
Transcription Factors Pmr1 and Pmr2 Cooperatively Regulate Melanin Biosynthesis, Conidia Development and Secondary Metabolism in Pestalotiopsis microspora

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Growth Conditions
2.2. Bioinformatic Analysis
2.3. Gene Cloning, Plasmid Construction and Genetic Manipulation
2.4. RNA Preparation and Quantitative Real-Time PCR
2.5. Phenotype Observation
2.6. Conidia Quantification and Secondary Metabolite Profiling
3. Results
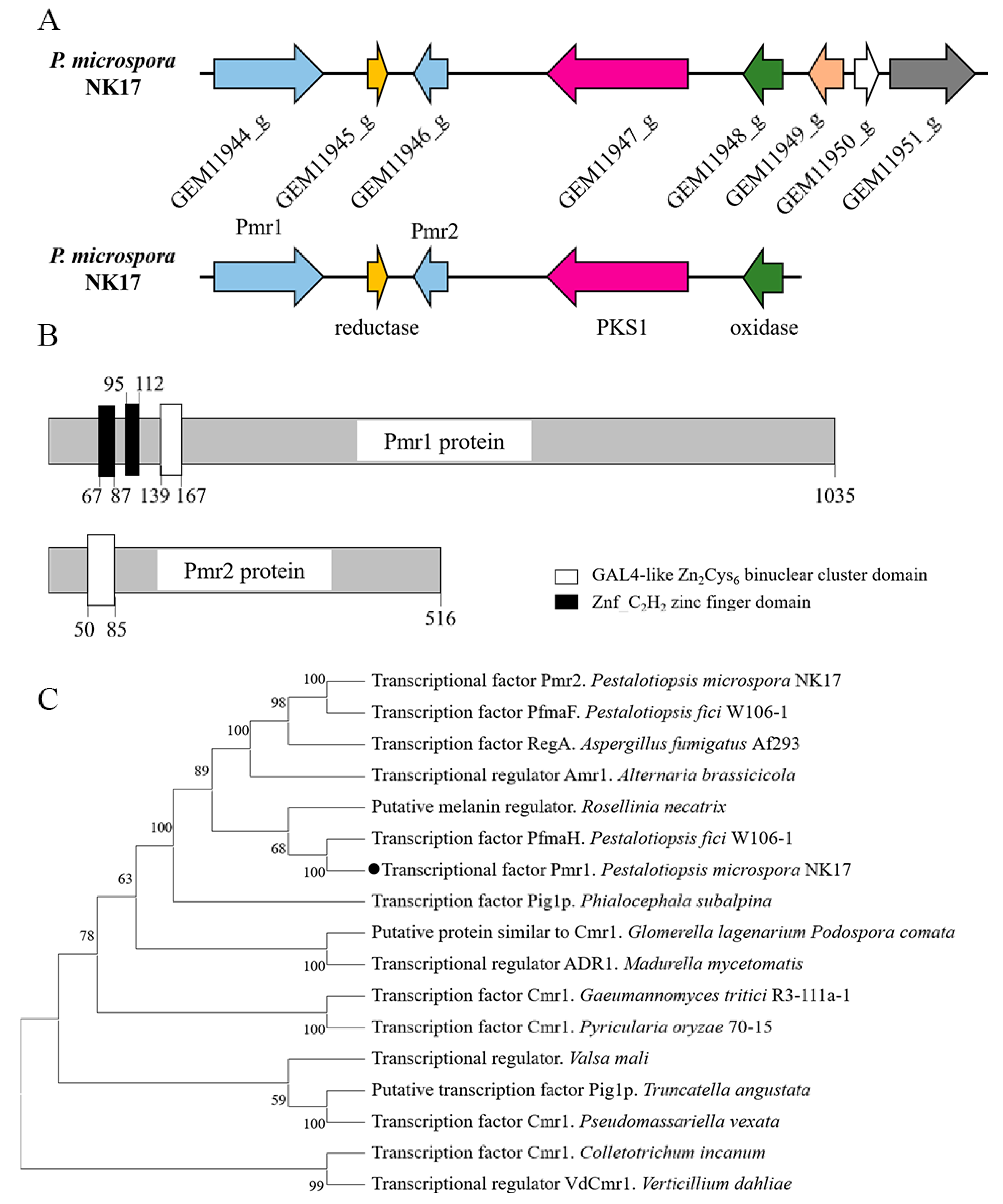
3.1. Chracterization of the Transcription Factors in the Gene Cluster of Melanin Biosynthesis
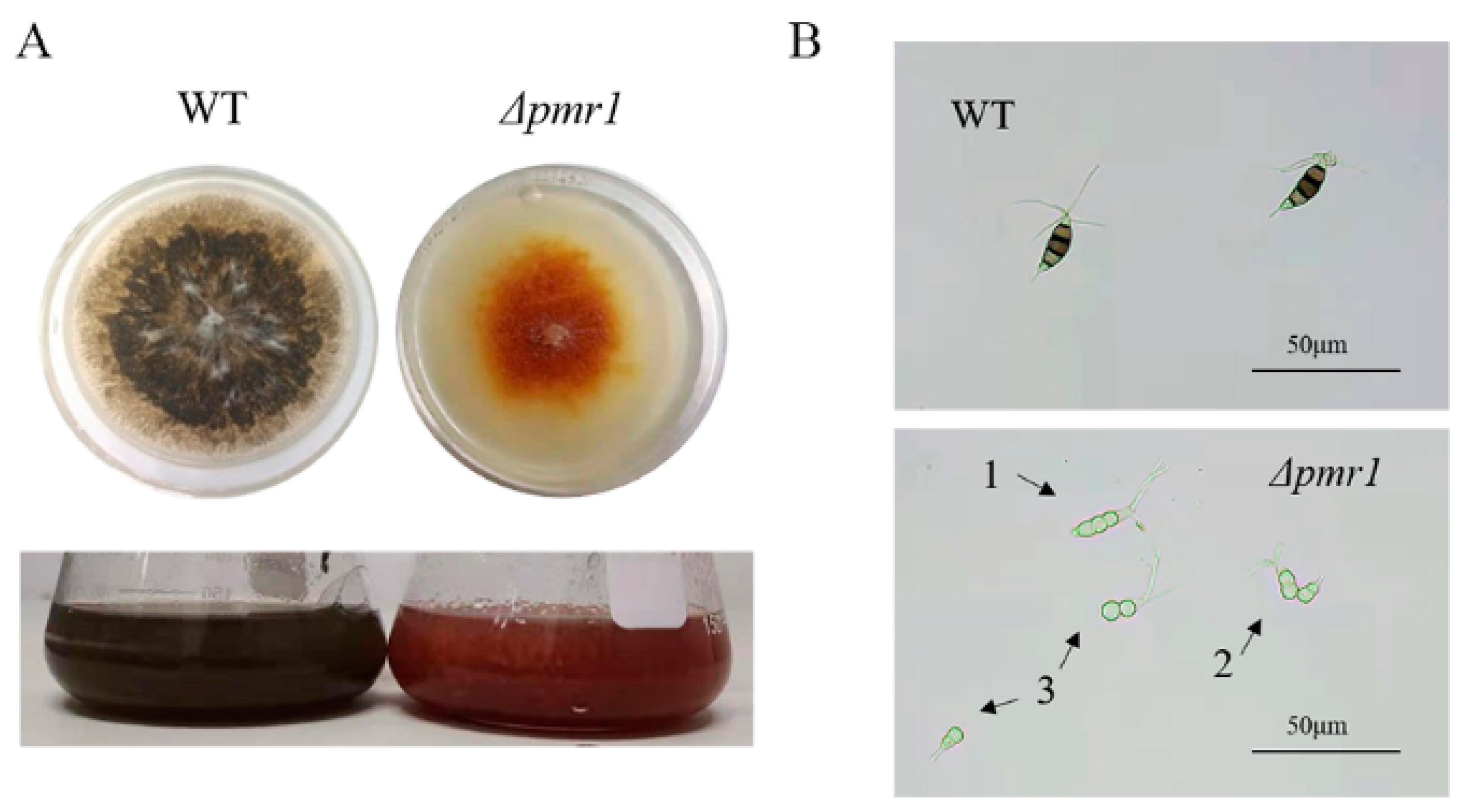
3.2. Effects of Pmr1 on Conidial Pigmentation and Morphogenesis
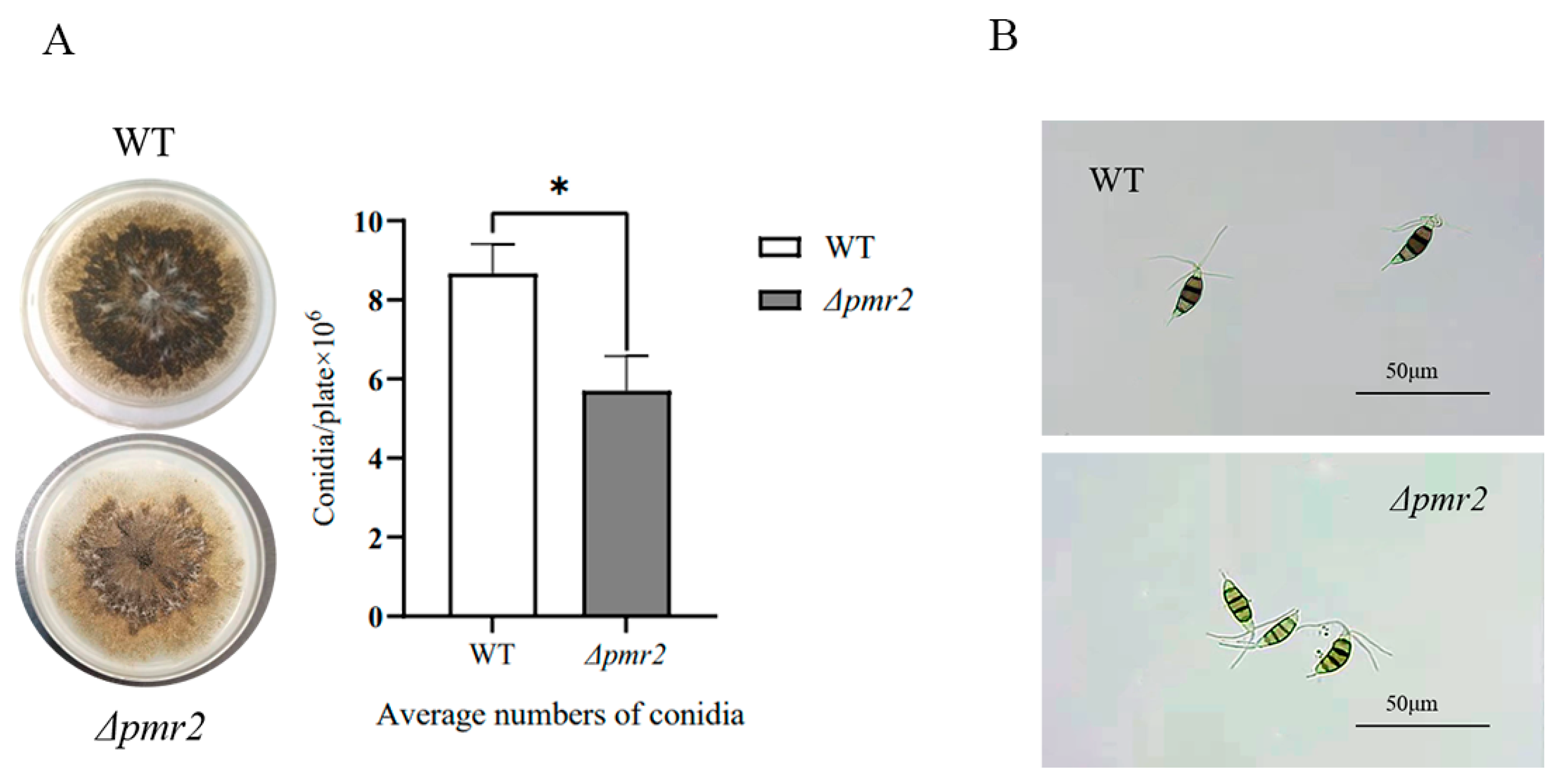
3.3. Pmr2 Influences Conidial Pigmentation and Conidia Production
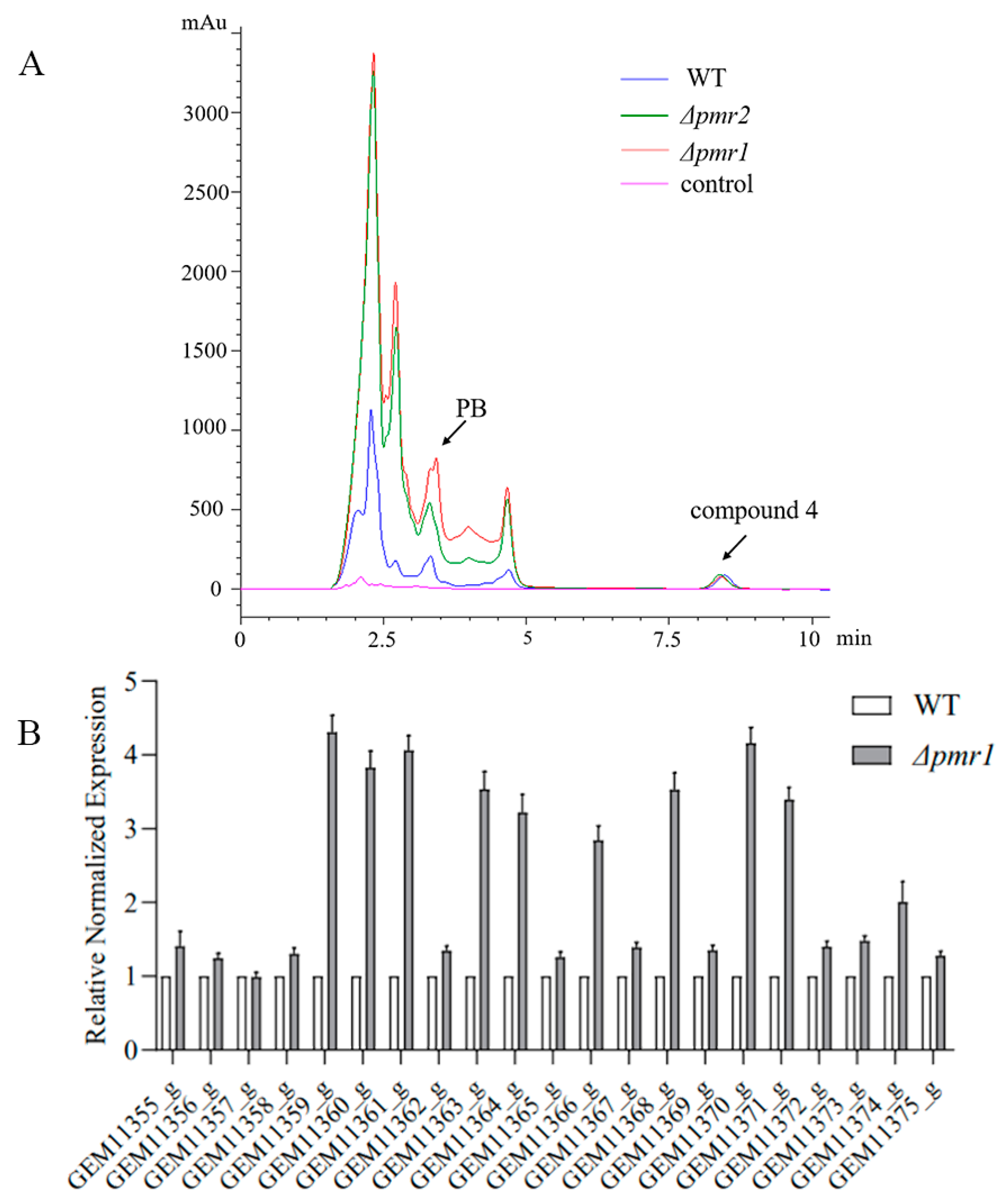
3.4. Roles of Pmr1 and Pmr2 in Biosynthesis of Secondary Metabolites
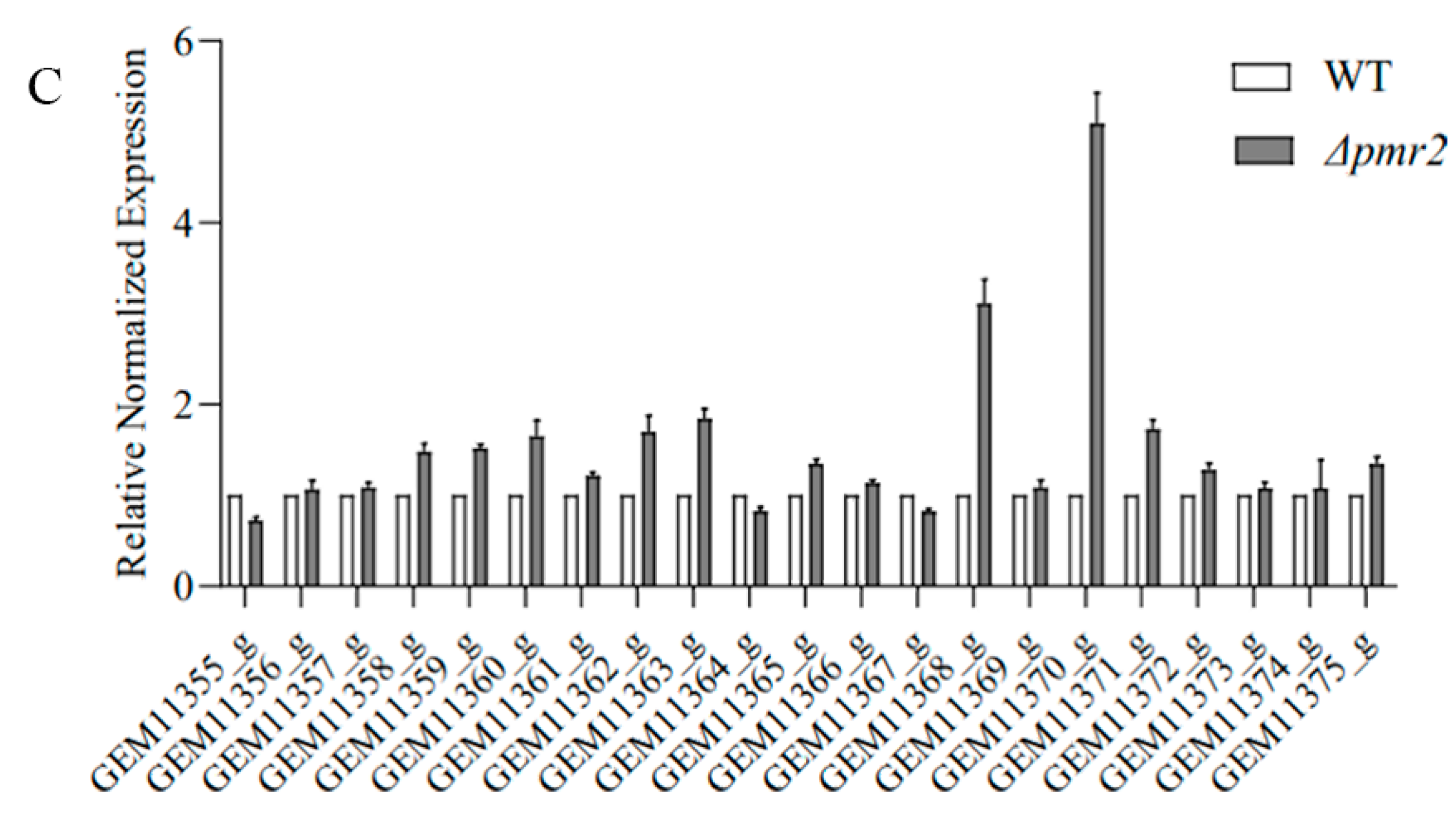
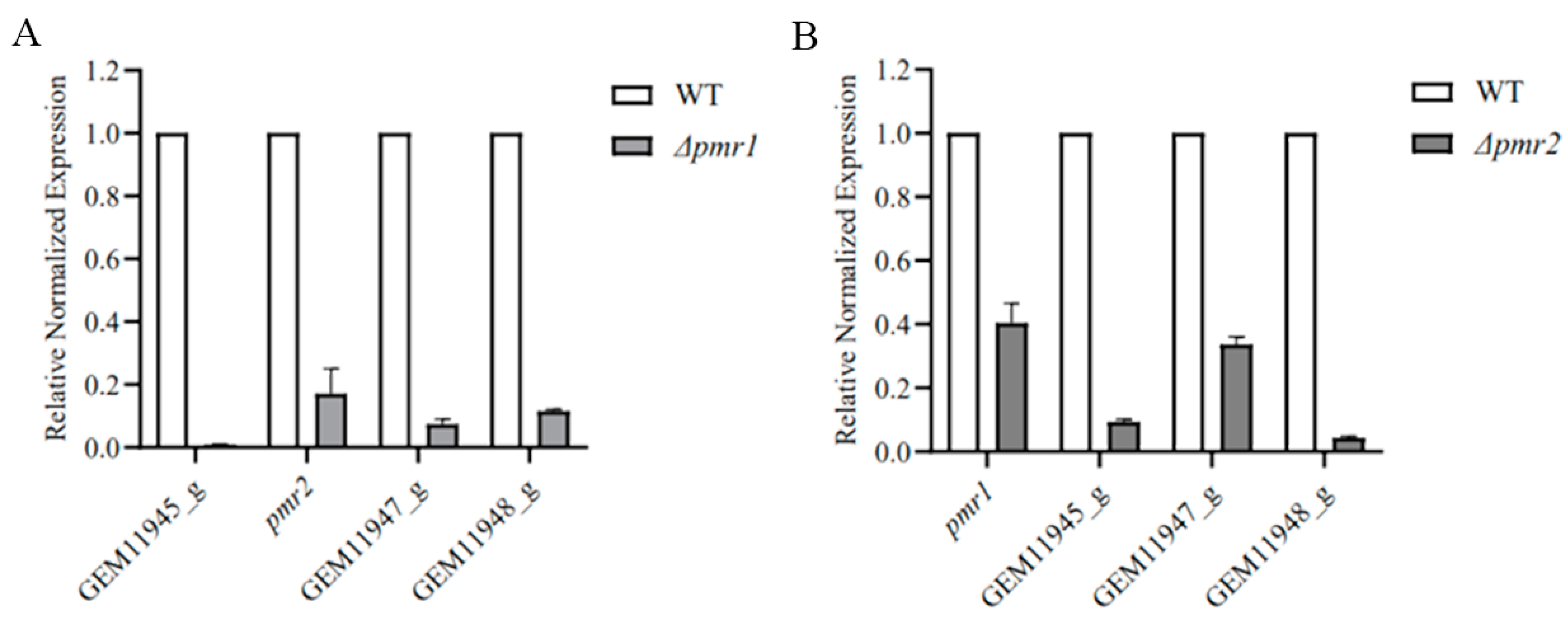
3.5. Pmr1 and Pmr2 Both Regulate the Expression of Genes in Melanin Biosynthesis Cluster
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Camacho, E.; Vij, R.; Chrissian, C.; Prados-Rosales, R.; Gil, D. The structural unit of melanin in the cell wall of the fungal pathogen Cryptococcus neoformans. J. Biol. Chem. 2019, 294, 10471–10489. [Google Scholar] [CrossRef]
- Eisenman, H.C.; Greer, E.M.; McGrail, C.W. The role of melanins in melanotic fungi for pathogenesis and environmental survival. Appl. Microbiol. Biotechnol. 2020, 104, 4247–4257. [Google Scholar] [CrossRef] [PubMed]
- Heidrich, D.; Corbellini, V.A.; Mendes, S.D.C.; Fernandes, E.K.; Lazzarotto, L.; Ribeiro, A.C.; Zanette, R.A.; Scroferneker, M.L. Melanin: Quantification and protection against oxidative stress in chromoblastomycosis agents. Med. Mycol. 2019, 57, 260–263. [Google Scholar] [CrossRef]
- Mironenko, N.V.; Alekhina, I.A.; Zhdanova, N.N.; Bulat, S.A. Intraspecific variation in gamma-radiation resistance and genomic structure in the filamentous fungus Alternaria alternata: A case study of strains inhabiting Chernobyl reactor no. 4. Ecotoxicol. Environ. Saf. 2000, 45, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Kaewmalakul, J.; Nosanchuk, J.D.; Vanittanakom, N.; Youngchim, S. Melanization and morphological effects on antifungal susceptibility of Penicillium marneffei. Antonie Leeuwenhoek 2014, 106, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Chand, R.; Dubey, R.S.; Shah, K. Effect of Tricyclazole on morphology, virulence and enzymatic alterations in pathogenic fungi Bipolaris sorokiniana for management of spot blotch disease in barley. World J. Microbiol. Biotechnol. 2015, 31, 23–35. [Google Scholar] [CrossRef]
- Eisenman, H.C.; Nosanchuk, J.D.; Webber, J.B.; Emerson, R.J.; Camesano, T.A.; Casadevall, A. Microstructure of cell wall-associated melanin in the human pathogenic fungus Cryptococcus neoformans. Biochemistry 2005, 44, 3683–3693. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wang, X.; Fan, A.; Zheng, Y.; Liu, X. A cryptic pigment biosynthetic pathway uncovered by heterologous expression is essential for conidial development in Pestalotiopsis fic. Mol. Microbiol. 2017, 105, 469–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A. antiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, 339–346. [Google Scholar] [CrossRef]
- Yoshitaka, T.; Yasuyuki, K.; Itaru, K.; Iwao, F. Temporal Transcriptional Pattern of Three Melanin Biosynthesis Genes, PKS1, SCD1, and THR1, in Appressorium-Differentiating and Nondifferentiating Conidia of Colletotrichum lagenarium. Appl. Environ. Microbiol. 1997, 63, 351–354. [Google Scholar]
- Cho, Y.; Srivastava, A.; Ohm, R.A.; Lawrence, C.B.; Wang, K.H. Transcription factor Amr1 induces melanin biosynthesis and suppresses virulence in Alternaria brassicicola. PLoS Pathog. 2012, 8, e1002974. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yang, L.; Wu, M.; Chen, W.; Li, G. A Single-Nucleotide Deletion in the Transcription Factor Gene bcsmr1 Causes Sclerotial-Melanogenesis Deficiency in Botrytis cinerea. Front. Microbiol. 2017, 8, 2492. [Google Scholar] [CrossRef]
- Zhang, P.; Zhou, S.; Wang, G.; An, Z.; Liu, X. Two transcription factors cooperatively regulate DHN melanin biosynthesis and development in Pestalotiopsis fici. Mol. Microbiol. 2019, 112, 649–666. [Google Scholar] [CrossRef]
- Yu, X.; Huo, L.; Liu, H.; Chen, L.; Wang, Y. Melanin is required for the formation of the multi-cellular conidia in the endophytic fungus Pestalotiopsis microspora. Microbiol. Res. 2015, 179, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.N.; Ji, Y.; Pan, J.; Yu, Y.; Chen, H.; Zhu, X. A new taxol-producing fungus (Pestalotiopsis malicola) and evidence for taxol as a transient product in the culture. Afr. J. Biotechnol. 2011, 10, 6647–6654. [Google Scholar]
- Blin, K.; Pascal, A.V.; de-Los, S.E.L.C.; Del, C.F.; Lee, S.Y.; Medema, M.H.; Weber, T. The antiSMASH database version 2: A comprehensive resource on secondary metabolite biosynthetic gene clusters. Nucleic Acids Res. 2019, 47, D625–D630. [Google Scholar] [CrossRef] [Green Version]
- Paz, Z.; Garcia-Pedrajas, M.D.; Andrews, D.L.; Klosterman, S.J.; Baeza-Montanez, L. One step construction of Agrobacterium-Recombination-ready-plasmids (OSCAR), an efficient and robust tool for ATMT based gene deletion construction in fungi. Fungal Genet. Biol. 2011, 48, 677–684. [Google Scholar] [CrossRef]
- Wang, D.; Akhberdi, O.; Hao, X.R.; Chen, L.F.; Liu, Y.J.; Zhu, X.D. Amino acid sensor kinase Gcn2 is required for conidiation, secondary metabolism, and cell wall integrity in the Taxol-producer Pestalotiopsis microspora. Front. Microbiol. 2017, 8, 1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pihet, M.; Vandeputte, P.; Tronchin, G.; Renier, G.; Saulnier, P. Melanin is an essential component for the integrity of the cell wall of Aspergillus fumigatus conidia. BMC Microbiol. 2009, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, L.; Xie, Q.; Yu, X.; Duan, A. A Gene Cluster for the Biosynthesis of Dibenzodioxocinons in the Endophyte Pestalotiopsis microspora, a Taxol Producer. J. Microbiol. Biotechnol. 2019, 29, 1570–1579. [Google Scholar] [CrossRef] [PubMed]
- Engh, I.; Nowrousian, M.; Kück, U. Regulation of melanin biosynthesis via the dihydroxynaphthalene pathway is dependent on sexual development in the ascomycete Sordaria macrospora. FEMS Microbiol. 2007, 275, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Plonka, P.M.; Grabacka, M. Melanin synthesis in microorganisms-biotechnological and medical aspects. Acta Biochim. Pol. 2006, 53, 429–443. [Google Scholar] [CrossRef] [Green Version]
- Kihara, J.; Moriwaki, A.; Tanaka, N.; Tanaka, C.; Ueno, M. Characterization of the BMR1 gene encoding a transcription factor for melanin biosynthesis genes in the phytopathogenic fungus Bipolaris oryza. FEMS Microbiol. Lett. 2008, 281, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Hu, X.; Fang, Y.; Anchieta, A.; Goldman, P.H. Transcription factor VdCmr1 is required for pigment production, protection from UV irradiation, and regulates expression of melanin biosynthetic genes in Verticillium dahliae. Microbiology 2018, 164, 685–696. [Google Scholar] [CrossRef]
- Franzen, A.J.; Cunha, M.M.; Batista, E.J.; Seabra, S.H.; Rozental, S. Effects of tricyclazole (5-methyl-1,2,4-triazol[3,4] benzothiazole), a specific DHN-melanin inhibitor, on the morphology of Fonsecaea pedrosoi conidia and sclerotic cells. Microsc. Res. Tech. 2006, 69, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Hao, X.R.; Lou, J.; Zhang, P.; Pan, J.; Zhu, X.D. A PKS gene, pks-1, is involved in chaetoglobosin biosynthesis, pig-mentation and sporulation in Chaetomium globosum. Sci. China Life Sci. 2012, 55, 1100–1108. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Oide, S.; Zhang, N.; Choi, M.Y.; Turgeon, B.G. ChLae1 and ChVel1 regulate T-toxin production, virulence, oxidative stress response, and development of the of the maize pathogen Cochliobolus heterostrophus. PLoS Pathog. 2012, 8, e1002542. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Li, Z.; Liu, Y.; Zhang, P.; Hao, X.; Zhu, X. Transcription Factors Pmr1 and Pmr2 Cooperatively Regulate Melanin Biosynthesis, Conidia Development and Secondary Metabolism in Pestalotiopsis microspora. J. Fungi 2022, 8, 38. https://doi.org/10.3390/jof8010038
Zhou M, Li Z, Liu Y, Zhang P, Hao X, Zhu X. Transcription Factors Pmr1 and Pmr2 Cooperatively Regulate Melanin Biosynthesis, Conidia Development and Secondary Metabolism in Pestalotiopsis microspora. Journal of Fungi. 2022; 8(1):38. https://doi.org/10.3390/jof8010038
Chicago/Turabian StyleZhou, Mengkai, Ze Li, Yanjie Liu, Ping Zhang, Xiaoran Hao, and Xudong Zhu. 2022. "Transcription Factors Pmr1 and Pmr2 Cooperatively Regulate Melanin Biosynthesis, Conidia Development and Secondary Metabolism in Pestalotiopsis microspora" Journal of Fungi 8, no. 1: 38. https://doi.org/10.3390/jof8010038