
All-Trans Retinoic Acid Effect on Candida albicans Growth and Biofilm Formation

 , ,
, ,  , , and
, , and 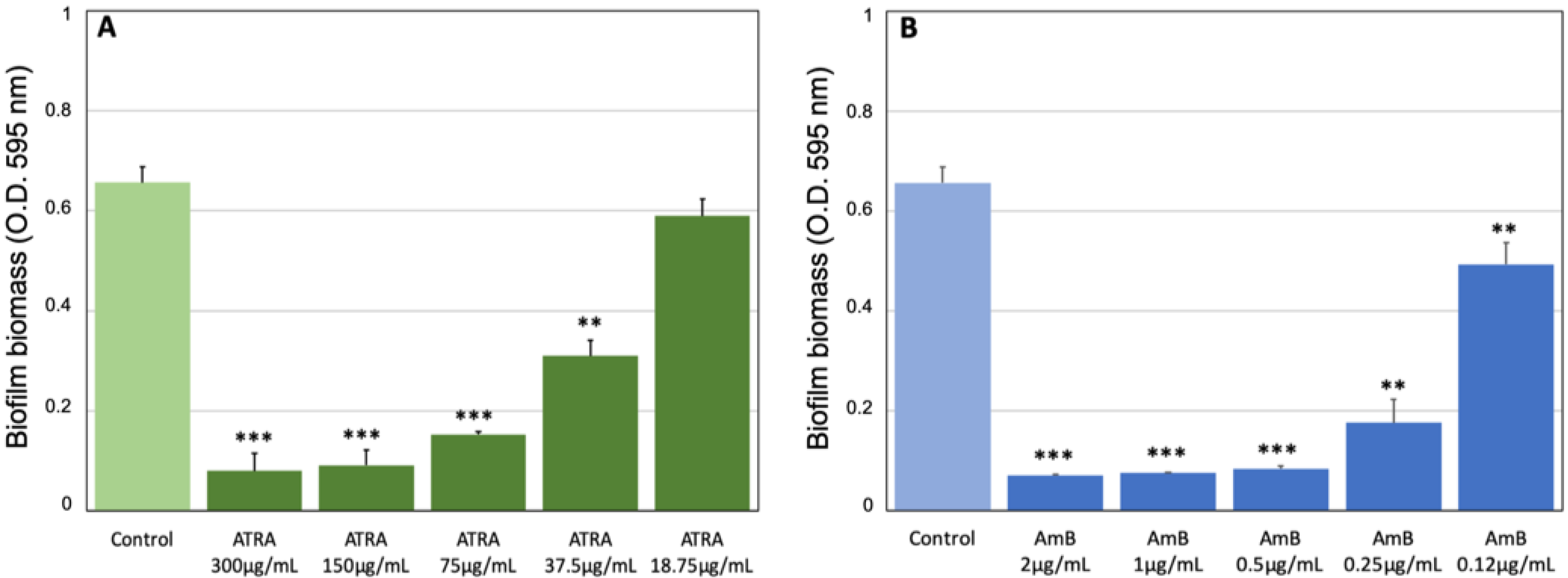{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Candida Strain and Growth Conditions
2.2. Antimicrobial Agents
2.3. Antifungal Susceptibility Testing
2.4. Biofilm Quantification by Crystal Violet and XTT Assays
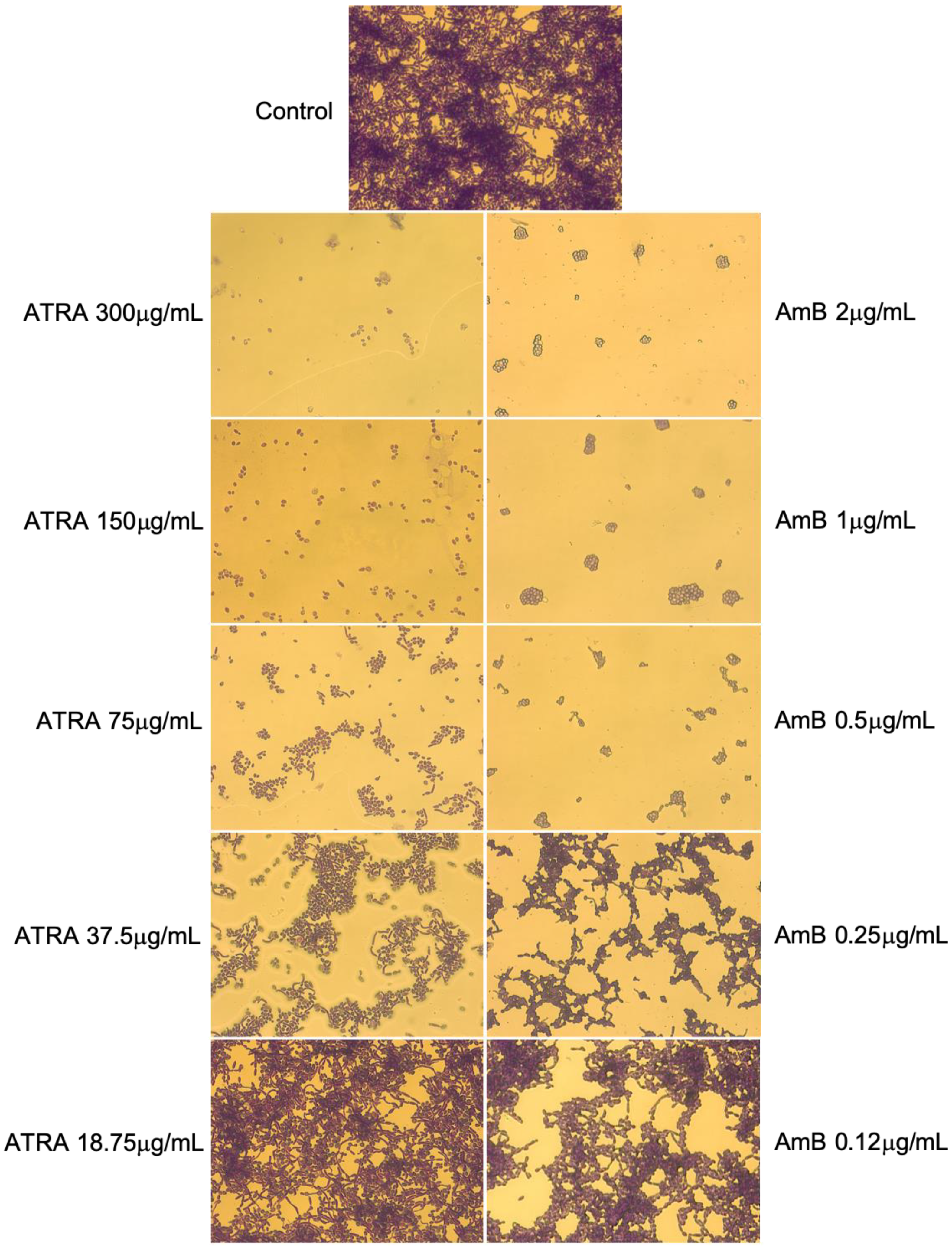
2.5. Hyphal Growth Inhibition Assay
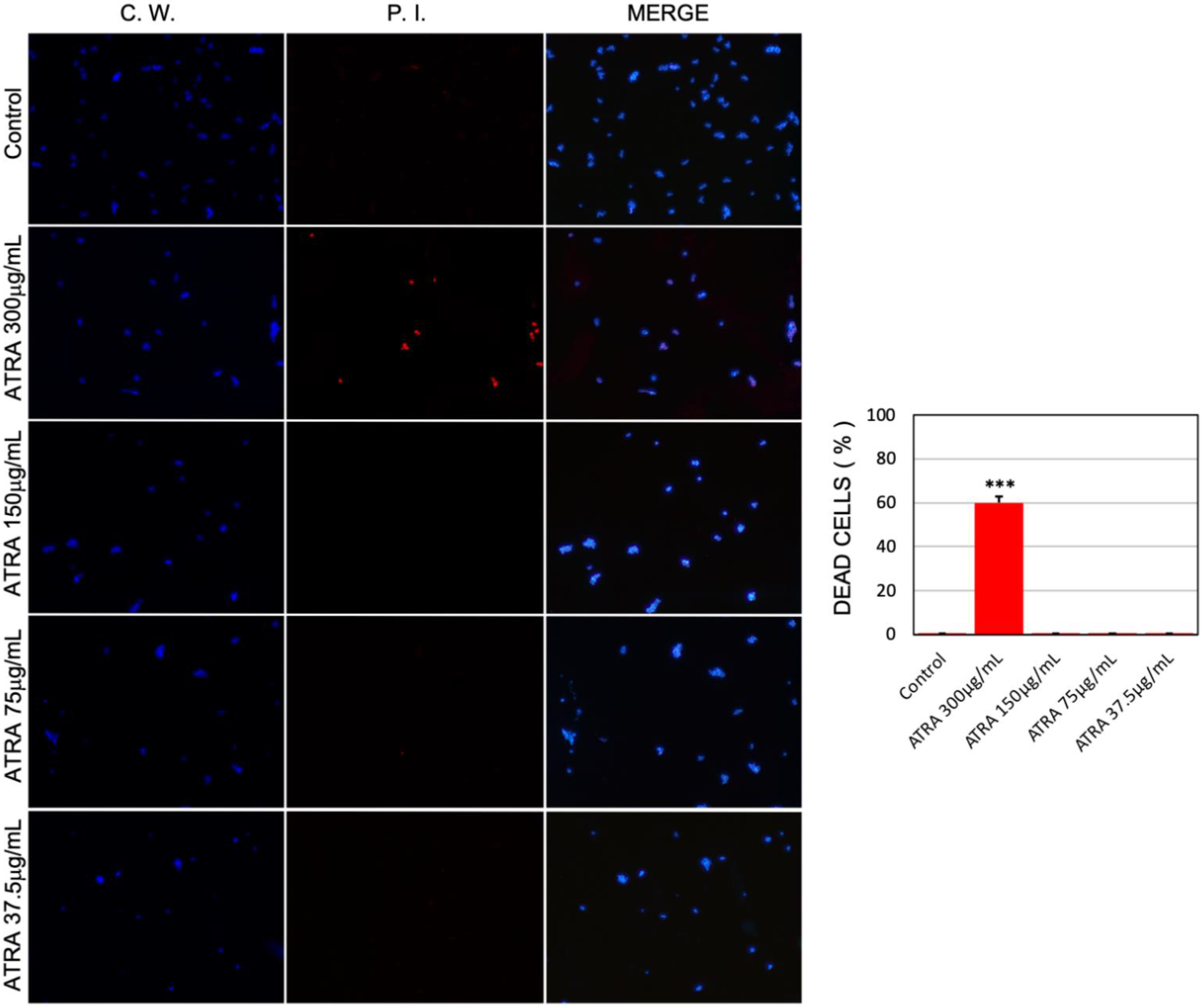
2.6. Visualization and Quantification of Vitality of Candida albicans Cells
2.7. Observations by Transmission Electron Microscopy
2.8. Statistical Analysis
3. Results
3.1. Antifungal Effect of ATRA against C. albicans Growth
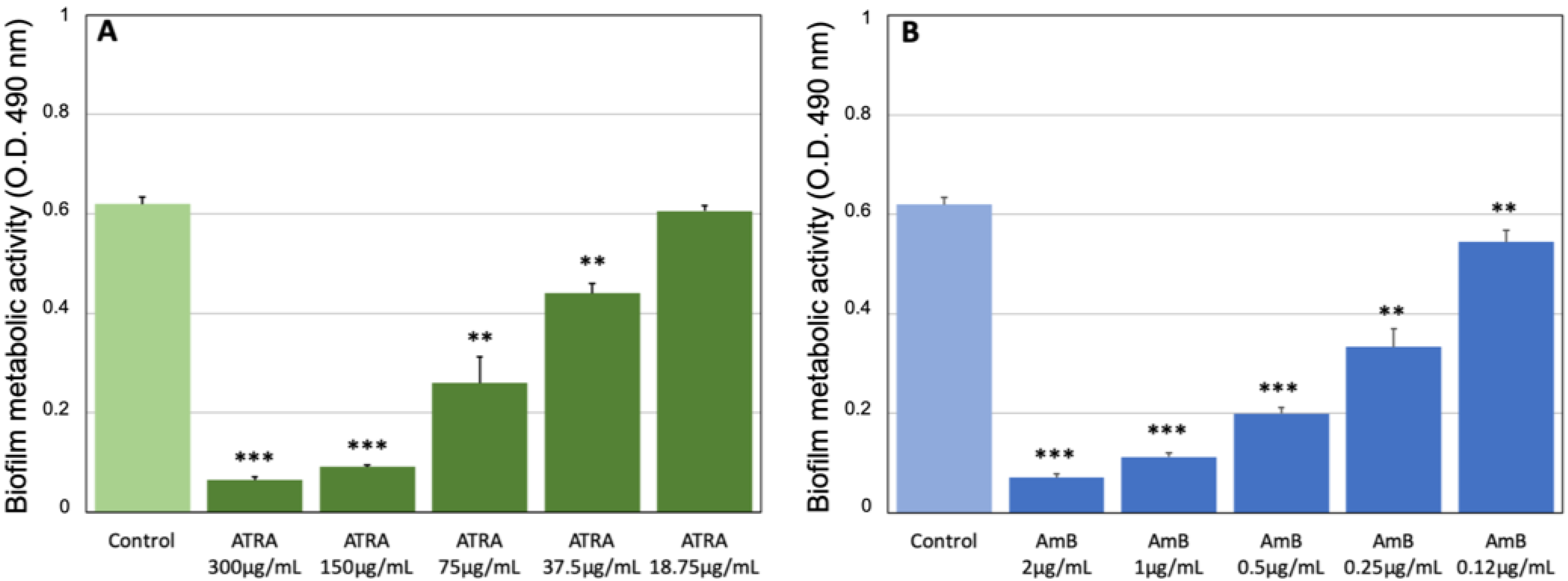
3.2. Effect of ATRA on C. albicans Biofilm Production
3.3. ATRA Exerts a Dose-Dependent Fungicidal and/or Fungistatic Activity against C. albicans
3.4. Impact of ATRA on Yeast-to-Hyphal Dimorphic Transition of C. albicans
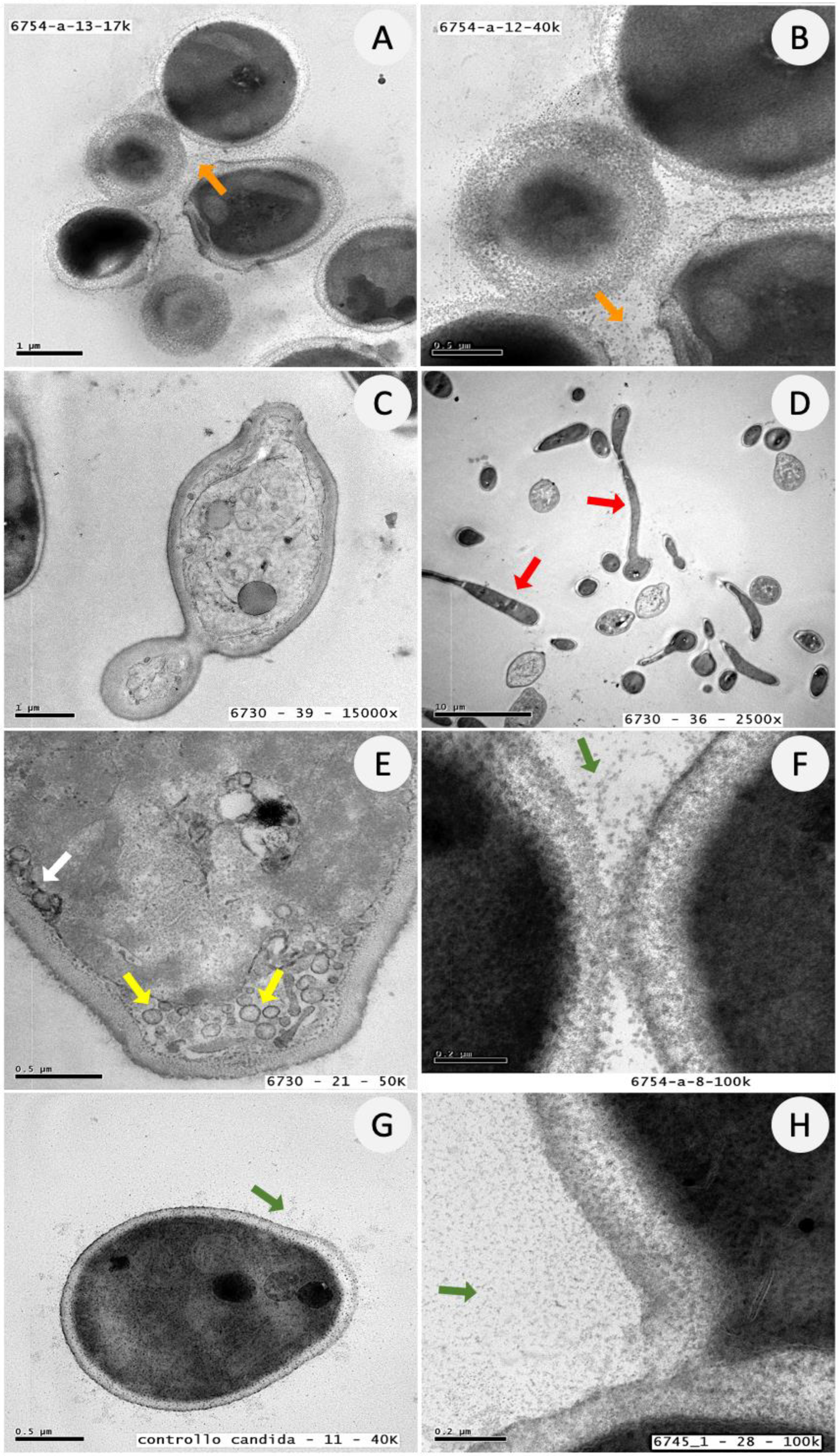
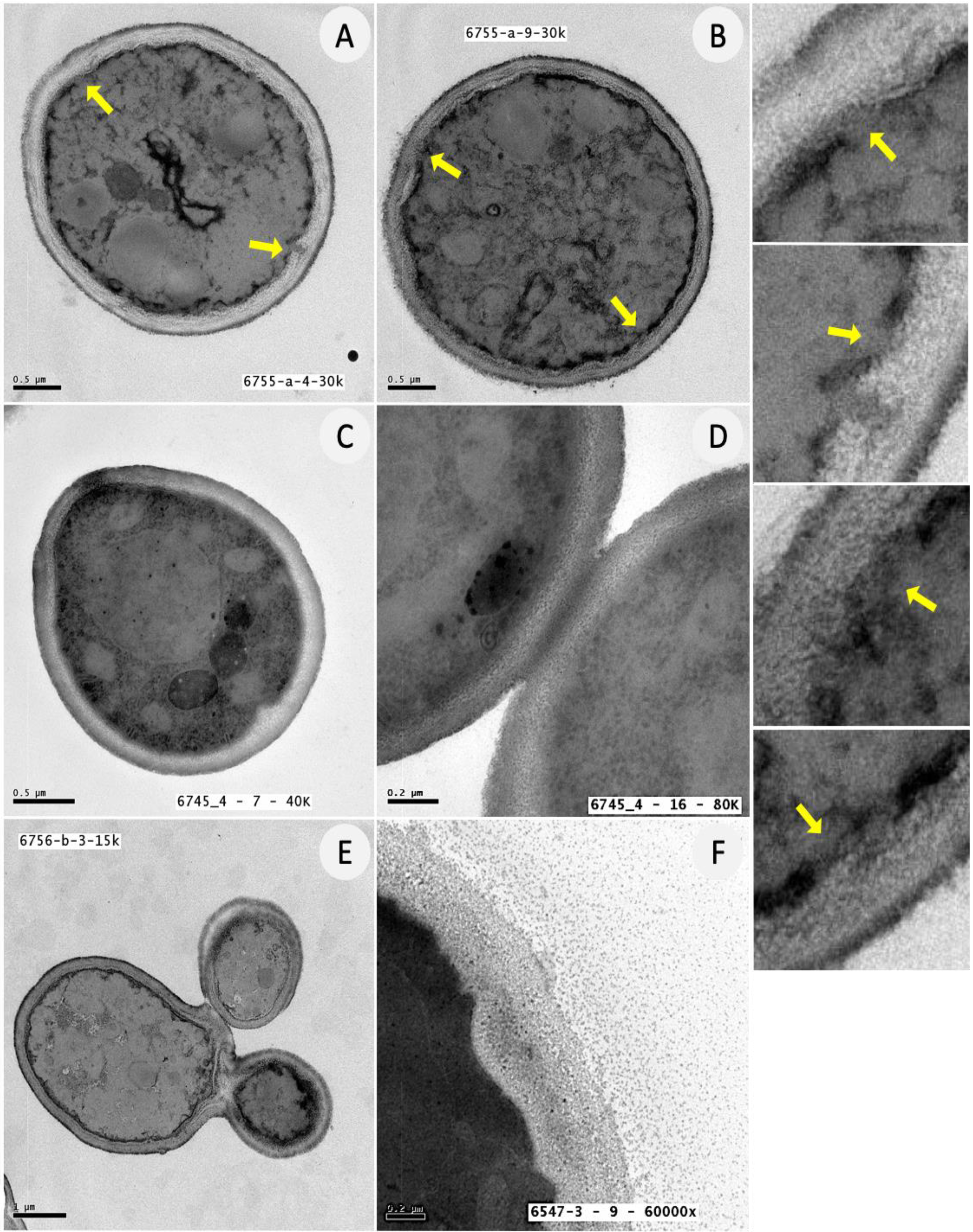
3.5. Ultrastructural Analysis of C. albicans Biofilm Cells upon Exposure to ATRA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roemer, T.; Krysan, D.J. Antifungal drug development: Challenges, unmet clinical needs, and new approaches. Cold Spring Harb. Perspect. Med. 2014, 4, a019703. [Google Scholar] [CrossRef]
- Garcia-Vidal, C.; Upton, A.; Kirby, K.A.; Marr, K.A. Epidemiology of invasive mold infections in allogeneic stem cell transplant recipients: Biological risk factors for infection according to time after transplantation. Clin. Infect. Dis. 2008, 47, 1041–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [Green Version]
- Arkowitz, R.A.; Bassilana, M. Recent advances in understanding Candida albicans hyphal growth. F1000Research 2019, 8, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nami, S.; Mohammadi, R.; Vakili, M.; Khezripour, K.; Mirzaei, H.; Morovati, H. Fungal vaccines, mechanism of actions and immunology: A comprehensive review. Biomed. Pharmacother. 2019, 109, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Spivak, E.S.; Hanson, K.E. Candida auris: An Emerging Fungal Pathogen. J. Clin. Microbiol. 2018, 56, e01588-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lone, S.A.; Ahmad, A. Candida auris-the growing menace to global health. Mycoses 2019, 62, 620–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Nosocomial bloodstream infections in US hospitals: Analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Wall, G.; Montelongo-Jauregui, D.; Vidal Bonifacio, B.; Lopez-Ribot, J.L.; Uppuluri, P. Candida albicans biofilm growth and dispersal: Contributions to pathogenesis. Curr. Opin. Microbiol. 2019, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.; Dos Santos Fontenelle, R.O.; de Brito, E.H.S.; de Morais, S.M. Biofilm of Candida albicans: Formation, regulation and resistance. J. Appl. Microbiol. 2021, 131, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, D.M.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Comparison of biofilms formed by Candida albicans and Candida parapsilosis on bioprosthetic surfaces. Infect. Immun. 2002, 70, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Ramage, G.; Wickes, B.L.; Lopez-Ribot, J.L. Biofilms of Candida albicans and their associated resistance to antifungal agents. Am. Clin. Lab. 2001, 20, 42–44. [Google Scholar] [PubMed]
- Perlroth, J.; Choi, B.; Spellberg, B. Nosocomial fungal infections: Epidemiology, diagnosis, and treatment. Med. Mycol. 2007, 45, 321–346. [Google Scholar] [CrossRef]
- Cauda, R. Candidaemia in patients with an inserted medical device. Drugs 2009, 69 (Suppl. S1), 33–38. [Google Scholar] [CrossRef]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atiencia-Carrera, M.B.; Cabezas-Mera, F.S.; Tejera, E.; Machado, A. Prevalence of biofilms in Candida spp. bloodstream infections: A meta-analysis. PLoS ONE 2022, 17, e0263522. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cerdeira, C.; Martínez-Herrera, E.; Carnero-Gregorio, M.; López-Barcenas, A.; Fabbrocini, G.; Fida, M.; El-Samahy, M.; González-Cespón, J.L. Pathogenesis and Clinical Relevance of Candida Biofilms in Vulvovaginal Candidiasis. Front. Microbiol. 2020, 11, 544480. [Google Scholar] [CrossRef]
- McKloud, E.; Delaney, C.; Sherry, L.; Kean, R.; Williams, S.; Metcalfe, R.; Thomas, R.; Richardson, R.; Gerasimidis, K.; Nile, C.J.; et al. Recurrent Vulvovaginal Candidiasis: A Dynamic Interkingdom Biofilm Disease of Candida and Lactobacillus. mSystems 2021, 6, e0062221. [Google Scholar] [CrossRef]
- Nett, J.E.; Sanchez, H.; Cain, M.T.; Andes, D.R. Genetic basis of Candida biofilm resistance due to drug-sequestering matrix glucan. J. Infect. Dis. 2010, 202, 171–175. [Google Scholar] [CrossRef] [Green Version]
- Cosio, T.; Gaziano, R.; Zuccari, G.; Costanza, G.; Grelli, S.; Di Francesco, P.; Bianchi, L.; Campione, E. Retinoids in Fungal Infections: From Bench to Bedside. Pharmaceuticals 2021, 14, 962. [Google Scholar] [CrossRef]
- Cosio, T.; Di Prete, M.; Gaziano, R.; Lanna, C.; Orlandi, A.; Di Francesco, P.; Bianchi, L.; Campione, E. Trifarotene: A Current Review and Perspectives in Dermatology. Biomedicines 2021, 9, 237. [Google Scholar] [CrossRef] [PubMed]
- Iyer, N.; Vaishnava, S. Vitamin A at the interface of host-commensal-pathogen interactions. PLoS Pathog. 2019, 15, e1007750. [Google Scholar] [CrossRef] [Green Version]
- Campione, E.; Cosio, T.; Lanna, C.; Mazzilli, S.; Ventura, A.; Dika, E.; Gaziano, R.; Dattola, A.; Candi, E.; Bianchi, L. Predictive role of vitamin A serum concentration in psoriatic patients treated with IL-17 inhibitors to prevent skin and systemic fungal infections. J. Pharmacol. Sci. 2020, 144, 52–56. [Google Scholar] [CrossRef]
- Campione, E.; Paternò, E.J.; Costanza, G.; Diluvio, L.; Carboni, I.; Marino, D.; Favalli, C.; Chimenti, S.; Bianchi, L.; Orlandi, A. Tazarotene as alternative topical treatment for onychomycosis. Drug Des. Devel. Ther. 2015, 9, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campione, E.; Gaziano, R.; Marino, D.; Orlandi, A. Fungistatic activity of all-trans retinoic acid against Aspergillus fumigatus and Candida albicans. Drug Des. Devel. Ther. 2016, 10, 1551–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campione, E.; Gaziano, R.; Doldo, E.; Marino, D.; Falconi, M.; Iacovelli, F.; Tagliaferri, D.; Pacello, L.; Bianchi, L.; Lanna, C.; et al. Antifungal Effect of All-trans Retinoic Acid against Aspergillus fumigatus In Vitro and in a Pulmonary Aspergillosis In Vivo Model. Antimicrob. Agents Chemother. 2021, 65, e01874-20. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 4th ed.; CLSI standard M27; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Sabbatini, S.; Monari, C.; Ballet, N.; Decherf, A.C.; Bozza, S.; Camilloni, B.; Perito, S.; Vecchiarelli, A. Anti-Biofilm Properties of Saccharomyces cerevisiae CNCM I-3856 and Lacticaseibacillus rhamnosus ATCC 53103 Probiotics against G. vaginalis. Microorganisms 2020, 8, 1294. [Google Scholar] [CrossRef]
- Gulati, M.; Lohse, M.B.; Ennis, C.L.; Gonzalez, R.E.; Perry, A.M.; Bapat, P.; Arevalo, A.V.; Rodriguez, D.L.; Nobile, C.J. In Vitro Culturing and Screening of Candida albicans Biofilms. Curr. Protoc. Microbiol. 2018, 50, e60. [Google Scholar] [CrossRef]
- Ramage, G.; Vande Walle, K.; Wickes, B.L.; López-Ribot, J.L. Standardized method for in vitro antifungal susceptibility testing of Candida albicans biofilms. Antimicrob. Agents Chemother. 2001, 45, 2475–2479. [Google Scholar] [CrossRef] [Green Version]
- Sangetha, S.; Zuraini, Z.; Suryani, S.; Sasidharan, S. In situ TEM and SEM studies on the antimicrobial activity and prevention of Candida albicans biofilm by Cassia spectabilis extract. Micron 2009, 40, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, B.J.; Arendrup, M.C. Invasive Candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [Green Version]
- McCarty, T.P.; Pappas, P.G. Invasive Candidiasis. Infect. Dis. Clin. North. Am. 2016, 30, 103–124. [Google Scholar] [CrossRef]
- Gonzalez-Lara, M.F.; Ostrosky-Zeichner, L. Invasive Candidiasis. Semin. Respir. Crit. Care Med. 2020, 41, 3–12. [Google Scholar] [CrossRef]
- Chan, A.K.Y.; Tsang, Y.C.; Chu, C.H.; Tsang, C.S.P. Aspirin as an Antifungal-Lock Agent in Inhibition of Candidal Biofilm Formation in Surgical Catheters. Infect. Drug Resist. 2021, 14, 1427–1433. [Google Scholar] [CrossRef]
- Schinabeck, M.K.; Long, L.A.; Hossain, M.A.; Chandra, J.; Mukherjee, P.K.; Mohamed, S.; Ghannoum, M.A. Rabbit model of Candida albicans biofilm infection: Liposomal amphotericin B antifungal lock therapy. Antimicrob. Agents Chemother. 2004, 48, 1727–1732. [Google Scholar] [CrossRef] [Green Version]
- Salinas, B.; Guembe, M.; Cussó, L.; Kestler, M.; Guinea, J.; Desco, M.; Muñoz, P.; Bouza, E. Assessment of the anti-biofilm effect of micafungin in an animal model of catheter-related candidemia. Med. Mycol. 2019, 57, 496–503. [Google Scholar] [CrossRef]
- Lobo, C.I.V.; Lopes, A.C.U.A.; Klein, M.I. Compounds with Distinct Targets Present Diverse Antimicrobial and Antibiofilm Efficacy against Candida albicans and Streptococcus mutans, and Combinations of Compounds Potentiate Their Effect. J. Fungi 2021, 7, 340. [Google Scholar] [CrossRef]
- Liu, Y.; Filler, S.G. Candida albicans Als3, a multifunctional adhesin and invasin. Eukaryot. Cell. 2011, 10, 168–173. [Google Scholar] [CrossRef]
- Mogavero, S.; Sauer, F.M.; Brunke, S.; Allert, S.; Schulz, D.; Wisgott, S.; Jablonowski, N.; Elshafee, O.; Krüger, T.; Kniemeyer, O.; et al. Candidalysin delivery to the invasion pocket is critical for host epithelial damage induced by Candida albicans. Cell. Microbiol. 2021, 23, e13378. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.P.; Moyes, D.L. Adaptive immune responses to Candida albicans infection. Virulence 2015, 6, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornby, J.M.; Jensen, E.C.; Lisec, A.D.; Tasto, J.J.; Jahnke, B.; Shoemaker, R.; Dussault, P.; Nickerson, K.W. Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl. Environ. Microbiol. 2001, 67, 2982–2992. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, J.; Rodrigues, M.L.; Janbon, G. Extracellular Vesicles in Fungi: Past, Present, and Future Perspectives. Front. Cell. Infect. Microbiol. 2020, 10, 346. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; Mukherjee, P.K. Candida Biofilms: Development, Architecture, and Resistance. Microbiol. Spectr. 2015, 3, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Robbins, N.; Uppuluri, P.; Nett, J.; Rajendran, R.; Ramage, G.; Lopez-Ribot, J.L.; Andes, D.; Cowen, L.E. Hsp90 governs dispersion and drug resistance of fungal biofilms. PLoS Pathog. 2011, 7, e1002257. [Google Scholar] [CrossRef]
- O’Meara, T.R.; Robbins, N.; Cowen, L.E. The Hsp90 Chaperone Network Modulates Candida Virulence Traits. Trends Microbiol. 2017, 25, 809–819. [Google Scholar] [CrossRef]
- Muñoz, M.J.; Jimenez, J. Genetic interactions between Hsp90 and the Cdc2 mitotic machinery in the fission yeast Schizosaccharomyces pombe. Mol. Gen. Genet. 1999, 261, 242–250. [Google Scholar] [CrossRef]
- Aligue, R.; Akhavan-Niak, H.; Russell, P. A role for Hsp90 in cell cycle control: Wee1 tyrosine kinase activity requires interaction with Hsp90. EMBO J. 1994, 13, 6099–6106. [Google Scholar] [CrossRef]
- Brown, A.J.; Leach, M.D.; Nicholls, S. The relevance of heat shock regulation in fungal pathogens of humans. Virulence. 2010, 1, 330–332. [Google Scholar] [CrossRef]
- Gong, Y.; Li, T.; Yu, C.; Sun, S. Candida albicans Heat Shock Proteins and Hsps-Associated Signaling Pathways as Potential Antifungal Targets. Front. Cell. Infect. Microbiol. 2017, 7, 520. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; An, M.; Shen, H.; Huang, X.; Yao, X.; Liu, J.; Zhu, F.; Zhang, S.; Chen, S.; He, L.; et al. The non-Geldanamycin Hsp90 inhibitors enhanced the antifungal activity of fluconazole. Am. J. Transl. Res. 2015, 7, 2589–2602. [Google Scholar] [PubMed]
- Hall, J.A.; Grainger, J.R.; Spencer, S.P.; Belkaid, Y. The role of retinoic acid in tolerance and immunity. Immunity 2011, 35, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klassert, T.E.; Hanisch, A.; Bräuer, J.; Klaile, E.; Heyl, K.A.; Mansour, M.K.; Tam, J.M.; Vyas, J.M.; Slevogt, H. Modulatory role of vitamin A on the Candida albicans-induced immune response in human monocytes. Med. Microbiol. Immunol. 2014, 203, 425. [Google Scholar] [CrossRef] [Green Version]
- Girmenia, C.; Lo Coco, F.; Breccia, M.; Latagliata, R.; Spadea, A.; D’Andrea, M.; Gentile, G.; Micozzi, A.; Alimena, G.; Martino, P.; et al. Infectious complications in patients with acute promyelocytic leukaemia treated with the AIDA regimen. Leukemia 2003, 17, 925–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arce, F.; Gätjens-Boniche, O.; Vargas, E.; Valverde, B.; Dìaz, C. Apoptiotic events induced by naturally occurring retinoids ATRA and 13-cis retinoic acid on human hepatoma cell lines Hep3B and HepG2. Cancer Lett. 2005, 229, 271–281. [Google Scholar] [CrossRef]
- Freemantle, S.J.; Spinella, M.J.; Dmitrovsky, E. Retinoids in cancer therapy and chemoprevention: Promise meets resistance. Oncogene 2003, 22, 7305–7315. [Google Scholar] [CrossRef] [Green Version]
- Agadir, A.; Cornic, M.; Lefebvre, P.; Gourmel, B.; Jérôme, M.; Degos, L.; Fenaux, P.; Chomienne, C. All-trans retinoic acid pharmacokinetics and bioavailability in acute promyelocytic leukemia: Intracellular concentrations and biologic response relationship. J. Clin. Oncol. 1995, 13, 2517–2523. [Google Scholar] [CrossRef]
- Jing, J.; Nelson, C.; Paik, J.; Shirasaka, Y.; Amory, J.K.; Isoherranen, N. Physiologically Based Pharmacokinetic Model of All-trans-Retinoic Acid with Application to Cancer Populations and Drug Interactions. J. Pharmacol. Exp. Ther. 2017, 361, 246–258. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pistoia, E.S.; Cosio, T.; Campione, E.; Pica, F.; Volpe, A.; Marino, D.; Di Francesco, P.; Monari, C.; Fontana, C.; Favaro, M.; et al. All-Trans Retinoic Acid Effect on Candida albicans Growth and Biofilm Formation. J. Fungi 2022, 8, 1049. https://doi.org/10.3390/jof8101049
Pistoia ES, Cosio T, Campione E, Pica F, Volpe A, Marino D, Di Francesco P, Monari C, Fontana C, Favaro M, et al. All-Trans Retinoic Acid Effect on Candida albicans Growth and Biofilm Formation. Journal of Fungi. 2022; 8(10):1049. https://doi.org/10.3390/jof8101049
Chicago/Turabian StylePistoia, Enrico Salvatore, Terenzio Cosio, Elena Campione, Francesca Pica, Antonio Volpe, Daniele Marino, Paolo Di Francesco, Claudia Monari, Carla Fontana, Marco Favaro, and et al. 2022. "All-Trans Retinoic Acid Effect on Candida albicans Growth and Biofilm Formation" Journal of Fungi 8, no. 10: 1049. https://doi.org/10.3390/jof8101049