
Saccharomyces cerevisiae Cells Lacking the Zinc Vacuolar Transporter Zrt3 Display Improved Ethanol Productivity in Lignocellulosic Hydrolysates

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Media and Growth Conditions
2.3. Cell Viability Assays
2.4. Assessment of Reactive Oxygen Species (ROS) Accumulation in Response to Acetic Acid Treatment by Flow Cytometry
2.5. Assessment of Catalase and Superoxide Dismutase Activity
2.5.1. Preparation of Protein Extracts
2.5.2. Assessment of Mitochondrial Superoxide Dismutase Activity by Native Gel Electrophoresis
2.5.3. Assessment of Catalase Activity
2.6. Assessment of Pep4 Localization and Vacuolar Membrane Permeabilization by Fluorescence Microscopy
2.7. Evaluation of Vacuolar pH by Flow Cytometry
2.8. Zinc Supplementation and Limitation Assays
2.9. Assessment of zrt3Δ Cell Growth and Fermentative Performance in the Presence of Acetic Acid
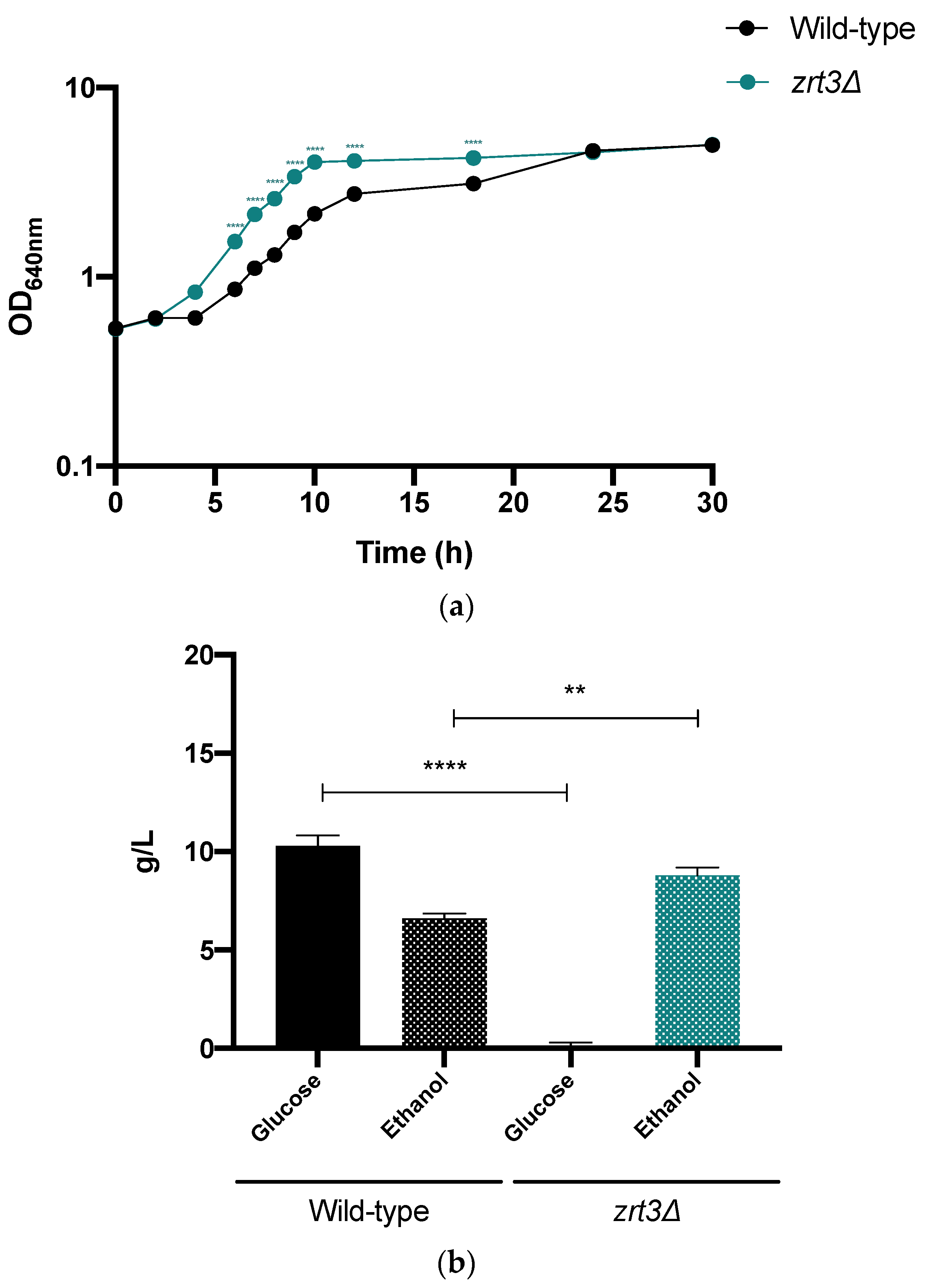
2.10. Assessment of zrt3Δ Cell Growth and Fermentative Performance in a Lignocellulosic Hydrolysate
3. Results
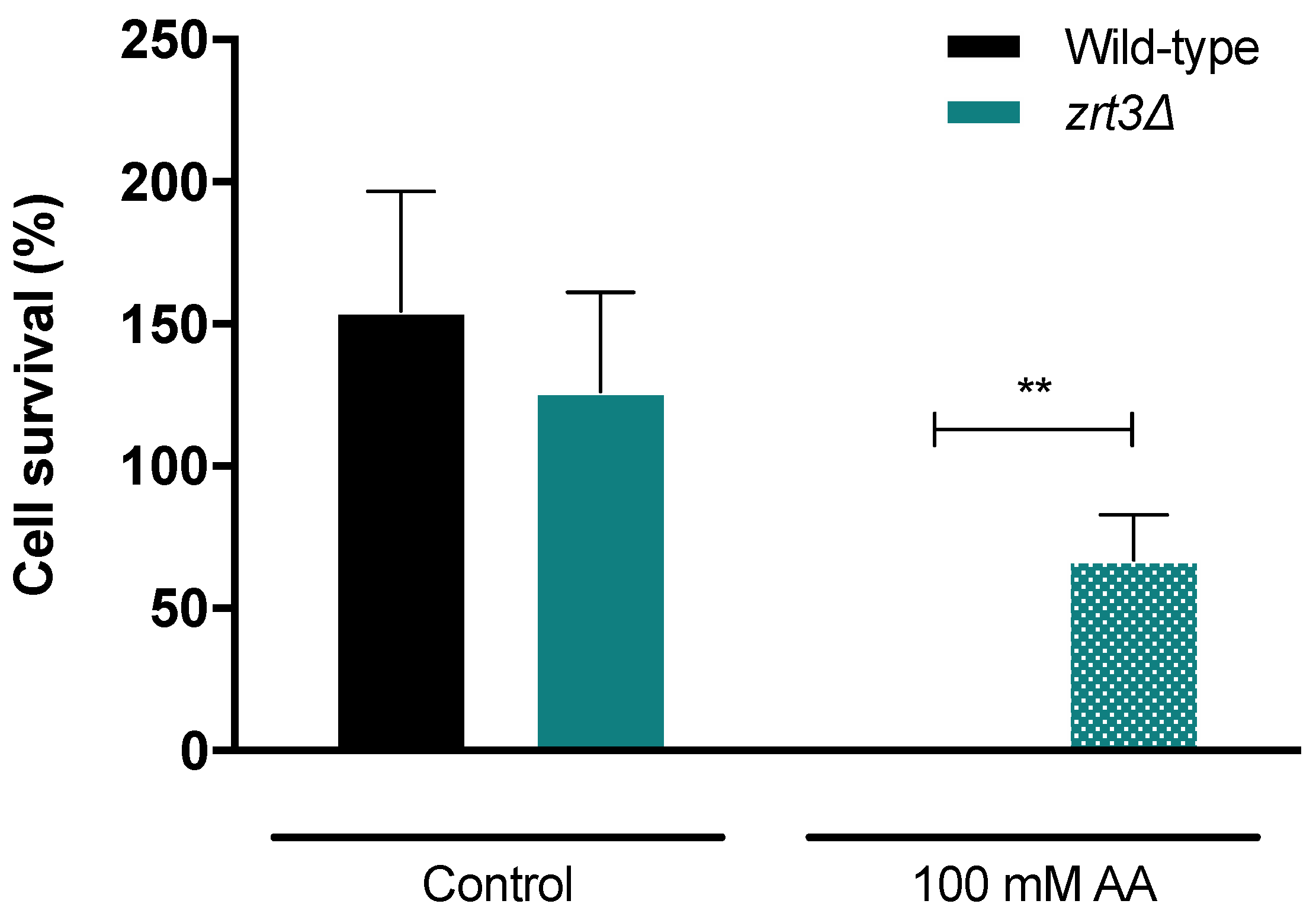
3.1. The zrt3Δ Strain Is Resistant to Acetic Acid-Induced Cell Death
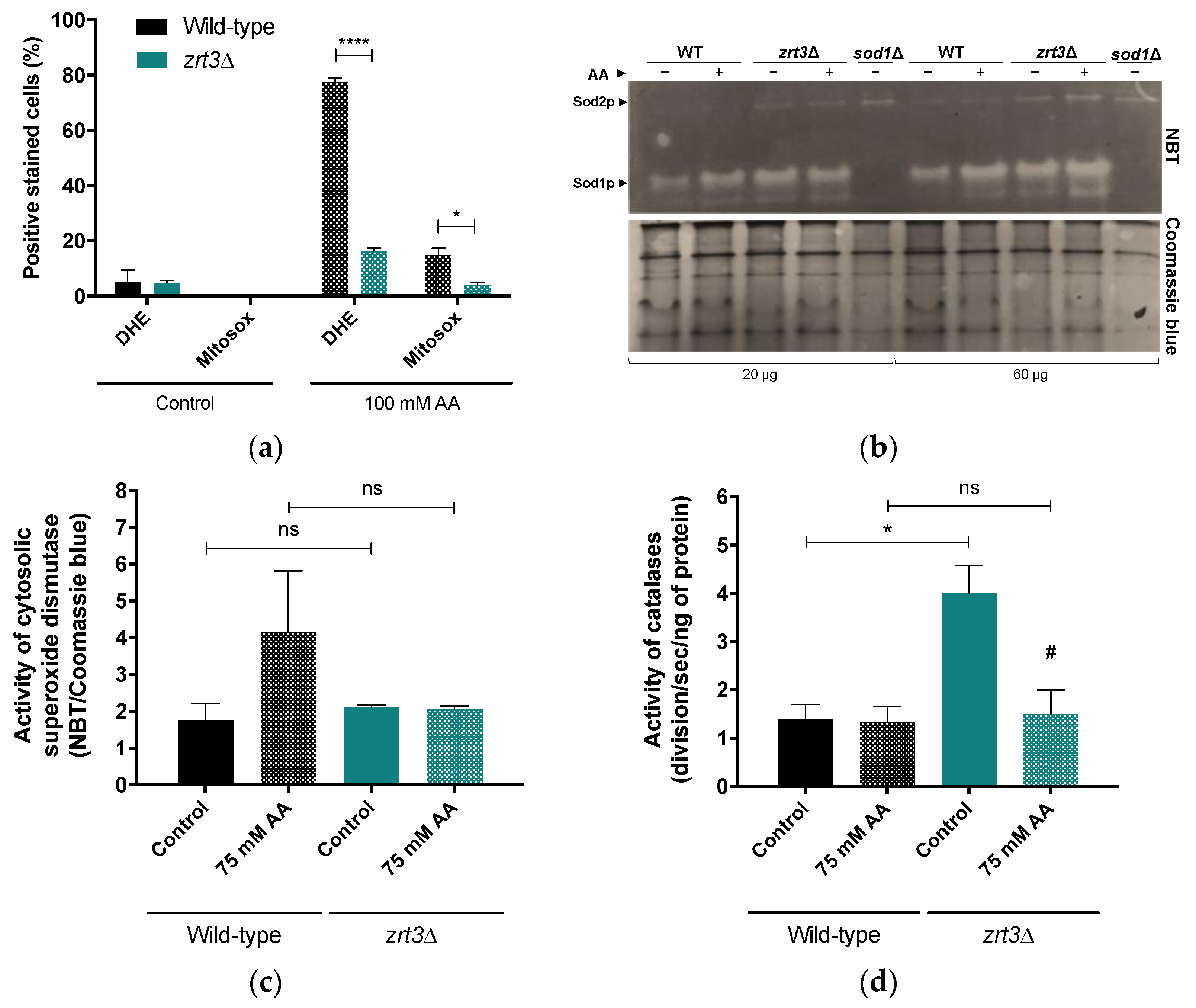
3.2. The Acetic Acid Resistance Phenotype of zrt3Δ Is Associated with Lower Accumulation of Superoxide Anion and Increased Activity of Catalases
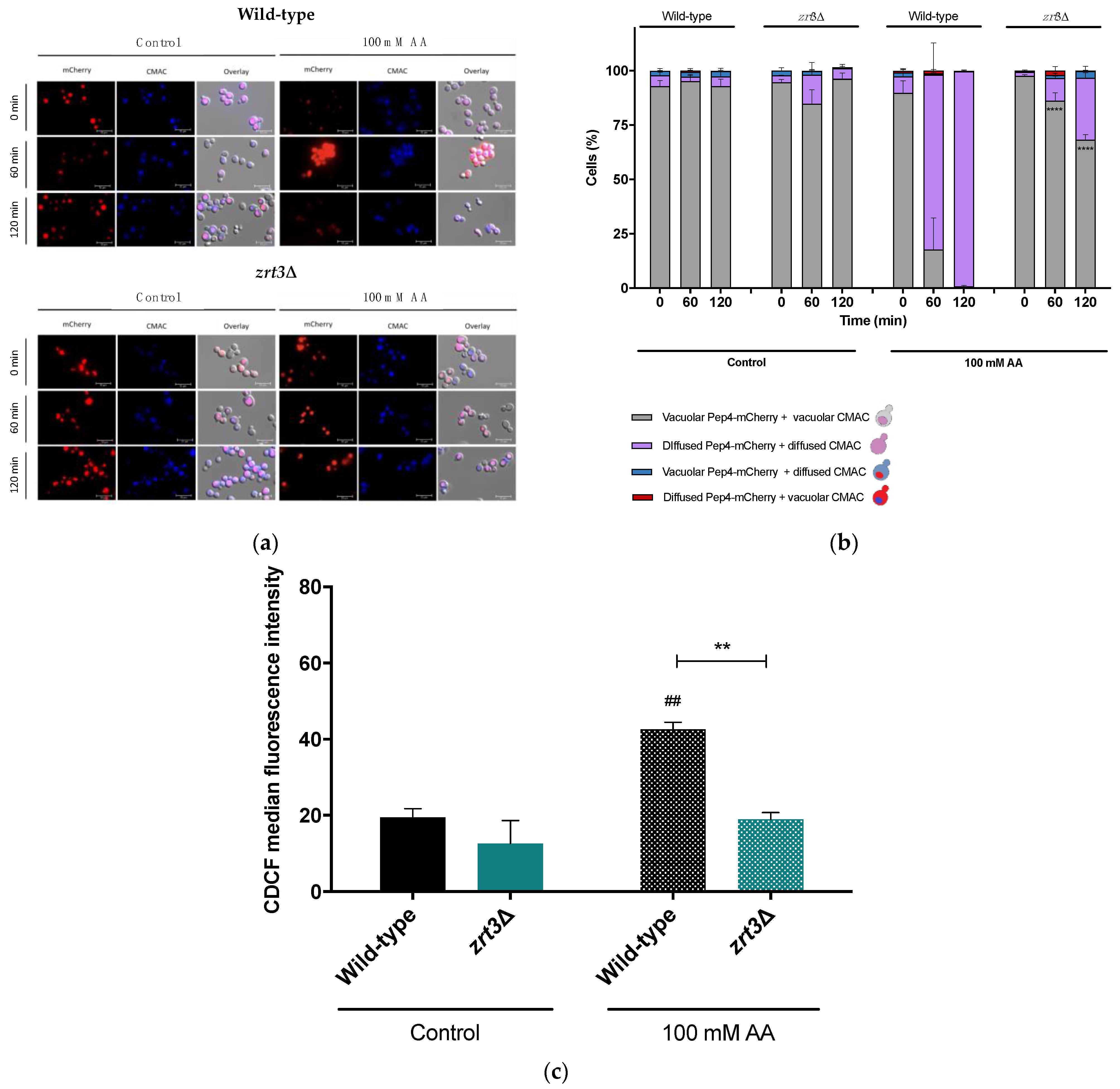
3.3. The zrt3Δ Acetic Acid Resistance Phenotype Is Associated with Delayed Vacuolar Alterations and Maintenance of Vacuolar pH
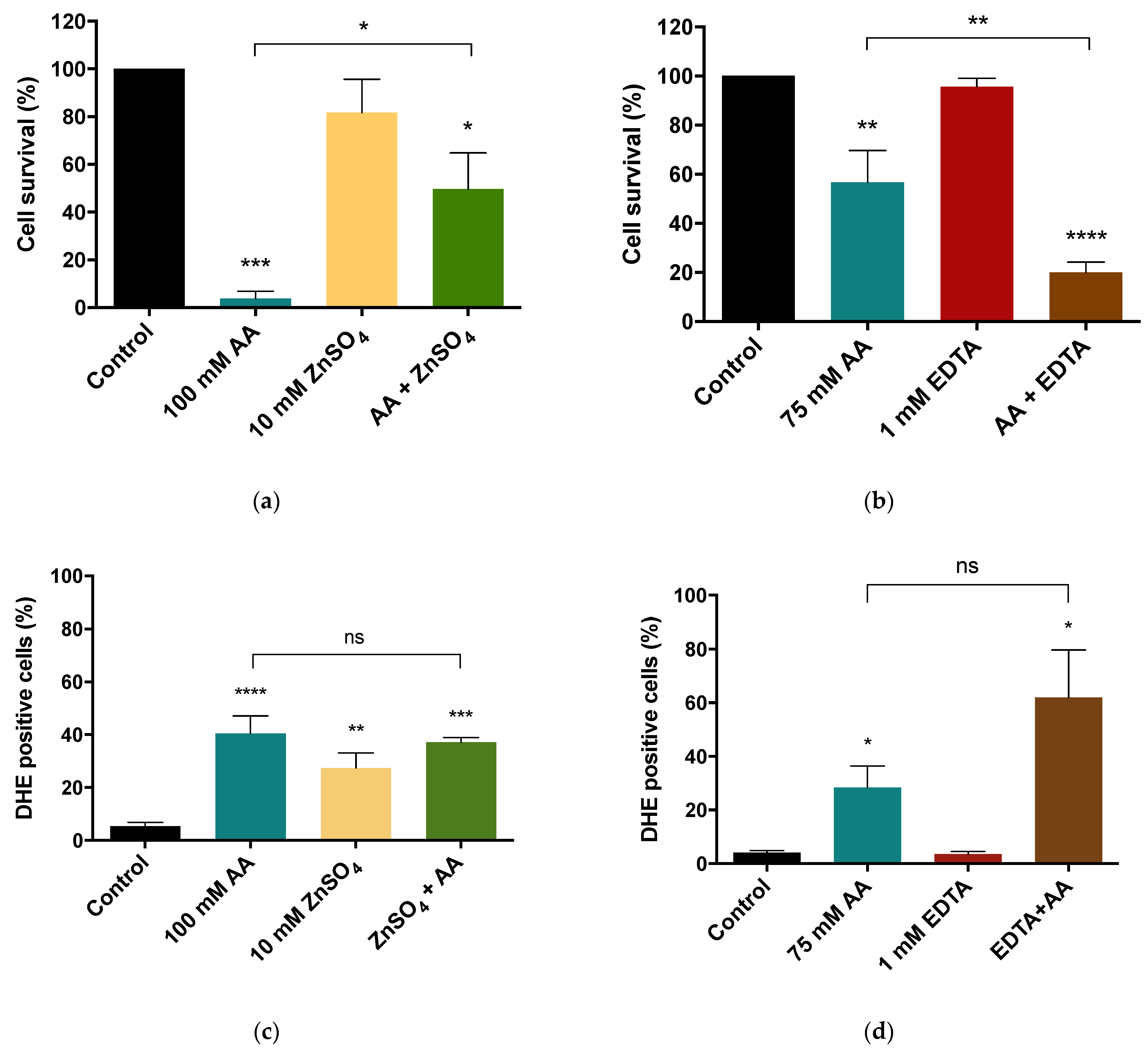
3.4. Loss of Cell Survival and Superoxide Anion Accumulation Induced by Acetic Acid Are Affected by Zinc Availability
3.5. zrt3Δ Displays Improved Fermentative Performance in the Presence of Acetic Acid
3.6. zrt3Δ Shows Improved Fermentative Performance in a Lignocellulosic Hydrolysate
4. Discussion
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| AA | Acetic acid |
| CAT | Catalase |
| CDCFDA | 5-(and -6)-carboxy-2′, 7′-dichlorofluorescein diacetate |
| c.f.u. | Colony Forming Units |
| CMAC | 7-amino-4-chloromethylcoumarin |
| DIC | Differential Interference Contrast |
| DHE | Dihydroethidium |
| DMSO | Dimethyl sulfoxide |
| F | Furfural |
| FA | Formic acid |
| GHG | Greenhouse gases |
| HMF | Hydroxymethylfurfural |
| LE | Lignocellulosic ethanol |
| LH | Lignocellulosic hydrolysates |
| NBT | Nitro Blue Tetrazolium |
| RCD | Regulated cell death |
| ROS | Reactive oxygen species |
| S. cerevisiae | Saccharomyces cerevisiae |
| Schef. stipitis | Scheffersomyces stipitis |
| SOD | Superoxide dismutase |
| VMP | Vacuolar membrane permeabilization |
| Vpr | Vine pruning residue |
| YEPD | Yeast Extract Peptone Dextrose |
References
- Lin, Y.; Tanaka, S. Ethanol fermentation from biomass resources: Current state and prospects. Appl. Microbiol. Biotechnol. 2006, 69, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Sharma, P.; Saran, A.K.; Singh, N.; Bishnoi, N.R. Comparative study on ethanol production from pretreated sugarcane bagasse using immobilized Saccharomyces cerevisiae on various matrices. Renew. Energy 2013, 50, 488–493. [Google Scholar] [CrossRef]
- Tursi, A. A review on biomass: Importance, chemistry, classification, and conversion. Biofuel Res. J. 2019, 6, 962–979. [Google Scholar] [CrossRef]
- Cunha, J.T.; Soares, P.O.; Baptista, S.L.; Costa, C.E.; Domingues, L. Engineered Saccharomyces cerevisiae for lignocellulosic valorization: A review and perspectives on bioethanol production. Bioengineered 2020, 11, 883–903. [Google Scholar] [CrossRef]
- Jönsson, L.J.; Martín, C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour. Technol. 2016, 199, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Cunha, J.T.; Romaní, A.; Costa, C.E.; Sá-Correia, I.; Domingues, L. Molecular and physiological basis of Saccharomyces cerevisiae tolerance to adverse lignocellulose-based process conditions. Appl. Microbiol. Biotechnol. 2019, 103, 159–175. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Gutierrez, D.; Sommer, C.; Andryushkova, A.; Kroemer, G.; Madeo, F. A higher spirit: Avoiding yeast suicide during alcoholic fermentation. Cell Death Differ. 2012, 19, 913–914. [Google Scholar] [CrossRef]
- Zhang, G.C.; Kong, I.I.; Wei, N.; Peng, D.; Turner, T.L.; Sung, B.H.; Sohn, J.H.; Jin, Y.S. Optimization of an acetate reduction pathway for producing cellulosic ethanol by engineered yeast. Biotechnol. Bioeng. 2016, 113, 2587–2596. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Xu, Z.; Jönsson, L.J. Profiling of Saccharomyces cerevisiae transcription factors for engineering the resistance of yeast to lignocellulose-derived inhibitors in biomass conversion. Microb. Cell Fact. 2017, 16, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.K.; Jo, J.H.; Jin, Y.S.; Seo, J.H. Enhanced ethanol fermentation by engineered Saccharomyces cerevisiae strains with high spermidine contents. Bioprocess Biosyst. Eng. 2017, 40, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Skerker, J.M.; Kong, I.I.; Kim, H.; Maurer, M.J.; Zhang, G.C.; Peng, D.; Wei, N.; Arkin, A.P.; Jin, Y.S. Metabolic engineering of a haploid strain derived from a triploid industrial yeast for producing cellulosic ethanol. Metab. Eng. 2017, 40, 176–185. [Google Scholar] [CrossRef]
- Jayakody, L.N.; Turner, T.L.; Yun, E.J.; Kong, I.I.; Liu, J.J.; Jin, Y.S. Expression of Gre2p improves tolerance of engineered xylose-fermenting Saccharomyces cerevisiae to glycolaldehyde under xylose metabolism. Appl. Microbiol. Biotechnol. 2018, 102, 8121–8133. [Google Scholar] [CrossRef] [PubMed]
- Unrean, P.; Gätgens, J.; Klein, B.; Noack, S.; Champreda, V. Elucidating cellular mechanisms of Saccharomyces cerevisiae tolerant to combined lignocellulosic-derived inhibitors using high-Throughput phenotyping and multiomics analyses. FEMS Yeast Res. 2018, 18, foy106. [Google Scholar] [CrossRef] [PubMed]
- Cunha, J.T.; Costa, C.E.; Ferraz, L.; Romaní, A.; Johansson, B.; Sá-Correia, I.; Domingues, L. HAA1 and PRS3 overexpression boosts yeast tolerance towards acetic acid improving xylose or glucose consumption: Unravelling the underlying mechanisms. Appl. Microbiol. Biotechnol. 2018, 102, 4589–4600. [Google Scholar] [CrossRef] [PubMed]
- Tran Nguyen Hoang, P.; Ko, J.K.; Gong, G.; Um, Y.; Lee, S.M. Genomic and phenotypic characterization of a refactored xylose-utilizing Saccharomyces cerevisiae strain for lignocellulosic biofuel production. Biotechnol. Biofuels 2018, 11, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.K.; Enkh-Amgalan, T.; Gong, G.; Um, Y.; Lee, S.M. Improved bioconversion of lignocellulosic biomass by Saccharomyces cerevisiae engineered for tolerance to acetic acid. GCB Bioenergy 2020, 12, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. I: Inhibition and detoxification. Bioresour. Technol. 2000, 74, 17–24. [Google Scholar] [CrossRef]
- Prudêncio, C.; Sansonetty, F.; Côrte-Real, M. Flow cytometric assessment of cell structural and functional changes induced by acetic acid in the yeasts Zygosaccharomyces bailii and Saccharomyces cerevisiae. Cytometry 1998, 31, 307–313. [Google Scholar] [CrossRef]
- Leão, C.; van Uden, N. Transport of lactate and other short-chain monocarboxylates in the yeast Candida utilis. Appl. Microbiol. Biotechnol. 1986, 23, 389–393. [Google Scholar] [CrossRef]
- Palma, M.; Guerreiro, J.F.; Sá-Correia, I. Adaptive Response and Tolerance to Acetic Acid in Saccharomyces cerevisiae and Zygosaccharomyces bailii: A Physiological Genomics Perspective. Front. Microbiol. 2018, 9, 274. [Google Scholar] [CrossRef]
- Sousa, M.J.; Ludovico, P.; Rodrigues, F.; Leo, C.; Côrte-Real, M. Stress and Cell Death in Yeast Induced by Acetic Acid. In Cell Metabolism—Cell Homeostasis and Stress Response; Bubulya, P., Ed.; IntechOpen: London, UK, 2012. [Google Scholar]
- Ludovico, P.; Sousa, M.J.; Silva, M.T.; Leão, C.; Côrte-Real, M. Saccharomyces cerevisiae commits to a programmed cell death process in response to acetic acid. Microbiology 2001, 147, 2409–2415. [Google Scholar] [CrossRef] [Green Version]
- Pinto, I.; Cardoso, H.; Leão, C.; van Uden, N. High enthalpy and low enthalpy death in Saccharomyces cerevisiae induced by acetic acid. Biotechnol. Bioeng. 1989, 33, 1350–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, S.R.; Rego, A.; Martins, V.M.; Santos-Pereira, C.; Sousa, M.J.; Côrte-Real, M. Regulation of Cell Death Induced by Acetic Acid in Yeasts. Front. Cell Dev. Biol. 2021, 9, 642375. [Google Scholar] [CrossRef]
- Pereira, C.; Chaves, S.; Alves, S.; Salin, B.; Camougrand, N.; Manon, S.; Sousa, M.J.; Côrte-Real, M. Mitochondrial degradation in acetic acid-induced yeast apoptosis: The role of Pep4 and the ADP/ATP carrier. Mol. Microbiol. 2010, 76, 1398–1410. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.; Duarte, A.M.; Fernandes, T.R.; Chaves, S.R.; Pacheco, A.; Leão, C.; Côrte-Real, M.; Sousa, M.J. Genome-wide identification of genes involved in the positive and negative regulation of acetic acid-induced programmed cell death in Saccharomyces cerevisiae. BMC Genom. 2013, 14, 838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannattasio, S.; Guaragnella, N.; Côrte-Real, M.; Passarella, S.; Marra, E. Acid stress adaptation protects Saccharomyces cerevisiae from acetic acid-induced programmed cell death. Gene 2005, 354, 93–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, C.; Zhang, M.; Fang, Q.; Xiong, L.; Zhao, X.; Hasunuma, T.; Bai, F.; Kondo, A. The impact of zinc sulfate addition on the dynamic metabolic profiling of Saccharomyces cerevisiae subjected to long term acetic acid stress treatment and identification of key metabolites involved in the antioxidant effect of zinc. Metallomics 2015, 7, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Zinc: An antioxidant and anti-inflammatory agent: Role of zinc in degenerative disorders of aging. J. Trace Elem. Med. Biol. 2014, 28, 364–371. [Google Scholar] [CrossRef]
- Xue, C.; Zhao, X.-Q.; Yuan, W.-J.; Bai, F.-W. Improving ethanol tolerance of a self-flocculating yeast by optimization of medium composition. AGRIS 2013, 24, 2257–2261. [Google Scholar] [CrossRef]
- Ismail, K.S.K.; Sakamoto, T.; Hasunuma, T.; Zhao, X.-Q.; Kondo, A. Zinc, magnesium, and calcium ion supplementation confers tolerance to acetic acid stress in industrial Saccharomyces cerevisiae utilizing xylose. Biotechnol. J. 2014, 9, 1519–1525. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-Q.; Zhang, M.-M.; Xing, Q.; Ye, P.-L.; Hasunuma, T.; Kondo, A.; Zhao, X.-Q. Improved ethanol fermentation by promoter replacement of zinc responsive genes IPL1, PRP6 and RTC1 in Saccharomyces cerevisiae. Biochem. Eng. J. 2021, 178, 108274. [Google Scholar] [CrossRef]
- Devirgiliis, C.; Murgia, C.; Danscher, G.; Perozzi, G. Exchangeable zinc ions transiently accumulate in a vesicular compartment in the yeast Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2004, 323, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Simm, C.; Lahner, B.; Salt, D.; LeFurgey, A.; Ingram, P.; Yandell, B.; Eide, D.J. Saccharomyces cerevisiae vacuole in zinc storage and intracellular zinc distribution. Eukaryot. Cell 2007, 6, 1166–1177. [Google Scholar] [CrossRef] [Green Version]
- MacDiarmid, C.W.; Milanick, M.A.; Eide, D.J. Biochemical properties of vacuolar zinc transport systems of Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 39187–39194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDiarmid, C.W.; Gaither, L.A.; Eide, D. Zinc transporters that regulate vacuolar zinc storage in Saccharomyces cerevisiae. EMBO J. 2000, 19, 2845–2855. [Google Scholar] [CrossRef] [Green Version]
- Dikalov, S.I.; Harrison, D.G. Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxid. Redox Signal. 2014, 20, 372–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Weydert, C.J.; Cullen, J.J. Measurement of superoxide dismutase, catalase and glutathione peroxidase in cultured cells and tissue. Nat. Protoc. 2010, 5, 51–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefan, C.J.; Blumer, K.J. A syntaxin homolog encoded by VAM3 mediates down-regulation of a yeast G protein-coupled receptor. J. Biol. Chem. 1999, 274, 1835–1841. [Google Scholar] [CrossRef] [Green Version]
- Roberts, C.J.; Raymond, C.K.; Yamashiro, C.T.; Stevens, T.H. Methods for studying the yeast vacuole. Guid. Yeast Genet. Mol. Biol. 1991, 194, 644–661. [Google Scholar]
- Fernandes, A.R.; Durão, P.J.; Santos, P.M.; Sá-Correia, I. Activation and significance of vacuolar H+-ATPase in Saccharomyces cerevisiae adaptation and resistance to the herbicide 2,4-dichlorophenoxyacetic acid. Biochem. Biophys. Res. Commun. 2003, 312, 1317–1324. [Google Scholar] [CrossRef]
- Teixeira, M.C.; Raposo, L.R.; Mira, N.P.; Lourenço, A.B.; Sá-Correia, I. Genome-wide identification of Saccharomyces cerevisiae genes required for maximal tolerance to ethanol. Appl. Environ. Microbiol. 2009, 75, 5761–5772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, R.L.; Cygnarowicz, M.L. A mathematical approach toward defining and calculating the duration of the lag phase. Food Microbiol. 1990, 7, 237–240. [Google Scholar] [CrossRef]
- Alves-Araújo, C.; Pacheco, A.; Almeida, M.J.; Spencer-Martins, I.; Leão, C.; Sousa, M.J. Sugar utilization patterns and respiro-fermentative metabolism in the baker’s yeast Torulaspora Delbrueckii. Microbiol. 2007, 153, 898–904. [Google Scholar] [CrossRef] [Green Version]
- Kasavi, C.; Finore, I.; Lama, L.; Nicolaus, B.; Oliver, S.G.; Toksoy Oner, E.; Kirdar, B. Evaluation of industrial Saccharomyces cerevisiae strains for ethanol production from biomass. Biomass Bioenergy 2012, 45, 230–238. [Google Scholar] [CrossRef]
- Carvalho, P.; Costa, C.E.; Baptista, S.L.; Domingues, L. Yeast cell factories for sustainable whey-to-ethanol valorisation towards a circular economy. Biofuel Res. J. 2021, 8, 1529–1549. [Google Scholar] [CrossRef]
- Maitra, S.; Narang, A. Quantifying the parametric sensitivity of ethanol production by Scheffersomyces (Pichia) stipitis: Development and verification of a method based on the principles of growth on mixtures of complementary substrates. Microbiology 2018, 164, 1348–1360. [Google Scholar] [CrossRef]
- Nakanishi, S.C.; Soares, L.B.; Biazi, L.E.; Nascimento, V.M.; Costa, A.C.; Rocha, G.J.M.; Ienczak, J.L. Fermentation strategy for second generation ethanol production from sugarcane bagasse hydrolyzate by Spathaspora passalidarum and Scheffersomyces stipitis. Biotechnol. Bioeng. 2017, 114, 2211–2221. [Google Scholar] [CrossRef]
- Zhang, W.; Geng, A. Improved ethanol production by a xylose-fermenting recombinant yeast strain constructed through a modified genome shuffling method. Biotechnol. Biofuels 2012, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cunha, J.T.; Soares, P.O.; Romaní, A.; Thevelein, J.M.; Domingues, L. Xylose fermentation efficiency of industrial Saccharomyces cerevisiae yeast with separate or combined xylose reductase/xylitol dehydrogenase and xylose isomerase pathways. Biotechnol. Biofuels 2019, 12, 1–14. [Google Scholar] [CrossRef]
- Stolp, Z.D.; Kulkarni, M.; Liu, Y.; Zhu, C.; Jalisi, A.; Lin, S.; Casadevall, A.; Cunningham, K.W.; Pineda, F.J.; Teng, X.; et al. Gene-dependent yeast cell death pathway requires AP-3 vesicle trafficking leading to vacuole membrane permeabilization. bioRxiv 2021. [Google Scholar] [CrossRef]
- Guaragnella, N.; Antonacci, L.; Giannattasio, S.; Marra, E.; Passarella, S. Catalase T and Cu, Zn-superoxide dismutase in the acetic acid-induced programmed cell death in Saccharomyces cerevisiae. FEBS Lett. 2008, 582, 210–214. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, D.J. Oxidative stress responses of the yeast Saccharomyces cerevisiae. Yeast 1998, 14, 1511–1527. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Gospodaryov, D.V. Catalases protect cellular proteins from oxidative modification in Saccharomyces cerevisiae. Cell Biol. Int. 2005, 29, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Morano, K.A.; Grant, C.M.; Moye-Rowley, W.S. The response to heat shock and oxidative stress in Saccharomyces cerevisiae. Genetics 2012, 190, 1157–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guaragnella, N.; Antonacci, L.; Passarella, S.; Marra, E.; Giannattasio, S. Achievements and perspectives in yeast acetic acid-induced programmed cell death pathways. Biochem. Soc. Trans. 2011, 39, 1538–1543. [Google Scholar] [CrossRef]
- Dell’Orco, M.; Milani, P.; Arrigoni, L.; Pansarasa, O.; Sardone, V.; Maffioli, E.; Polveraccio, F.; Bordoni, M.; Diamanti, L.; Ceroni, M.; et al. Hydrogen peroxide-mediated induction of SOD1 gene transcription is independent from Nrf2 in a cellular model of neurodegeneration. Biochim. Biophys. Acta Gene Regul. Mech. 2016, 1859, 315–323. [Google Scholar] [CrossRef]
- Martins, D.; English, A.M. Catalase activity is stimulated by H2O2 in rich culture medium and is required for H2O2 resistance and adaptation in yeast. Redox Biol. 2014, 2, 308–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devin, A.; Dejean, L.; Beauvoit, B.; Chevtzoff, C.; Avéret, N.; Bunoust, O.; Rigoulet, M. Growth yield homeostasis in respiring yeast is due to a strict mitochondrial content adjustment. J. Biol. Chem. 2006, 281, 26779–26784. [Google Scholar] [CrossRef] [Green Version]
- Eide, D.J. The oxidative stress of zinc deficiency. Metallomics 2011, 3, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Vanmarcke, G.; Demeke, M.M.; Foulquié-Moreno, M.R.; Thevelein, J.M. Identification of the major fermentation inhibitors of recombinant 2G yeasts in diverse lignocellulose hydrolysates. Biotechnol. Biofuels 2021, 14, 92. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R.M.; Modig, T.; Petersson, A.; Hähn-Hägerdal, B.; Lidén, G.; Gorwa-Grauslund, M.F. Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates by Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 2007, 82, 340–349. [Google Scholar] [CrossRef]
- Pereira, F.B.; Guimarães, P.M.R.; Gomes, D.G.; Mira, N.P.; Teixeira, M.C.; Sá-Correia, I.; Domingues, L. Identification of candidate genes for yeast engineering to improve bioethanol production in very high gravity and lignocellulosic biomass industrial fermentations. Biotechnol. Biofuels 2011, 4, 57. [Google Scholar] [CrossRef] [Green Version]
- Rosen, M.A. Environmental sustainability tools in the biofuel industry. Biofuel Res. J. 2018, 5, 751–752. [Google Scholar] [CrossRef] [Green Version]
- Aghbashlo, M.; Khounani, Z.; Hosseinzadeh-Bandbafha, H.; Gupta, V.K.; Amiri, H.; Lam, S.S.; Morosuk, T.; Tabatabaei, M. Exergoenvironmental analysis of bioenergy systems: A comprehensive review. Renew. Sustain. Energy Rev. 2021, 149, 111399. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
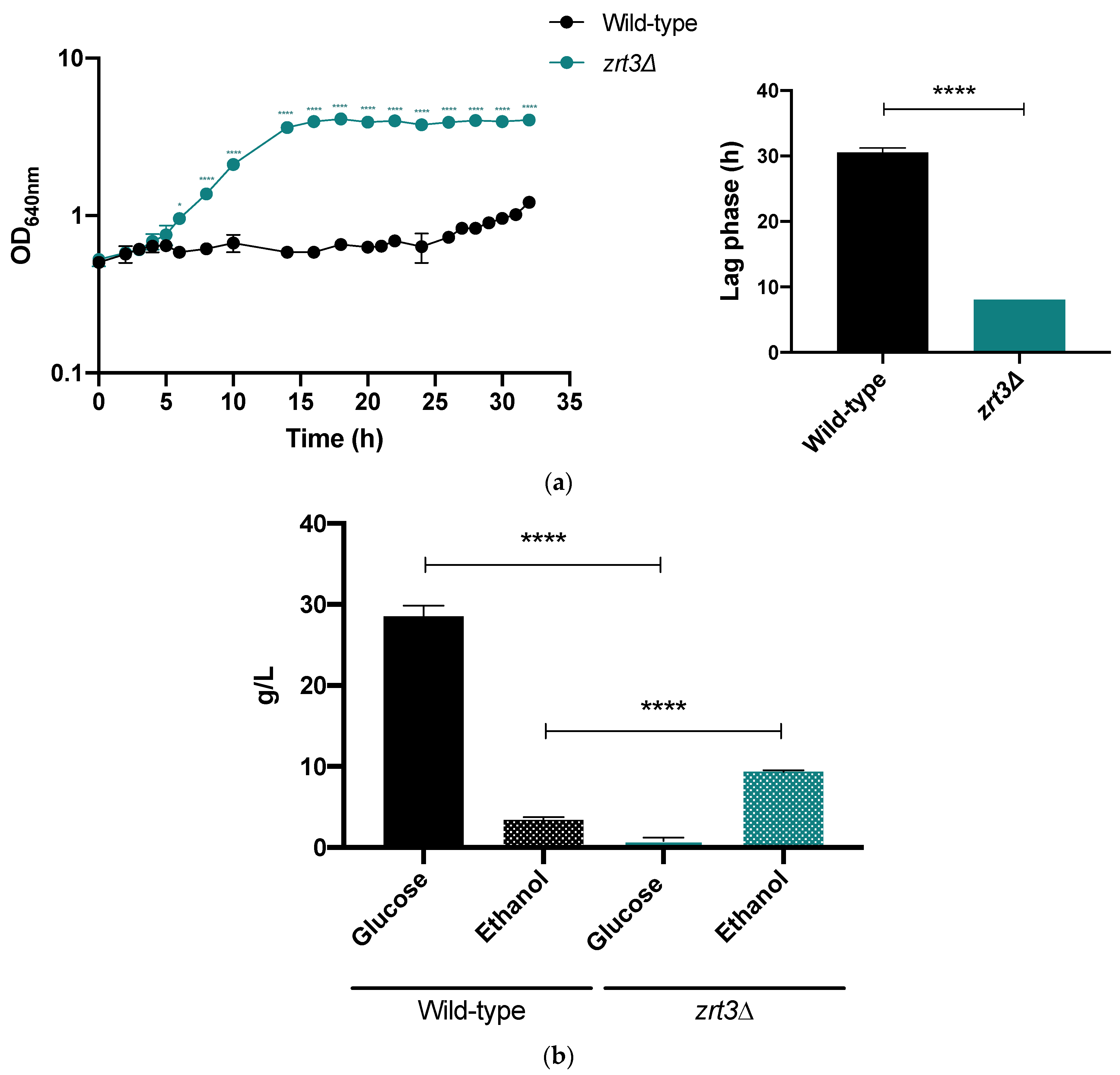{kind=link}
{kind=link}
| S. cerevisiae Strain | Modifications | Lignocellulosic Hydrolysate Composition (g/L) | Outcomes | References |
|---|---|---|---|---|
| D452-2 (MATα, leu2, his3, ura3, and can1) | Expression of the adhE and ACS gene from E. coli and a Salmonella mutant, respectively; xylose consumption: multiple copies of XYL1, XYL2, and XYL3 (genes constituting the xylose-assimilating pathway), and deletion of PHO13 and ALD6 | Miscanthus hydrolysate (20 glucose, 50 xylose, 10 acetic acid, 1 HMF, and 2 furfural) | Acetic acid consumption and faster xylose consumption; higher ethanol production and lower glycerol and xylitol production | [8] |
| BY4741 (MATa, his3Δ1, leu2Δ0, met15Δ0, ura3Δ0) | Overexpression of YAP1, STB5, WAR1, PDR8, CAT8, PUT3, and GZF3, separately | Sugarcane bagasse hydrolysate (0.89 furfural, 0.11 HMF,1.4 acetic acid, 0.03 formic acid and 0.05 levulinic acid); spruce hydrolysate (0.36 furfural, 0.03 HMF, 0.72 acetic acid, 0.27 formic acid, and 0.12 levulinic acid) | Increased relative growth rates | [9] |
| D452-2 (MATα, leu2, his3, ura3, and can1) | Overexpression of SPE3 and deletion of TPO1 and OAZ1 | Corn stover hydrolysate (3.3 acetic acid, 0.8 HMF, and 0.4 furfural) | Improved ethanol productivity | [10] |
| Haploid derivative of ATCC 4124 strain | Expression of the adhE gene from E. coli; xylose consumption: expression of XYL1, XYL2, and XYL3 from Scheffersomyces stipitis and deletion of PHO13 and ALD6 | Miscanthus hydrolysate (20 glucose, 55 xylose, 10 acetic acid, 1 HMF, and 2 furfural) | Higher ethanol productivity and lower by-product yield | [11] |
| D452-2 (MATα, leu2, his3, ura3, and can1) | Overexpression of GRE2; xylose consumption: multiple copies of XYL1, XYL2, and XYL3, and knockout ALD6 | Miscanthus hydrolysate (20 glucose, 50 xylose, 10 acetic acid, 1 HMF, and 2 furfural) | Greater robustness towards toxic hydrolysate; increased xylose consumption rate, inhibitor tolerance and ethanol production | [12] |
| BY4742 (MATa, his3Δ1, leu2Δ0, lys2Δ0, ura3Δ0) | Overexpression of TRX1 | Diluted bagasse hydrolysate (44 glucose, 5.8 xylose, 4.1 acetic acid, 0.6 furfural, and 0.2 HMF) | Larger ethanol production in titter, yield, and productivity; increased levels of protectant metabolites (trehalose, fatty acids, GABA and putrescine) | [13] |
| PE-2 (NCYC 3233) | Overexpression of HAA1 and/or PRS3; Xylose consumption: expression of XYL1 and XYL2 from Schef. stipitis, overexpression of XKS and TAL1, and deletion of GRE3 | Paulownia hydrolysate (5.84 acetic acid, 1.96 furfural, and 0.72 HMF) | Improved yeast adaptation to non-detoxified hydrolysate with high acetic acid content; increased yeast cell wall robustness under acetic acid stress situations | [14] |
| XUSAE57 | Overexpression of TAL1 and XKS1 and deletion of GRE3 and PHO13 | Sugarcane bagasse hydrolysate(34 glucose, 32 xylose, 3.1 acetic acid, 0.7 phenolics) | Improved xylose utilization and higher ethanol yield | [15,16] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terra-Matos, J.; Teixeira, M.O.; Santos-Pereira, C.; Noronha, H.; Domingues, L.; Sieiro, C.; Gerós, H.; Chaves, S.R.; Sousa, M.J.; Côrte-Real, M. Saccharomyces cerevisiae Cells Lacking the Zinc Vacuolar Transporter Zrt3 Display Improved Ethanol Productivity in Lignocellulosic Hydrolysates. J. Fungi 2022, 8, 78. https://doi.org/10.3390/jof8010078
Terra-Matos J, Teixeira MO, Santos-Pereira C, Noronha H, Domingues L, Sieiro C, Gerós H, Chaves SR, Sousa MJ, Côrte-Real M. Saccharomyces cerevisiae Cells Lacking the Zinc Vacuolar Transporter Zrt3 Display Improved Ethanol Productivity in Lignocellulosic Hydrolysates. Journal of Fungi. 2022; 8(1):78. https://doi.org/10.3390/jof8010078
Chicago/Turabian StyleTerra-Matos, Joana, Marta Oliveira Teixeira, Cátia Santos-Pereira, Henrique Noronha, Lucília Domingues, Carmen Sieiro, Hernâni Gerós, Susana Rodrigues Chaves, Maria João Sousa, and Manuela Côrte-Real. 2022. "Saccharomyces cerevisiae Cells Lacking the Zinc Vacuolar Transporter Zrt3 Display Improved Ethanol Productivity in Lignocellulosic Hydrolysates" Journal of Fungi 8, no. 1: 78. https://doi.org/10.3390/jof8010078