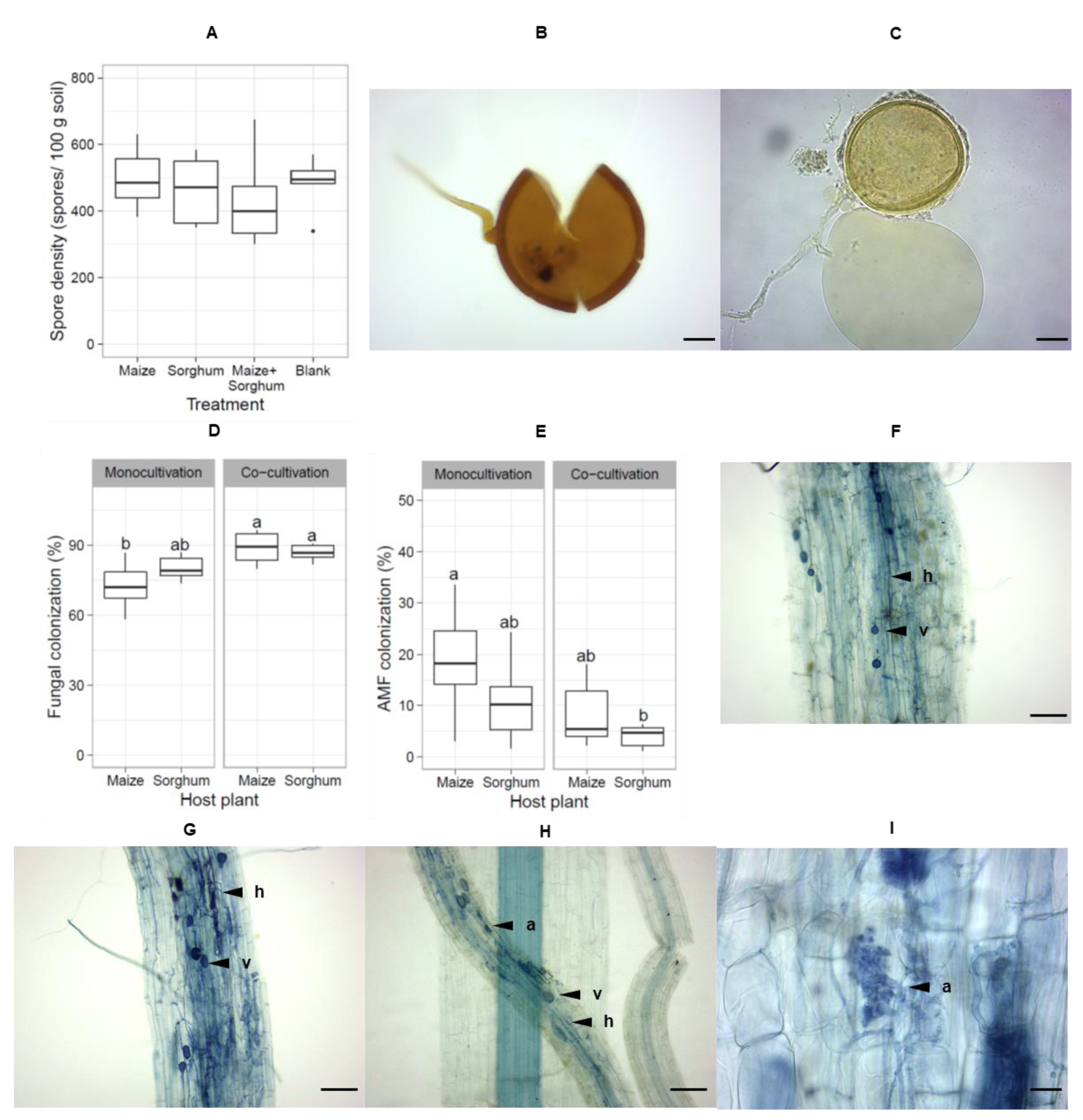
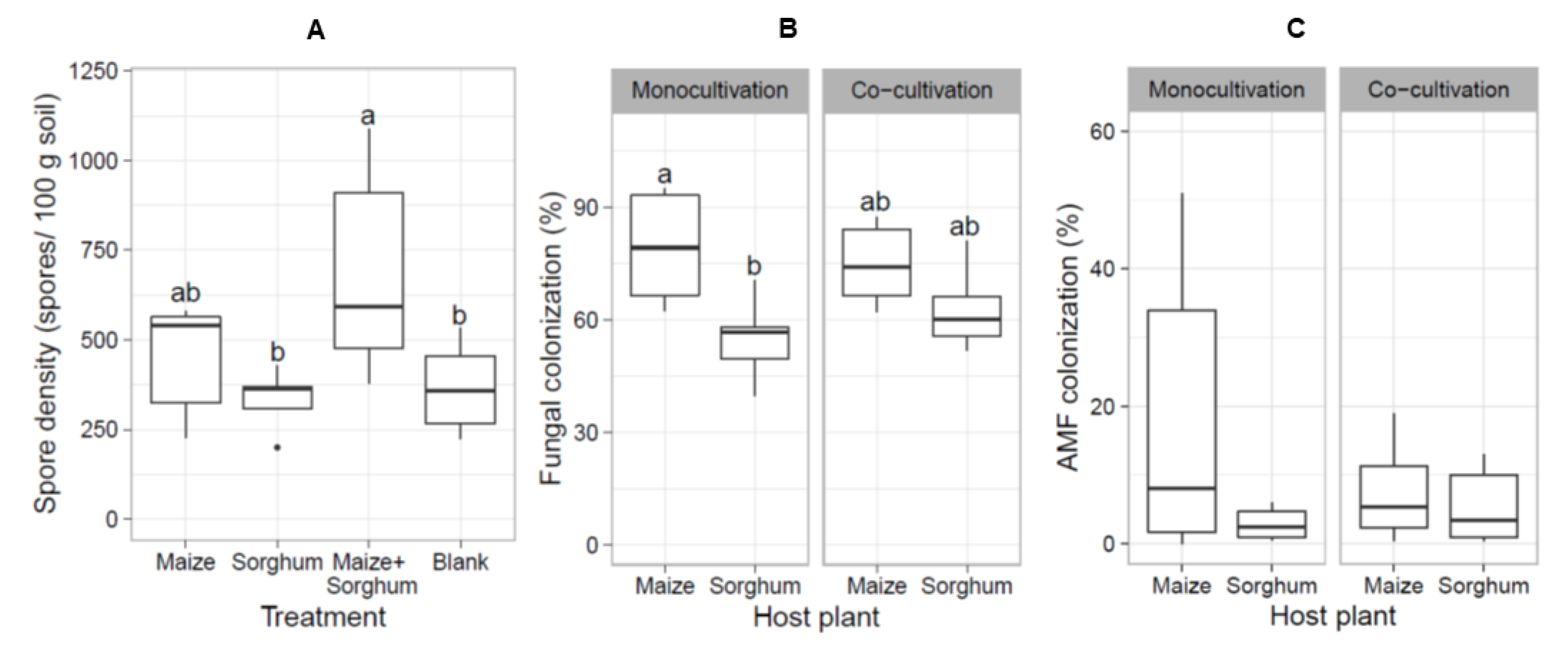
3.4. Co-Cultivation of Maize and Sorghum Improved the Production of AMF Spores
3.5. Trap Culture in the Greenhouse Shifted AMF Communities of Coastal Sand Dune Soil
4. Discussion
4.1. Soil in the Coastal Sand Dune Was Infertile
4.2. AMF Is Abundant in the Coastal Sand Dune Soil in the Dry Season
4.3. Plant Communities Affect AMF Communities in Coastal Sand Dune Ecosystems
4.4. Co-Cultivation of Host Plants Improves the Production of AMF Spores in Trap Culture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bucher, M. Functional biology of plant phosphate uptake at root and mycorrhiza interfaces. New Phytol. 2007, 173, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Keymer, A.; Gutjahr, C. Cross-kingdom lipid transfer in arbuscular mycorrhiza symbiosis and beyond. Curr. Opin. Plant Biol. 2018, 44, 137–144. [Google Scholar] [CrossRef]
- Campo, S.; Martín-Cardoso, H.; Olivé, M.; Pla, E.; Catala-Forner, M.; Martínez-Eixarch, M.; San Segundo, B. Effect of root colonization by arbuscular mycorrhizal fungi on growth, productivity and blast resistance in rice. Rice 2020, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Polispitak, K.; Thongpoem, P.; Singh, H.P.; Cha-um, S. Alleviation of salt stress in upland rice (Oryza sativa L. ssp. indica cv. Leum Pua) using arbuscular mycorrhizal fungi inoculation. Front. Plant Sci. 2020, 11, 348. [Google Scholar] [CrossRef] [Green Version]
- Chareesri, A.; De Deyn, G.B.; Sergeeva, L.; Polthanee, A.; Kuyper, T.W. Increased arbuscular mycorrhizal fungal colonization reduces yield loss of rice (Oryza sativa L.) under drought. Mycorrhiza 2020, 30, 315–328. [Google Scholar] [CrossRef] [Green Version]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular mycorrhizal fungi as natural biofertilizers: Let’s benefit from past successes. Front. Microbiol. 2016, 6, 1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oviatt, P.; Rillig, M.C. Mycorrhizal technologies for an agriculture of the middle. Plants People Planet 2021, 3, 454–461. [Google Scholar] [CrossRef]
- INVAM International Culture Colletion of (Vesicular) Arbuscular Micorrhizal Fungi. Available online: https://invam.ku.edu/ (accessed on 1 June 2021).
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A.; Morsy, M.; Eisen, J.A.; Leach, J.E.; Dangl, J.L. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLOS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, F. Selection of appropriate host plants used in trap culture of arbuscular mycorrhizal fungi. Mycorrhiza 2003, 13, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Gao, J.-L.; Zhu, H.-H.; Long, L.-K.; Xing, Q.-X.; Chen, J.-Z. Evaluation of the potential of trap plants to detect arbuscular mycorrhizal fungi using polymerase chain reaction-denaturing gradient gel electrophoresis analysis. Soil Sci. Plant Nutr. 2010, 56, 205–211. [Google Scholar] [CrossRef]
- Trejo-Aguilar, D.; Banuelos, J. Isolation and culture of arbuscular mycorrhizal fungi from field samples. Methods Mol. Biol. 2020, 2146, 1–18. [Google Scholar] [CrossRef]
- Oehl, F.; Sieverding, E.; Mäder, P.; Dubois, D.; Ineichen, K.; Boller, T.; Wiemken, A. Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia 2004, 138, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Vieira, L.C.; da Silva, D.K.A.; Escobar, I.E.C.; da Silva, J.M.; de Moura, I.A.; Oehl, F.; da Silva, G.A. Changes in an arbuscular mycorrhizal fungi community along an environmental gradient. Plants 2020, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Guzman, A.; Montes, M.; Hutchins, L.; DeLaCerda, G.; Yang, P.; Kakouridis, A.; Dahlquist-Willard, R.M.; Firestone, M.K.; Bowles, T.; Kremen, C. Crop diversity enriches arbuscular mycorrhizal fungal communities in an intensive agricultural landscape. New Phytol. 2021, 231, 447–459. [Google Scholar] [CrossRef]
- Pellegrino, E.; Gamper, H.A.; Ciccolini, V.; Ercoli, L. Forage rotations conserve diversity of arbuscular mycorrhizal fungi and soil fertility. Front. Microbiol. 2019, 10, 2969. [Google Scholar] [CrossRef] [PubMed]
- Stürmer, S.L.; Stürmer, R.; Pasqualini, D. Taxonomic diversity and community structure of arbuscular mycorrhizal fungi (Phylum Glomeromycota) in three maritime sand dunes in Santa Catarina state, south Brazil. Fungal Ecol. 2013, 6, 27–36. [Google Scholar] [CrossRef]
- da Silva, D.K.A.; Pereira, C.M.R.; de Souza, R.G.; da Silva, G.A.; Oehl, F.; Maia, L.C. Diversity of arbuscular mycorrhizal fungi in restinga and dunes areas in Brazilian Northeast. Biodivers. Conserv. 2012, 21, 2361–2373. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Y.; Qiu, Q.; Xin, G.; Yang, Z.; Shi, S. Flooding greatly affects the diversity of arbuscular mycorrhizal fungi communities in the roots of wetland plants. PLoS ONE 2011, 6, e24512. [Google Scholar] [CrossRef] [PubMed]
- Klinnawee, L.; Noirungsee, N.; Nopphakat, K.; Runsaeng, P.; Chantarachot, T. Flooding overshadows phosphorus availability in controlling the intensity of arbuscular mycorrhizal colonization in Sangyod Muang Phatthalung lowland indica rice. ScienceAsia 2021, 47, 202–210. [Google Scholar] [CrossRef]
- Land Development Department Key to the reference soil groups. In World Reference Base for Soil Resources of Thailand; Soil Resources Survey and Research Division, Land Development Department: Bangkok, Thailand, 2021; pp. 14–16.
- Fournier, J.; Gallon, R.K.; Paris, R. G2Sd: A new R package for the statistical analysis of unconsolidated sediments. Geomorphol. Reli. Process. Environ. 2014, 20, 73–78. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021.
- Shamrikova, E.V.; Kondratenok, B.M.; Tumanova, E.A.; Vanchikova, E.V.; Lapteva, E.M.; Zonova, T.V.; Lu-Lyan-Min, E.I.; Davydova, A.P.; Libohova, Z.; Suvannang, N. Transferability between soil organic matter measurement methods for database harmonization. Geoderma 2022, 412, 115547. [Google Scholar] [CrossRef]
- Kane, P.F. (Ed.) AOAC International AOAC Official Method 958.01. In Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 2016; p. 6. [Google Scholar]
- Kane, P.F. (Ed.) AOAC International AOAC Official Method 970.01. In Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 2016; p. 9. [Google Scholar]
- Kane, P.F. (Ed.) AOAC International AOAC Official Method 983.02. In Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 2016; pp. 23–26. [Google Scholar]
- Baird, R.; Bridgewater, L. 4500-Sulfate. In Standard Methods for the Examination of Water and Wastewater; Bridgewater, L., Ed.; American Public Health Association: Washington, DC, USA, 2017; pp. 199–200. [Google Scholar]
- Trejo-Aguilar, D.; Banuelos, J. Isolation and culture of arbuscular mycorrhizal fungi from field samples. In Arbuscular Mycorrhizal Fungi: Methods and Protocols; Ferrol, N., Lanfranco, L., Eds.; Springer: New York, NY, USA, 2020; pp. 1–18. ISBN 978-1-0716-0603-2. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Rivers, A.R.; Weber, K.C.; Gardner, T.G.; Liu, S.; Armstrong, S.D. ITSxpress: Software to rapidly trim internally transcribed spacer sequences with quality scores for marker gene analysis. F1000Research 2018, 7, 1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. UNITE QIIME Release for Fungi Version 10.05.2021. UNITE Community. Available online: https://plutof.ut.ee/#/doi/10.15156/BIO/1264708 (accessed on 24 June 2022).
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, S.; Young, J.P.W. Improved PCR primers for the detection and identification of arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 2008, 65, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal Omega, accurate alignment of very large numbers of sequences. Methods Mol. Biol. 2014, 1079, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S. Kmer: An R Package for Fast Alignment-Free Clustering of Biological Sequences; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 June 2021).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- López-García, Á. Analysis of arbuscular mycorrhizal fungal communities by terminal restriction fragment length polymorphism (TRFLP). In Arbuscular Mycorrhizal Fungi; Methods in Molecular Biology; Humana: New York, NY, USA, 2020; Volume 2146, pp. 117–136. [Google Scholar]
- Chaiyasen, A.; Young, J.P.W.; Teaumroong, N.; Gavinlertvatana, P.; Lumyong, S. Characterization of Arbuscular Mycorrhizal Fungus Communities of Aquilaria crassna and Tectona grandis Roots and Soils in Thailand Plantations. PLoS ONE 2014, 9, e112591. [Google Scholar] [CrossRef]
- Arrigo, N.; Holderegger, R.; Alvarez, N. Automated Scoring of AFLPs Using RawGeno v 2.0, a Free R CRAN Library BT. In Data Production and Analysis in Population Genomics: Methods and Protocols; Pompanon, F., Bonin, A., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 155–175. ISBN 978-1-61779-870-2. [Google Scholar]
- Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research, R Package Version 1.2-3. 2015. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 1 June 2021).
- Noppradit, P.; Pradit, S.; Muenhor, D.; Doungsuwan, N.; Whangsani, U.; Sama, N.; Towatana, P. Investigation of 37 years weather record and its relation to human health: A case study in Songkhla Province, Southern Thailand. Int. J. Agric. Technol. 2021, 17, 1507–1520. [Google Scholar]
- Orlandi Costa, H.A.; Orlandi Costa, H.A.; Stuermer, S.L.; Ragonezi, C.; Grazziotti, P.H.; Fonseca Santos Grazziotti, D.C.; Silva, E.d.B. Species richness and root colonization of arbuscular mycorrhizal fungi in Syngonanthus elegans, an endemic and threatened species from the Cerrado domain in Brazil. Cienc. E Agrotecnologia 2016, 40, 326–336. [Google Scholar] [CrossRef] [Green Version]
- Burley, S.T.; Harper, K.A.; Lundholm, J.T. Vegetation composition, structure and soil properties across coastal forest–barren ecotones. Plant Ecol. 2010, 211, 279–296. [Google Scholar] [CrossRef]
- Lammerts, E.J.; Grootjans, A.P. Nutrient deficiency in dune slack pioneer vegetation: A review. J. Coast. Conserv. 2014, 3, 87. [Google Scholar] [CrossRef] [Green Version]
- Nopphakat, K.; Runsaeng, P.; Klinnawee, L. Acaulospora as the dominant arbuscular mycorrhizal fungi in organic lowland rice paddies improves phosphorus availability in soils. Sustainability 2022, 14, 31. [Google Scholar] [CrossRef]
- Martins, S.C.; Sousa Neto, E.; de Piccolo, M.C.; Almeida, D.Q.A.; Camargo, P.B.de; do Carmo, J.B.; Porder, S.; Lins, S.R.M.; Martinelli, L.A. Soil texture and chemical characteristics along an elevation range in the coastal Atlantic Forest of Southeast Brazil. Geoderma Reg. 2015, 5, 106–116. [Google Scholar] [CrossRef]
- Turner, B.L.; Yavitt, J.B.; Harms, K.E.; Garcia, M.N.; Wright, S.J. Seasonal changes in soil organic matter after a decade of nutrient addition in a lowland tropical forest. Biogeochemistry 2015, 123, 221–235. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, D.; Liang, G.; Qiu, Q.; Liu, J.; Zhou, G.; Liu, S.; Chu, G.; Yan, J. Effects of precipitation on soil organic carbon fractions in three subtropical forests in southern China. J. Plant Ecol. 2016, 9, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Vallino, M.; Fiorilli, V.; Bonfante, P. Rice flooding negatively impacts root branching and arbuscular mycorrhizal colonization, but not fungal viability. Plant. Cell Environ. 2014, 37, 557–572. [Google Scholar] [CrossRef]
- Sivakumar, N. Effect of edaphic factors and seasonal variation on spore density and root colonization of arbuscular mycorrhizal fungi in sugarcane fields. Ann. Microbiol. 2013, 63, 151–160. [Google Scholar] [CrossRef]
- Fabián, D.; Guadarrama, P.; Hernadez-Cuevas, L.; Ramos-Zapata, J.A. Arbuscular mycorrhizal fungi in a coastal wetland in Yucatan, Mexico. Bot. Sci. 2018, 96, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Botnen, S.; Kauserud, H.; Carlsen, T.; Blaalid, R.; Høiland, K. Mycorrhizal fungal communities in coastal sand dunes and heaths investigated by pyrosequencing analyses. Mycorrhiza 2015, 25, 447–456. [Google Scholar] [CrossRef]
- Chialva, M.; Ghignone, S.; Cozzi, P.; Lazzari, B.; Bonfante, P.; Abbruscato, P.; Lumini, E. Water management and phenology influence the root-associated rice field microbiota. FEMS Microbiol. Ecol. 2020, 96, fiaa146. [Google Scholar] [CrossRef]
- Rodríguez-Echeverría, S.; Freitas, H. Diversity of AMF associated with Ammophila arenaria ssp. arundinacea in Portuguese sand dunes. Mycorrhiza 2006, 16, 543–552. [Google Scholar] [CrossRef] [Green Version]
- Bouamri, R.; Dalpé, Y.; Serrhini, M.N.; Bennani, A. Arbuscular mycorrhizal fungi species associated with rhizosphere of Phoenix dactylifera L. in Morocco. Afr. J. Biotechnol. 2006, 5, 510–516. [Google Scholar] [CrossRef]
- Selvakumar, G.; Shagol, C.C.; Kang, Y.; Chung, B.N.; Han, S.G.; Sa, T.M. Arbuscular mycorrhizal fungi spore propagation using single spore as starter inoculum and a plant host. J. Appl. Microbiol. 2018, 124, 1556–1565. [Google Scholar] [CrossRef]
- El Hilali, R.; Symanczik, S.; El kinany, S.; Oehl, F.; Ouahmane, L.; Bouamri, R. Cultivation, identification, and application of arbuscular mycorrhizal fungi associated with date palm plants in Drâa-Tafilalet oasis. Rhizosphere 2022, 22, 100521. [Google Scholar] [CrossRef]
- Han, X.; Xu, C.; Wang, Y.; Huang, D.; Fan, Q.; Xin, G.; Müller, C. Dynamics of Arbuscular Mycorrhizal Fungi in Relation to Root Colonization, Spore Density, and Soil Properties among Different Spreading Stages of the Exotic Plant Threeflower Beggarweed (Desmodium triflorum) in a Zoysia tenuifolia Lawn. Weed Sci. 2019, 67, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Eom, A.-H.; Hartnett, D.C.; Wilson, G.W.T. Host plant species effects on arbuscular mycorrhizal fungal communities in tallgrass prairie. Oecologia 2000, 122, 435–444. [Google Scholar] [CrossRef]
- Zubek, S.; Majewska, M.L.; Błaszkowski, J.; Stefanowicz, A.M.; Nobis, M.; Kapusta, P. Invasive plants affect arbuscular mycorrhizal fungi abundance and species richness as well as the performance of native plants grown in invaded soils. Biol. Fertil. Soils 2016, 52, 879–893. [Google Scholar] [CrossRef]
- Trejo-Aguilar, D.; Lara-Capistrán, L.; Maldonado-Mendoza, I.E.; Zulueta-Rodríguez, R.; Sangabriel-Conde, W.; Mancera-López, M.E.; Negrete-Yankelevich, S.; Barois, I. Loss of arbuscular mycorrhizal fungal diversity in trap cultures during long-term subculturing. IMA Fungus 2013, 4, 161–167. [Google Scholar] [CrossRef]
- Cousins, J.R.; Hope, D.; Gries, C.; Stutz, J.C. Preliminary assessment of arbuscular mycorrhizal fungal diversity and community structure in an urban ecosystem. Mycorrhiza 2003, 13, 319–326. [Google Scholar] [CrossRef]
- Covacevich, F.; Hernández Guijarro, K.; Crespo, E.M.; Lumini, E.; Rivero Mega, M.S.; Lugo, M.A. Arbuscular mycorrhizal fungi from argentinean highland puna soils unveiled by propagule multiplication. Plants 2021, 10, 1803. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter (Unit) | Dry Season | Wet Season |
|---|---|---|
| Organic matter (%w/w) | 0.8 ± 0.3 | 1.7 ± 0.4 * |
| Total N (ppm) | 418.5 ± 227.0 | 837.7 ± 271.3 * |
| NO3− (ppm) | 8.6 ± 3.9 | 10.7 ± 5.1 |
| NH4+ (ppm) | 2.9 ± 2.1 | 2.8 ± 0.8 |
| Total P (ppm) | 16.8 ± 2.8 | 25.8 ± 17.2 |
| Available P (ppm) | 13.0 ± 2.9 | 16.6 ± 12.3 |
| K (ppm) | 21.6 ± 16.4 | 9.0 ± 2.3 |
| Ca (ppm) | 57.9 ± 9.4 | 43.1 ± 8.0 * |
| Mg (ppm) | 8.9 ± 1.7 | 5.2 ± 0.4 * |
| S (ppm) | 20.9 ± 8.2 | 20.5 ± 17.1 |
| pH | 5.2 ± 0.3 | 5.5 ± 0.3 |
| EC (dS/m) | 0.010 ± 0.002 | 0.011 ± 0.001 |
| Moisture content (%w/w) | 0.7 ± 0.2 | 19.6 ± 1.2 * |
| Comparison | R2 | p-Value |
|---|---|---|
| Native soil × Maize | 0.36702 | 0.0083 ** |
| Native soil × Sorghum | 0.29207 | 0.0171 * |
| Native soil × Maize + Sorghum | 0.22444 | 0.017 * |
| Native soil × Blank | 0.3121 | 0.0089 ** |
| Maize × Sorghum | 0.12514 | 0.3605 |
| Maize × Maize + Sorghum | 0.14634 | 0.1581 |
| Maize × Blank | 0.27034 | 0.0092 ** |
| Sorghum × Maize + Sorghum | 0.09452 | 0.6704 |
| Sorghum × Blank | 0.17119 | 0.0775 |
| Maize + Sorghum × Blank | 0.15955 | 0.0677 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tenzin, U.W.; Noirungsee, N.; Runsaeng, P.; Noppradit, P.; Klinnawee, L. Dry-Season Soil and Co-Cultivated Host Plants Enhanced Propagation of Arbuscular Mycorrhizal Fungal Spores from Sand Dune Vegetation in Trap Culture. J. Fungi 2022, 8, 1061. https://doi.org/10.3390/jof8101061
Tenzin UW, Noirungsee N, Runsaeng P, Noppradit P, Klinnawee L. Dry-Season Soil and Co-Cultivated Host Plants Enhanced Propagation of Arbuscular Mycorrhizal Fungal Spores from Sand Dune Vegetation in Trap Culture. Journal of Fungi. 2022; 8(10):1061. https://doi.org/10.3390/jof8101061
Chicago/Turabian StyleTenzin, Ugyen Wangmo, Nuttapol Noirungsee, Phanthipha Runsaeng, Prakrit Noppradit, and Lompong Klinnawee. 2022. "Dry-Season Soil and Co-Cultivated Host Plants Enhanced Propagation of Arbuscular Mycorrhizal Fungal Spores from Sand Dune Vegetation in Trap Culture" Journal of Fungi 8, no. 10: 1061. https://doi.org/10.3390/jof8101061