
Recent Advances in Sorbicillinoids from Fungi and Their Bioactivities (Covering 2016–2021)


 and
and 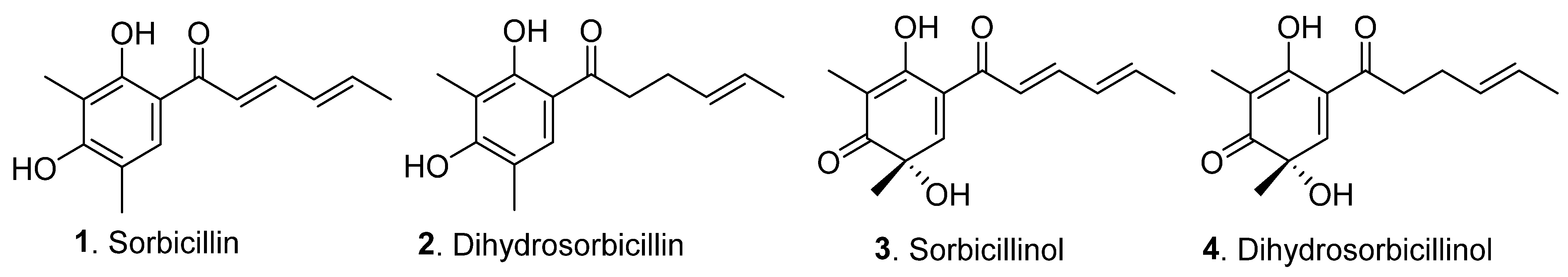{kind=link}
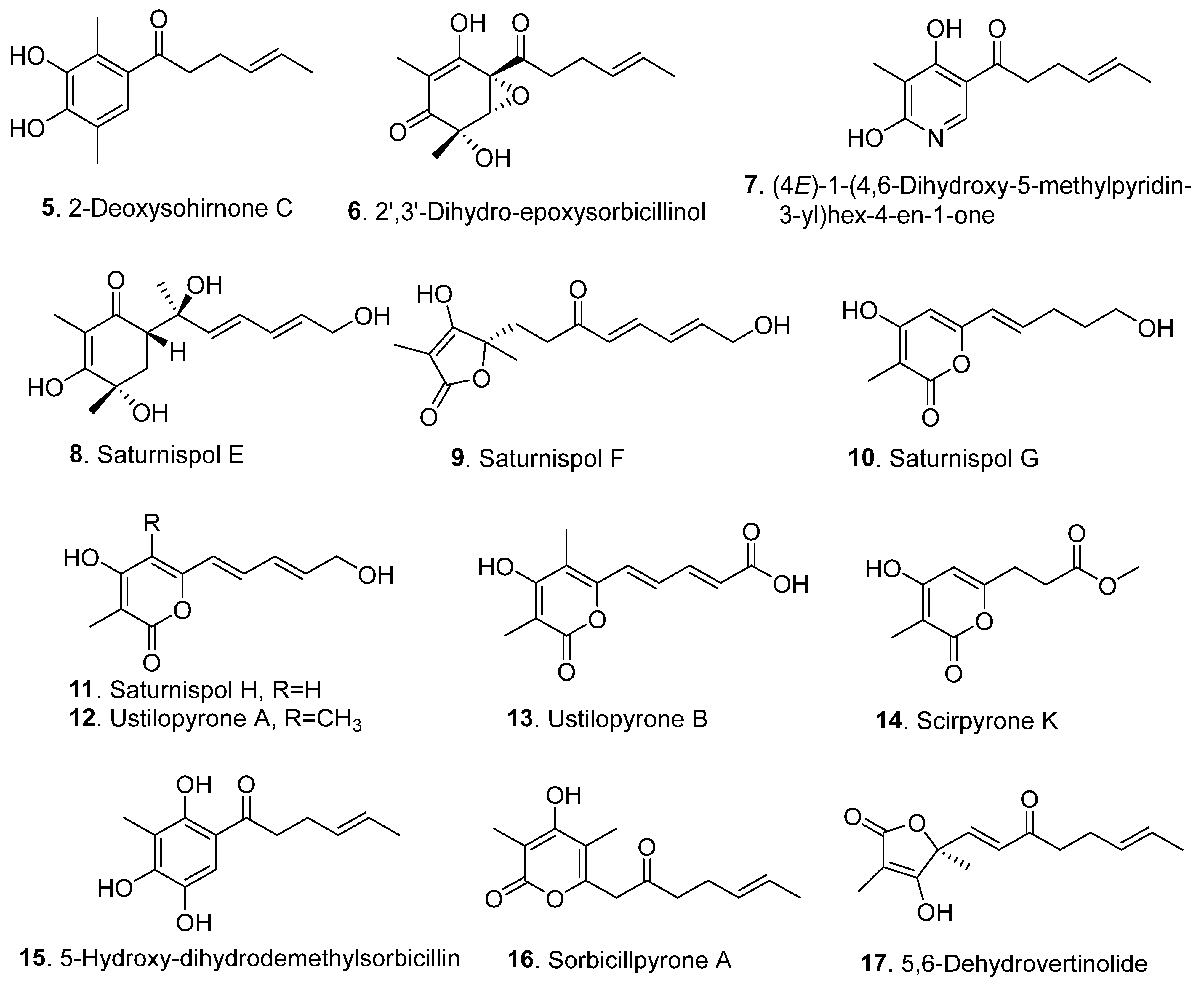{kind=link}
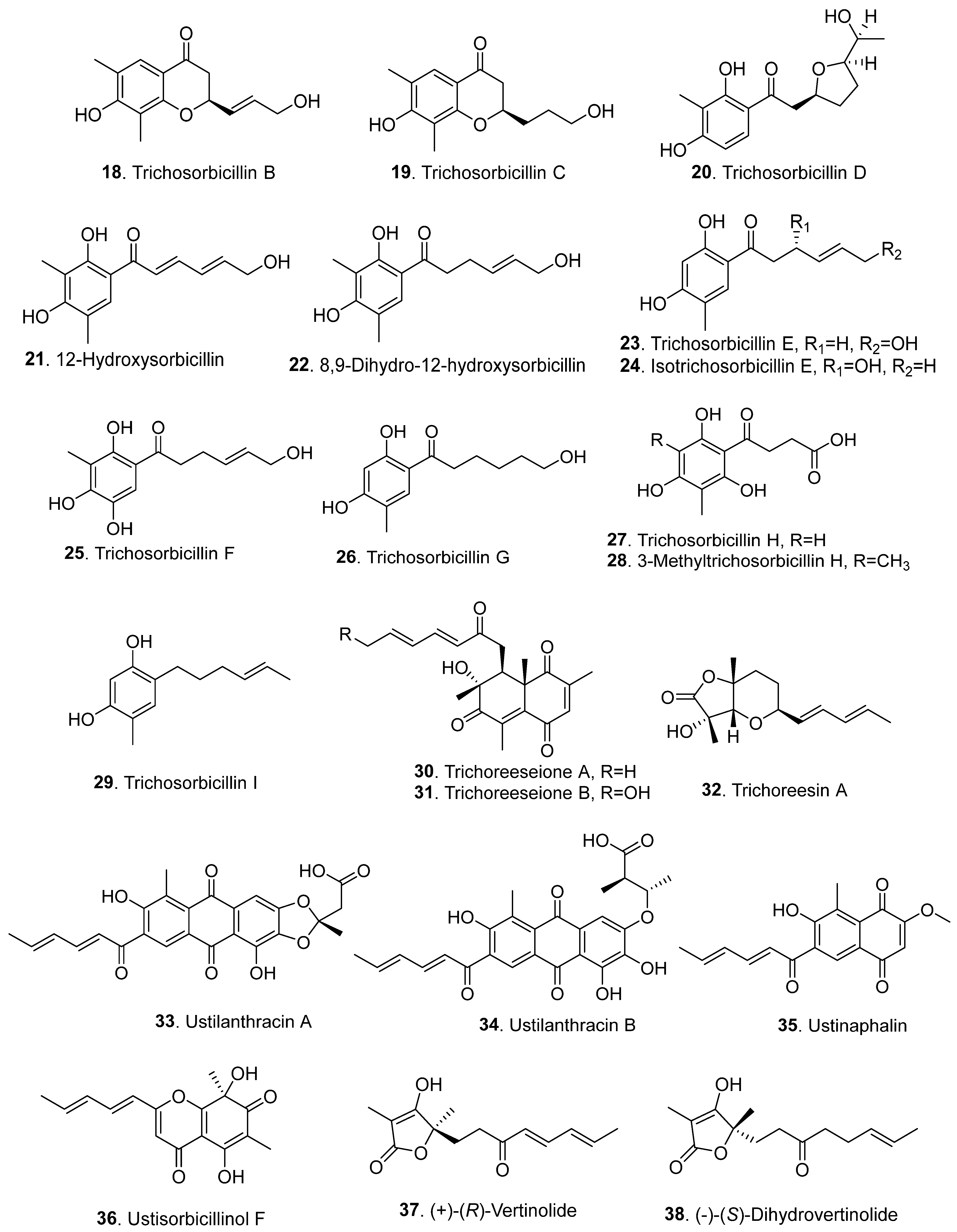{kind=link}
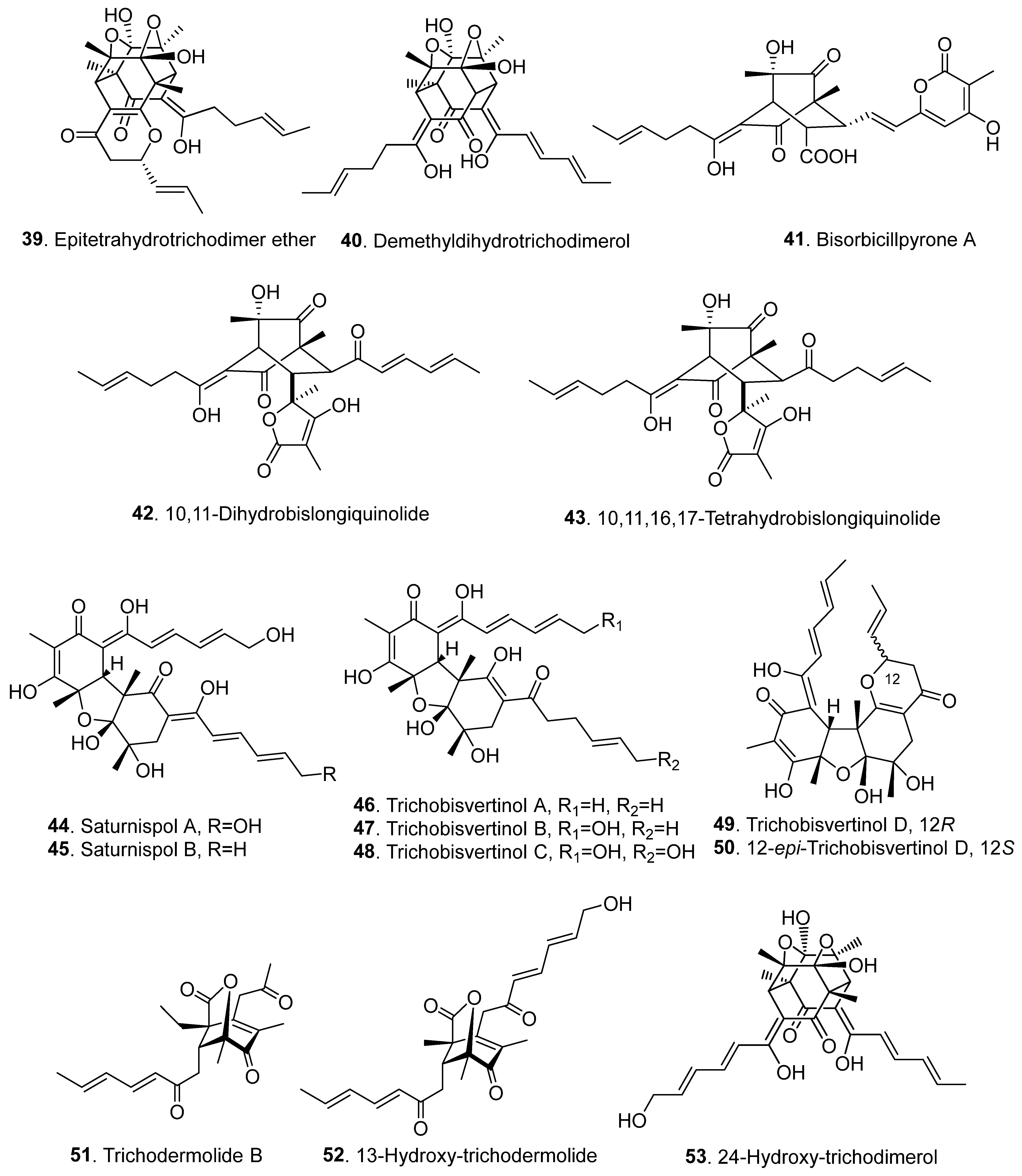{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Occurrence
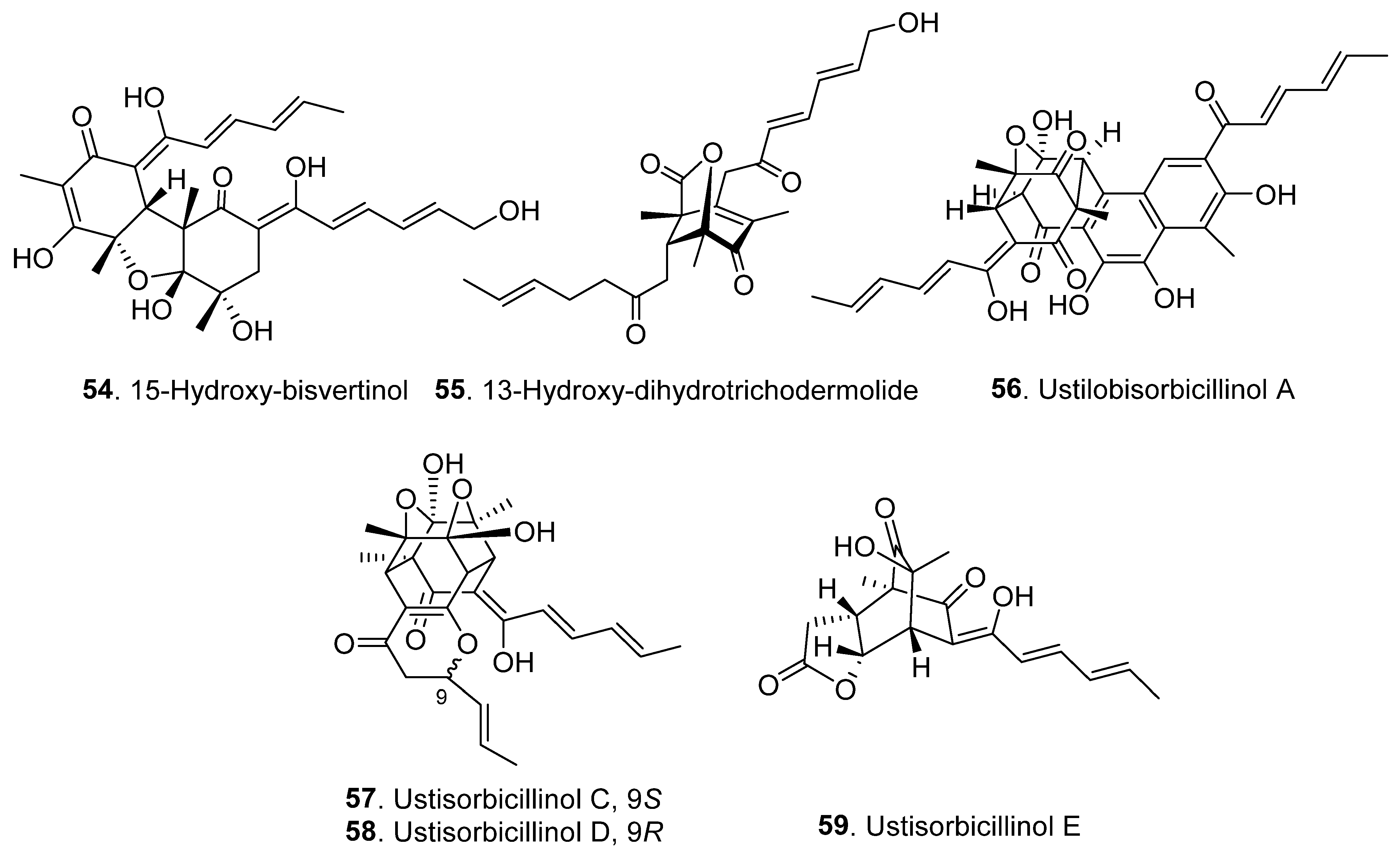
2.1. Monomeric Sorbicillinoids
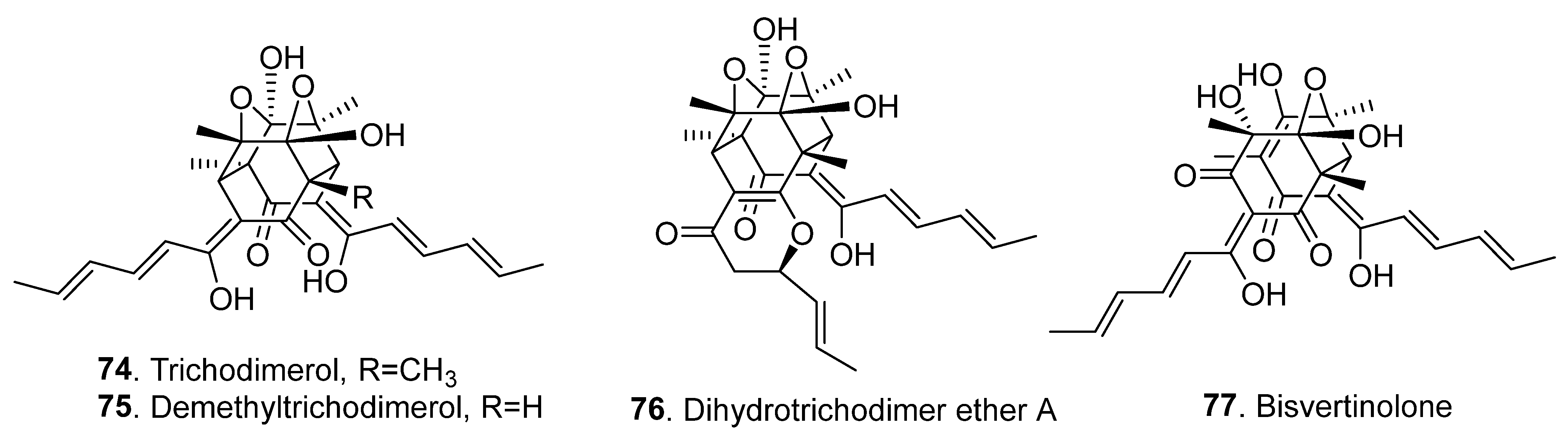
2.2. Bisorbicillinoids
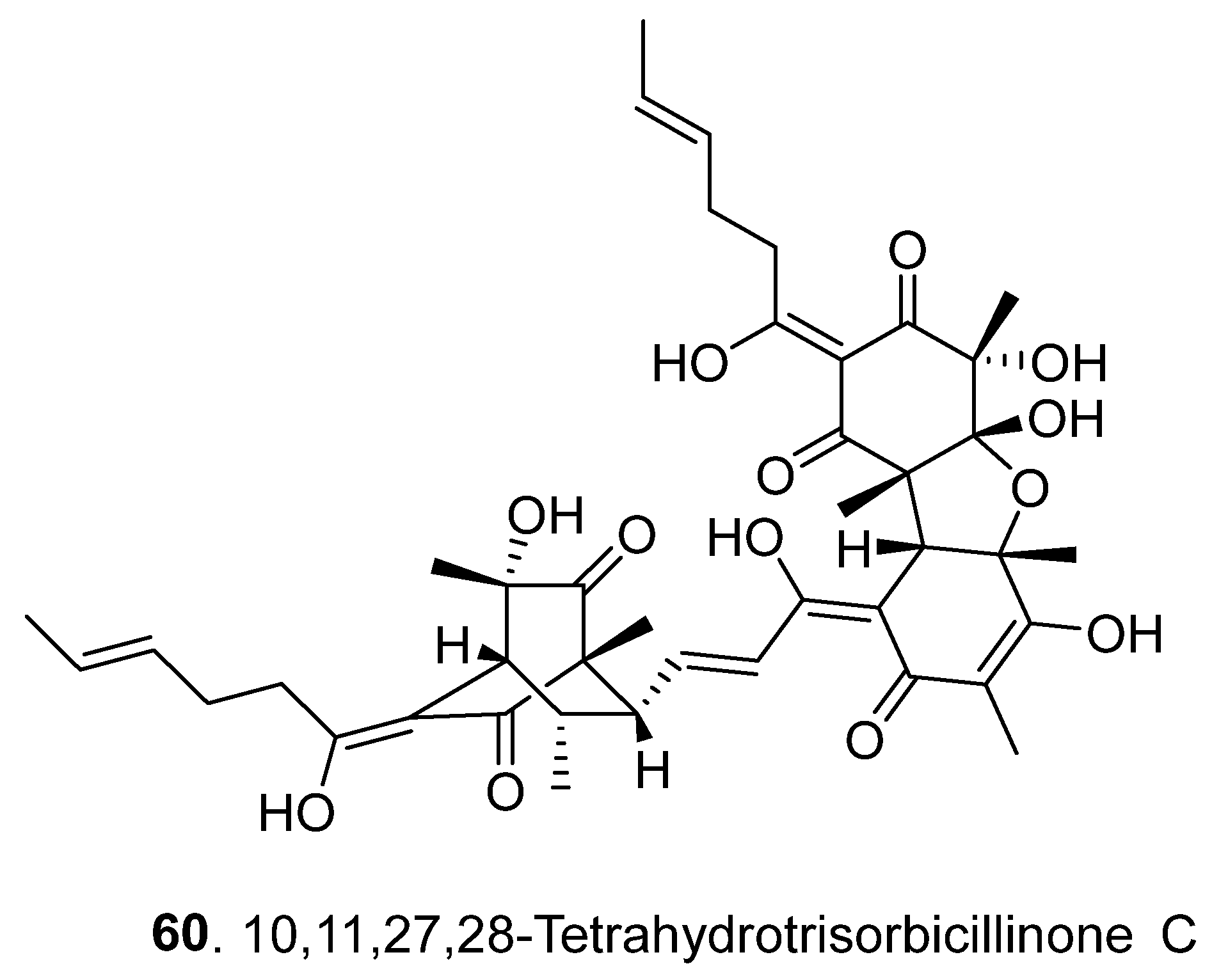
2.3. Trisorbicillinoids
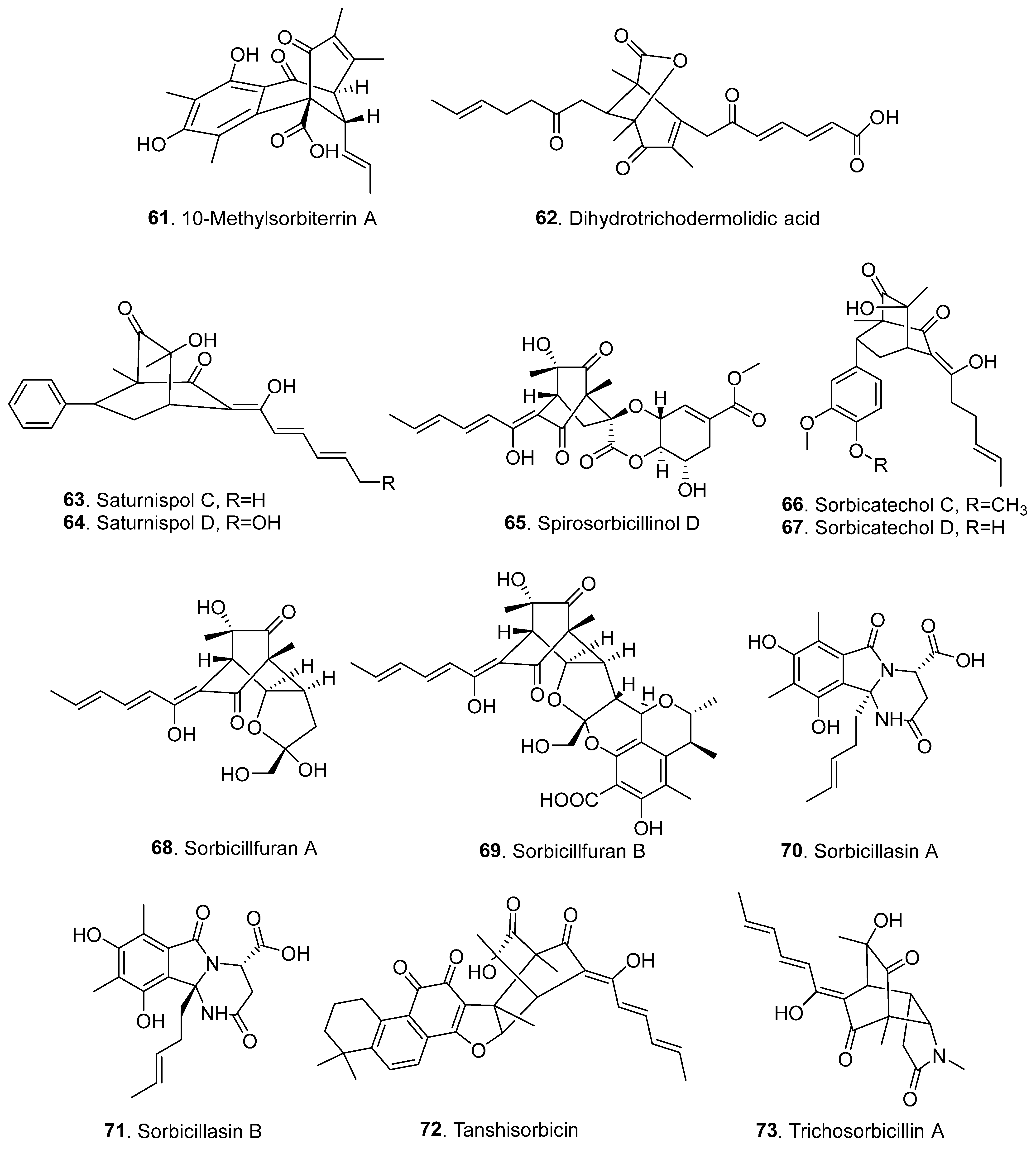
2.4. Hybrid Sorbicillinoids
3. Biological Activities
3.1. Cytotoxic Activity
3.2. Antibacterial Activity
3.3. Antifungal Activity
3.4. Anti-Inflammatory Activity
3.5. Phytotoxic Activity
3.6. α-Glucosidase Inhibitory Activity
3.7. Other Biological Activities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harned, A.M.; Volp, K.A. The sorbicillinoid family of natural products: Isolation, biosynthesis and synthetic studies. Nat. Prod. Rep. 2011, 28, 1790–1810. [Google Scholar] [CrossRef]
- Meng, J.; Wang, X.; Xu, D.; Fu, X.; Zhang, X.; Lai, D.; Zhou, L.; Zhang, G. Sorbicillinoids from fungi and their bioactivities. Molecules 2016, 21, 715. [Google Scholar] [CrossRef]
- Cram, D.J. Mold metabolites. II. The structure of sorbicillin, a pigment produced by the mold Penicillium notatum. J. Am. Chem. Soc. 1948, 70, 4240–4243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, S.-J.; Li, J.-J.; Liang, Z.-Z.; Zhao, C.-Q. Novel natural products from extremophilic fungi. Mar. Drugs 2018, 16, 194. [Google Scholar] [CrossRef] [Green Version]
- Han, P.; Zhang, X.; Xu, D.; Zhang, B.; Lai, D.; Zhou, L. Metabolites from Clonostachys fungi and their biological activities. J. Fungi 2020, 6, 229. [Google Scholar] [CrossRef] [PubMed]
- Derntl, C.; Guzman-Chavez, F.; Mello-de-Sousa, T.M.; Busse, H.-J.; Driessen, A.J.M.; Mach, R.L.; Mach-Aignr, A.R. In vivo study of the sorbicillinoid gene cluster in Trichoderma reeesei. Front. Mcrobiol. 2017, 8, 2037. [Google Scholar] [CrossRef]
- Guzman-Chavez, F.; Salo, O.; Nygard, Y.; Lankhorst, P.P.; Bovenberg, R.A.L.; Driessen, A.J.M. Mechanism and regulation of sorbicillin biosynthesis by Penicillium chrysorgenum. Microb. Biotechnol. 2017, 10, 958–968. [Google Scholar] [CrossRef] [PubMed]
- Al Fahad, A.; Abood, A.; Fisch, K.M.; Osipow, A.; Davison, J.; Avramovic, M.; Butts, C.P.; Piel, J.; Simpson, T.J.; Cox, R.J. Oxidative dearomatisation: The key step of sorbicillinoid biosynthesis. Chem. Sci. 2014, 5, 523–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.-S.; Zhou, Z.-F.; Yang, X.-H.; Lan, L.-F.; Gu, Y.-C.; Ye, B.-P.; Guo, Y.-W. Antibacterial sorbicillin and diketopiperazines from the endogenous fungus Penicillium sp. GD6 associated Chinese mangrove Bruguiera gymnorrhiza. Chin. J. Nat. Med. 2018, 16, 358–365. [Google Scholar] [CrossRef]
- Pang, X.; Zhou, X.; Lin, X.; Yang, B.; Tian, X.; Wang, J.; Xu, S.; Liu, Y. Structurally various sorbicillinoids from the deep-sea sediment derived fungus Penicillium sp. SCSIO06871. Bioorg. Chem. 2021, 107, 104600. [Google Scholar] [CrossRef]
- Ngo, M.T.; Nguyen, M.V.; Han, J.W.; Park, M.S.; Kim, H.; Choi, G.J. In vitro and in vivo antifungal activity of sorbicillinoids produced by Trichoderma longibrachiatum. J. Fungi 2021, 7, 428. [Google Scholar] [CrossRef]
- Ding, W.; Wang, F.; Li, Q.; Xue, Y.; Dong, Z.; Tian, D.; Chen, M.; Zhang, Y.; Hong, K.; Tang, J. Isolation and characterization of anti-inflammatory sorbicillinoids from the mangrove-derived fungus Penicillium sp. DM815. Chem. Biodivers. 2021, 18, e2100229. [Google Scholar] [CrossRef]
- Meng, J.; Cheng, W.; Heydari, H.; Wang, B.; Zhu, K.; Konuklugil, B.; Lin, W. Sorbicillinoid-based metabolites from a sponge-derived fungus Trichoderma saturnisporum. Mar. Drugs 2018, 16, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Gu, G.; Dang, P.; Zhang, X.; Wang, W.; Dai, J.; Liu, Y.; Lai, D.; Zhou, L. Sorbicillinoids from the fungus Ustilaginoidea virens and their phytotoxic, cytotoxic, and antimicrobial activities. Front. Chem. 2019, 7, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; He, Q.; Che, Q.; Zhang, G.; Zhu, T.; Gu, Q.; Li, D. Sorbicillasins A–B and scirpyrone K from a deep-sea-derived fungus, Phialocephala sp. FL30r. Mar. Drugs 2018, 16, 245. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Deng, Y.; Lin, X.; Chen, B.; Li, J.; Liu, H.; Chen, S.; Liu, L. Anti-inflammatory mono- and dimeric sorbicillinoids from the marine-derived fungus Trichoderma reesei 4670. J. Nat. Prod. 2019, 82, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Yang, L.-J.; Zhang, Y.-H.; Wu, J.-S.; Shi, T.; Halder, W.; Shao, C.-L.; Wang, C.-Y. Sorbicillinoid derivatives from sponge-derived fungus Trichoderma reesei (HN-2016-018). Front. Microbiol. 2020, 11, 1334. [Google Scholar] [CrossRef]
- Ma, X.-Y.; Shi, Z.-Z.; Ji, N.-Y. Sorbicillinoids from the alga-epiphytic fungus Trichoderma reesei Z56-8. Nat. Prod. Res. 2021. [Google Scholar] [CrossRef]
- Lai, D.; Meng, J.; Zhang, X.; Xu, D.; Dai, J.; Zhou, L. Ustilobisorbicillinol A, a cytotoxic sorbyl-containing aromatic polyketide from Ustilagninoidea virens. Org. Lett. 2019, 21, 1311–1314. [Google Scholar] [CrossRef]
- Sugaya, K.; Koshino, H.; Hongo, Y.; Yasunaga, K.; Onose, J.; Yoshikawa, K.; Abe, N. The biosynthesis of sorbicillinoids in Trichoderma sp. USF-2690: Prospect for the existence of a common precursor to sorbicillinol and 5-epihydroxyvertinolide, a new sorbicillinoid member. Tetrahedron Lett. 2008, 49, 654–657. [Google Scholar] [CrossRef]
- McMullin, D.R.; Renaud, J.B.; Barasubiye, T.; Sumarah, M.W.; Miller, J.D. Metabolites of Trichoderma species isolated from damp building materials. Can. J. Microbiol. 2017, 63, 621–632. [Google Scholar] [CrossRef]
- Supratman, U.; Suzuki, T.; Nakamura, T.; Yokoyama, Y.; Harneti, D.; Maharani, R.; Salam, S.; Abdullah, F.; Koseki, T.; Shiono, Y. New metabolites produced by endophyte Clonostachys rosea B5-2. Nat. Prod. Res. 2021, 35, 1525–1531. [Google Scholar] [CrossRef]
- Trifonov, L.S.; Bieri, J.H.; Prewo, R.; Dreiding, A.S.; Rast, D.M.; Hoesch, L. The constitution of vertinolide, a new derivative of tetronic acid, produced by Verticillium intertextum. Tetrahedron 1982, 38, 397–403. [Google Scholar] [CrossRef]
- Kahlert, L.; Bassiony, E.F.; Cox, R.J.; Shellam, E.J. Diels-Alder reactions during the biosynthesis of sorbicillinoids. Angew. Chem. Int. Ed. 2020, 59, 5816–5822. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Han, H.; Zhang, X.; Ma, C.; Sun, C.; Che, Q.; Gu, Q.; Zhu, T.; Zhang, G.; Li, D. Discovery of two new sorbicillinoids by overexpression of the global regulator LaeA in a marine-derived fungus Penicillium dipodomyis YJ-11. Mar. Drugs 2019, 17, 446. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.-J.; Zhu, T.; Liu, J.-T.; Ouyang, L.; Yang, F.; Lin, H.-W. New sorbicillinoid derivatives with GLP-1R and eEF2K affinities from a sponge-derived fungus Penicillium Chrysogenum 581F1. Nat. Prod. Res. 2020, 34, 2880–2886. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-L.; Zhang, D.; Lin, T.; He, Z.-H.; Yan, Q.-X.; Cai, Q.; Zhang, X.-K.; Yang, X.-W.; Chen, H.-F. Antiproliferative sorbicillinoids from the deep-sea-derived Penicillium allii-sativi. Front. Microbiol. 2021, 11, 636948. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, K.; Luo, X.; Wu, Z.; Gu, T.; Liao, S.; Lin, X.; Yang, B.; Liu, Y.; Fang, W.; et al. Sorbicillfurans A and B, two novel sorbicillinoid adducts from the fungus Penicillium citrinum SCSIO41402. Org. Biomol. Chem. 2019, 17, 8721. [Google Scholar] [CrossRef]
- He, W.; Liu, M.; Li, X.; Zhang, X.; Abdel-Mageed, W.M.; Li, L.; Wang, W.; Zhang, J.; Han, J.; Dai, H.; et al. Fungal biotransformation of tanshinone results in [4+2] cycloaddition with sorbicillinol: Evidence for enzyme catalysis and increased antibacterial activity. Appl. Microbiol. Biotechnol. 2016, 100, 8349–8357. [Google Scholar] [CrossRef]
- Lin, S.; Wu, Y.-Z.; Chen, K.-Y.; Ye, J.; Yang, X.-W.; Zhang, W.-D. Polyketides from the fungus Penicillium decumbens. J. Asian Nat. Prod. Res. 2018, 20, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Zhu, T.; Li, L.Y.; Cai, S.; Zhao, B.; Gu, Q. Cytotoxic sorbicillinoids and bisorbicillinoids from a marine-derived fungus Trichoderma sp. Chem. Pharm. Bull. 2009, 57, 220–223. [Google Scholar] [CrossRef] [Green Version]
- Ying, Y.-M.; Zhan, Z.-J.; Ding, Z.-S.; Shan, W.-G. Bioactive metabolites from Penicillium sp. P-1, a fungal endophyte in Huperzia serrata. Chem. Nat. Compd. 2011, 47, 541–544. [Google Scholar] [CrossRef]
- Yao, Y.; Li, J.; Jiang, C.-S.; Zhao, X.-X.; Miao, Z.-H.; Liu, H.-T.; Zheng, P.; Yao, W.-X.; Li, W.-Q. Trichodimerol and sorbicillin induced apoptosis of HL-60 cells is mediated by reactive oxygen species. Pharmazie 2015, 70, 394–398. [Google Scholar]
- Maskey, R.P.; Grün-Wollny, I.; Grün-Wollny, H. Sorbicillin analogues and related dimeric compounds from Penicillium notatum. J. Nat. Prod. 2005, 68, 865–870. [Google Scholar] [CrossRef]
- Corral, P.; Esposito, F.P.; Tedesco, P.; Falco, A.; Tortorella, E.; Tartaglione, L.; Festa, C.; D’Auria, M.V.; Gnavi, G.; Varese, G.C.; et al. Identification of a sorbicillinoid-producing Aspergillus strain with antimicrobial activity against Staphylococcus aureus: A new polyextremophilic marine fungus from Barents Sea. Mar. Biotechnol. 2018, 20, 502–511. [Google Scholar] [CrossRef]
- Meng, J.; Wang, B.; Cheng, W. Study on the secondary metabolites of Thichoderma sturnisporum. Chin. J. Mar. Drugs 2017, 36, 27–31. [Google Scholar]
- Yang, Z.; Qiao, Y.; Li, J.; Wu, F.-G.; Lin, F. Novel type of water-soluble photosensitizer from Trichoderma reesei for photodynamic inactivation of Gram-positive bacteria. Langmuir 2020, 36, 13227–13235. [Google Scholar] [CrossRef]
- Venkatesha, S.H.; Achaya, B.; Moudgil, K.D. Natural products as source of anti-inflammatory drugs. Inflammation 2017, 4, 1661–1690. [Google Scholar]
- Zhao, J.-L.; Zhang, M.; Liu, J.-M.; Tan, Z.; Chen, R.-D.; Xie, K.-B.; Dai, J.-G. Bioactive steroids and sorbicillinoids isolated from the endophytic fungus Trichoderma sp. Xy24. J. Asian Nat. Prod. Res. 2017, 19, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Marra, R.; Nicoletti, R.; Pagano, E.; DellaGreca, M.; Salvatore, M.M.; Borrelli, F.; Lombardi, N.; Vinale, F.; Woo, S.L.; Andolfi, A. Inhibitory effect of trichodermanone C, a sorbicillinoid produced by Trichoderma citrinoviride associated to the green alga Cladophora sp., on nitrite production in LPS-stimulated macrophages. Nat. Prod. Res. 2019, 33, 3389–3397. [Google Scholar] [CrossRef]
- Xu, D.; Xue, M.; Shen, Z.; Jia, X.; Hou, X.; Lai, D.; Zhou, L. Phytotoxic secondary metabolites from fungi. Toxins 2021, 13, 261. [Google Scholar] [CrossRef]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-derived bioactive compounds produced by endophytic fungi. Mini-Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef]
- Sivakumar, P.M.; Prabhawathi, V.; Zarrabi, A.; Akthar, S.; Prabhakar, P.K. Current trends in the therapeutic strategies for diabetes management. Curr. Med. Chem. 2021, 28, 4616–4637. [Google Scholar] [CrossRef]
- Deng, J.; Dai, H.; Wang, Y.; Chen, H.; Tan, Z.; Mei, W. Isolation and identification of the fungus Aspergillus sp. HNWSW-20 from Chinese agarwood and its secondary metabolites. Chin. J. Trop. Crops 2018, 39, 1618–1624. [Google Scholar]
- Abe, N.; Murata, T.; Hirota, A. Novel DPPH radical scavengers, bisorbicillinol and demethyltrichodimerol, from a fungus. Biosci. Biotechnol. Biochem. 1998, 62, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Sugaya, K.; Terajima, T.; Takahashi, A.; Onose, J.; Abe, N. Bisorbicillinol inhibits Lyn tyrosine kinase for allergic response on RBL-2H3 cells. Bioorg. Med. Chem. Lett. 2019, 29, 832–835. [Google Scholar] [CrossRef] [PubMed]
- Tilki, E.K.; Ozturk, S.E.; Ozarda, M.G.; Canturk, Z.; Dikmen, M. Investigation of the neuroprotective and neuritogenic effects of halotolerant Penicillium flavigenum-derived sorbicillin-like compounds on PC-12 Adh cells. Cytotechnology 2021, 73, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Tang, X.; Qiao, D.; Li, P.; Li, G. Studies on bioactive components of a marine-derived fungus Penicillium chrysogenum. Chin. J. Mar. Drugs 2019, 38, 77–81. [Google Scholar]
- Lu, S.; Sun, W.; Meng, J.; Wang, A.; Wang, X.; Tian, J.; Fu, X.; Dai, J.; Liu, Y.; Lai, D.; et al. Bioactive bis-naphtho-γ-pyrones from rice false smut pathogen Ustilaginoidea virens. J. Agric. Food Chem. 2015, 63, 3501–3508. [Google Scholar] [CrossRef]
- Meng, J.; Sun, W.; Mao, Z.; Xu, D.; Wang, X.; Lu, S.; Lai, D.; Liu, Y.; Zhou, L.; Zhang, G. Main ustilaginoidins and their distribution in rice false smut balls. Toxins 2015, 7, 4023–4034. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Dong, X.; Xu, D.; Meng, J.; Fu, X.; Wang, X.; Lai, D.; Zhou, L.; Liu, Y. Preparative separation of main ustilaginoidins from rice false smut balls by high-speed counter-current chromatography. Toxins 2016, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Fu, X.; Lin, F.; Sun, W.; Meng, J.; Wang, A.; Lai, D.; Zhou, L.; Liu, Y. The contents of ustiloxins A and B along with their distribution in rice false smut balls. Toxins 2016, 8, 262. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Lai, D.; Wang, W.; Dai, J.; Zhou, L.; Liu, Y. Ustiloxin G, a new cyclopeptide mycotoxin from rice false smut balls. Toxins 2017, 9, 54. [Google Scholar] [CrossRef]
- Sun, W.; Wang, A.; Xu, D.; Wang, W.; Meng, J.; Dai, J.; Liu, Y.; Lai, D.; Zhou, L. New ustilaginoidins from rice false smut balls caused by Villosiclava virens and their phytotoxic and cytotoxic activities. J. Agric. Food Chem. 2017, 65, 5151–5160. [Google Scholar] [CrossRef]
- Lai, D.; Meng, J.; Xu, D.; Zhang, X.; Liang, Y.; Han, Y.; Jiang, C.; Liu, H.; Wang, C.; Zhou, L.; et al. Determination of the absolute configurations of the stereogenic centers of ustilaginoidins by studying the biosynthetic monomers from a gene knockout mutant of Villosiclava virens. Sci. Rep. 2019, 9, 1855. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Li, P.; Han, P.; Gu, G.; Shan, T.; Lai, D.; Zhou, L. New nitrogen-containing metabolites from cultures of rice false smut pathogen Villosiclava virens. Nat. Prod. Res. 2021, 35, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Zhao, S.; Dang, P.; Zhou, Z.; Lai, D.; Zhou, L. Ustilaginoidin M1, a new bis-naphtho-γ-pyrone from the fungus Villosiclava virens. Nat. Prod. Res. 2021, 35, 1555–1560. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Yin, R.; Zhou, Z.; Gu, G.; Zhao, S.; Xu, J.-R.; Liu, J.; Peng, Y.; Lai, D.; Zhou, L. Elucidation of ustilaginoidin biosynthesis reveals a previously unrecognised class of ene-reductases. Chem. Sci. 2021, 12, 14883–14892. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lin, F.; Sun, W.; Yuan, S.; Zhou, Z.; Wu, F.-G.; Chen, Z. Constitutive hyperproduction of sorbicillinoids in Trichoderma reesei ZC121. Biotechnol. Biofuels 2018, 11, 291. [Google Scholar] [CrossRef]
- Ramesh, C.; Vinithkumar, N.V.; Kirubagraran, R.; Venil, C.K.; Dufosse, L. Multifaceted applications of microbial pigments: Current knowledge, challenges and future directions for public health implications. Microorganisms 2019, 7, 186. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Xu, J. Fungal pigments and their roles associated with human health. J. Fungi 2020, 6, 280. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Kubicek, E.M.; Kubicek, C.P. Several steps of lateral gene transfer followed by events of ‘birth-and-death’ evolution shaped a fungal sorbicillinoid biosynthetic gene cluster. BMC Evol. Biol. 2016, 16, 269. [Google Scholar] [CrossRef] [Green Version]
- Salo, O.; Guzman-Chavez, F.; Ries, M.I.; Lankhorst, P.P.; Bovenberg, R.A.L.; Vreeken, R.J.; Driessen, A.J.M. Identification of a polyketide synthase involved in sorbicillin biosynthesis by Penicillium chrysogenum. Appl. Environ. Microbiol. 2016, 82, 3971–3978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Chu, J. Characterization of two polyketide synthases involved in sorbicillinoid biosynthesis by Acremonium chrysogenum using the CRISPR/Cas9 system. Appl. Biochem. Biotechnol. 2019, 188, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Kahlert, L.; Cox, R.J.; Skellam, E. The same but different: Multiple functions of the fungal flavin dependent monooxygenase SorD from Penicillium chrysogenum. Chem. Commun. 2020, 56, 10934. [Google Scholar] [CrossRef] [PubMed]
- Monroy, A.A.; Stappler, E.; Schuster, A.; Sulyok, M.; Schmoll, M. A CRE1-regulated cluster is responsible for light dependent production of dihydrotrichotetronin in Trichoderma reesei. PLoS ONE 2017, 12, e0182530. [Google Scholar] [CrossRef] [Green Version]
- Hinterdobler, W.; Schuster, A.; Tisch, D.; Ozkan, E.; Bazafkan, H.; Schinnerl, J.; Brecker, L.; Bohmdorfer, S.; Schmoll, M. The role of PKAc1 in gene regulation and trichodimerol production in Trichoderma reesei. Fungal Biol. Biotechnol. 2019, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Hitzenhammer, E.; Buschl, C.; Sulyok, M.; Schuhmacher, R.; Kluger, B.; Wischnitzki, E.; Schmoll, M. YPR2 is a regulator of light modulated carbon and secondary metabolism in Trichoderma reesei. BMC Genom. 2019, 20, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sib, A.; Gulder, T.A.M. Stereoselective total synthesis of bisorbicillinoid natural products by enzymatic oxidative dearomatization/dimerization. Angew. Chem. Int. Ed. 2017, 56, 12888–12891. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Banwell, M.G.; Coote, M.L.; Lee, R.; Willis, A.C. Establishing the true structure of the sorbicillinoid-derived isolate rezishanone C by total synthesis. Chem. Asian J. 2017, 12, 1480–1484. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, X.; Zhang, X.; Xue, M.; Zhao, Z.; Zhang, H.; Xu, D.; Lai, D.; Zhou, L. Recent Advances in Sorbicillinoids from Fungi and Their Bioactivities (Covering 2016–2021). J. Fungi 2022, 8, 62. https://doi.org/10.3390/jof8010062
Hou X, Zhang X, Xue M, Zhao Z, Zhang H, Xu D, Lai D, Zhou L. Recent Advances in Sorbicillinoids from Fungi and Their Bioactivities (Covering 2016–2021). Journal of Fungi. 2022; 8(1):62. https://doi.org/10.3390/jof8010062
Chicago/Turabian StyleHou, Xuwen, Xuping Zhang, Mengyao Xue, Zhitong Zhao, Huizhen Zhang, Dan Xu, Daowan Lai, and Ligang Zhou. 2022. "Recent Advances in Sorbicillinoids from Fungi and Their Bioactivities (Covering 2016–2021)" Journal of Fungi 8, no. 1: 62. https://doi.org/10.3390/jof8010062