
Variations in the Community Structure of Fungal Microbiota Associated with Apple Fruit Shaped by Fruit Bagging-Based Practice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Orchard Site and Apple Sample Collection
2.2. Sampling
2.3. Bioinformatic Analysis
2.4. Statistical Analysis
- Chao1 estimator [55]:where n1 and n2 are the count of one sequence (i.e., singletons) and two sequences (i.e., doubletons), respectively, and Sobs is the observed number of species.
- Shannon estimator [56]:where Sobs is the number of observed OTUs, N = the total number of individuals in the community, and ni is the number of individuals in OUTi.
- Simpson estimator [57]:where Sobs is the number of observed OTUs or species in the community, N = the total number of individuals in the community, and ni is the number of individuals in OUTi.
3. Results
3.1. Characterization of High-Throughput Sequencing Data
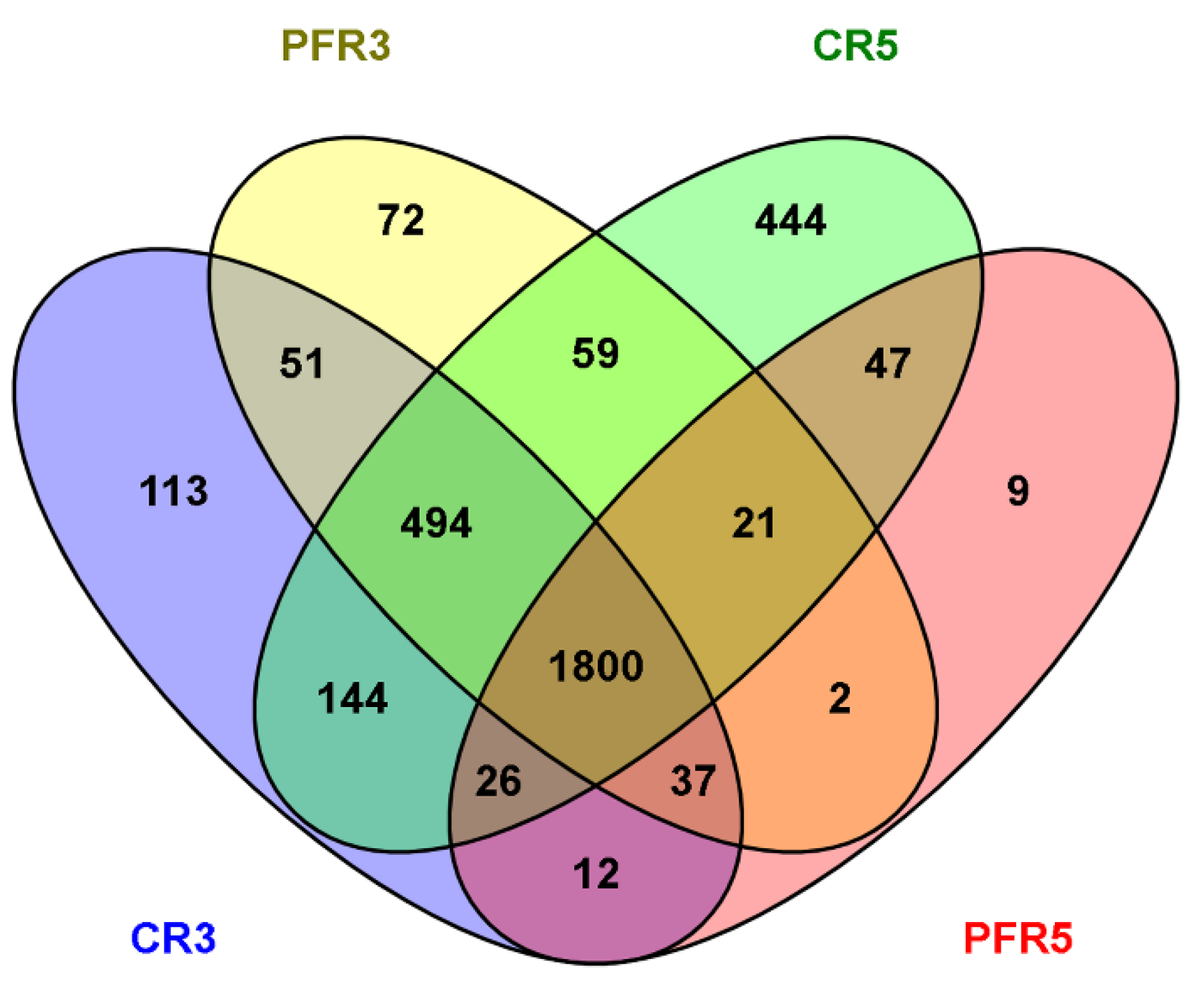
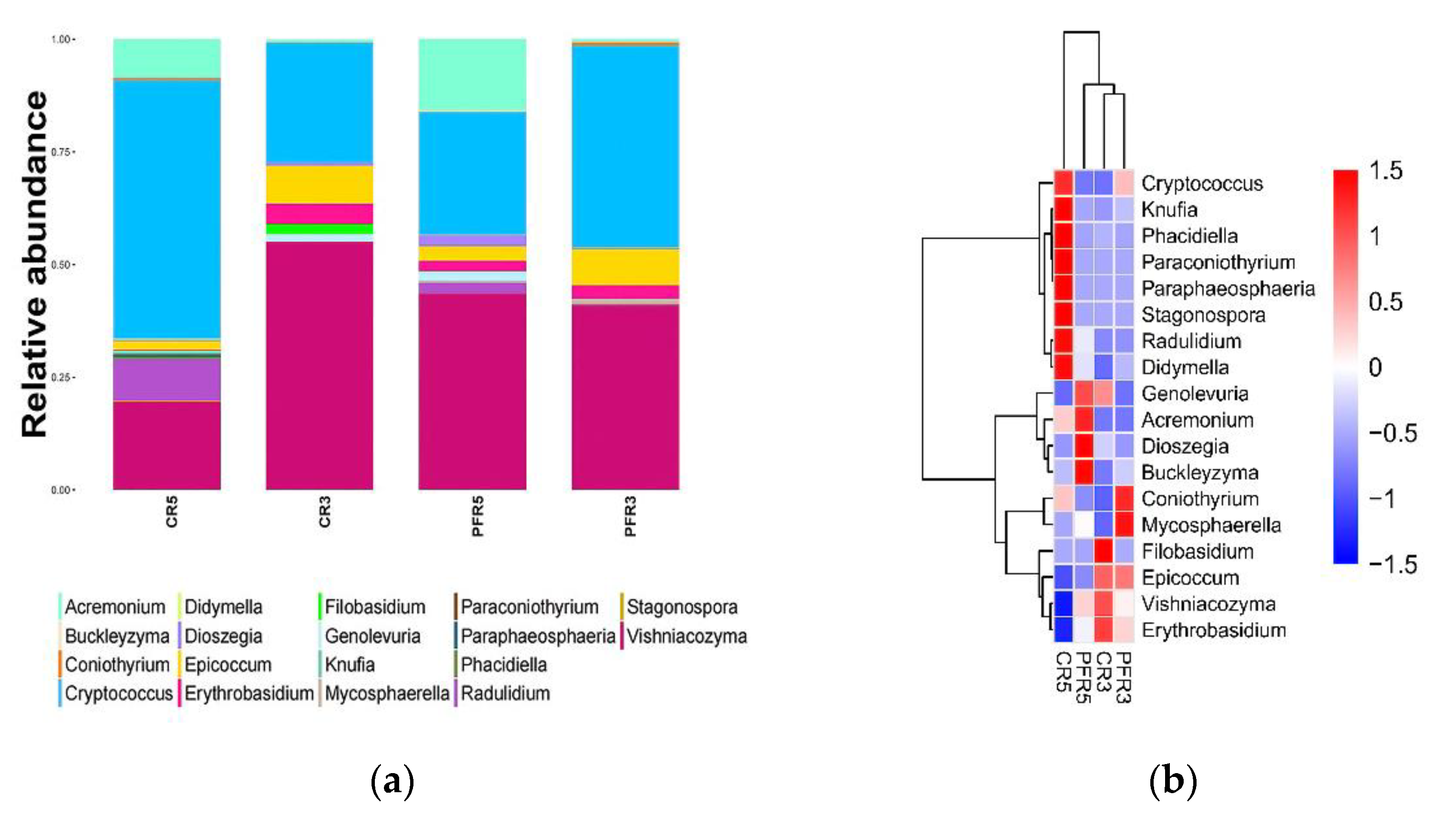
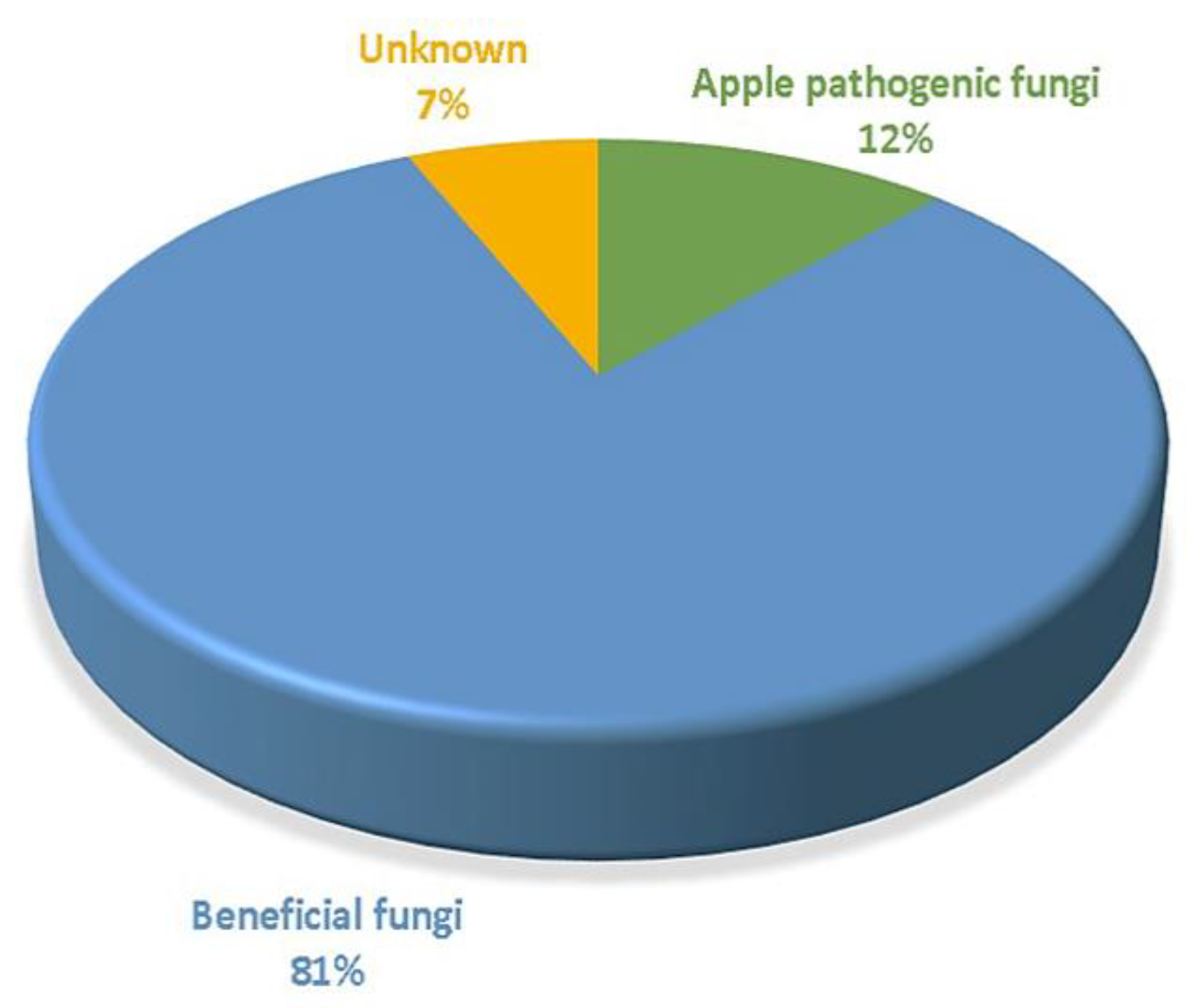
3.2. An Overall Attribution of Fungal Community Composition
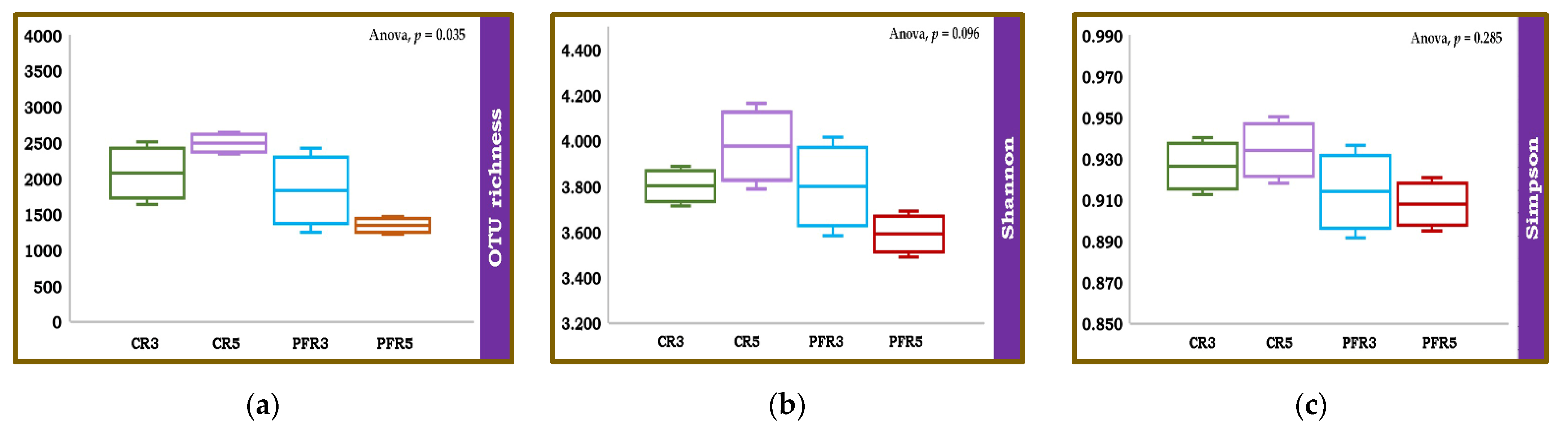
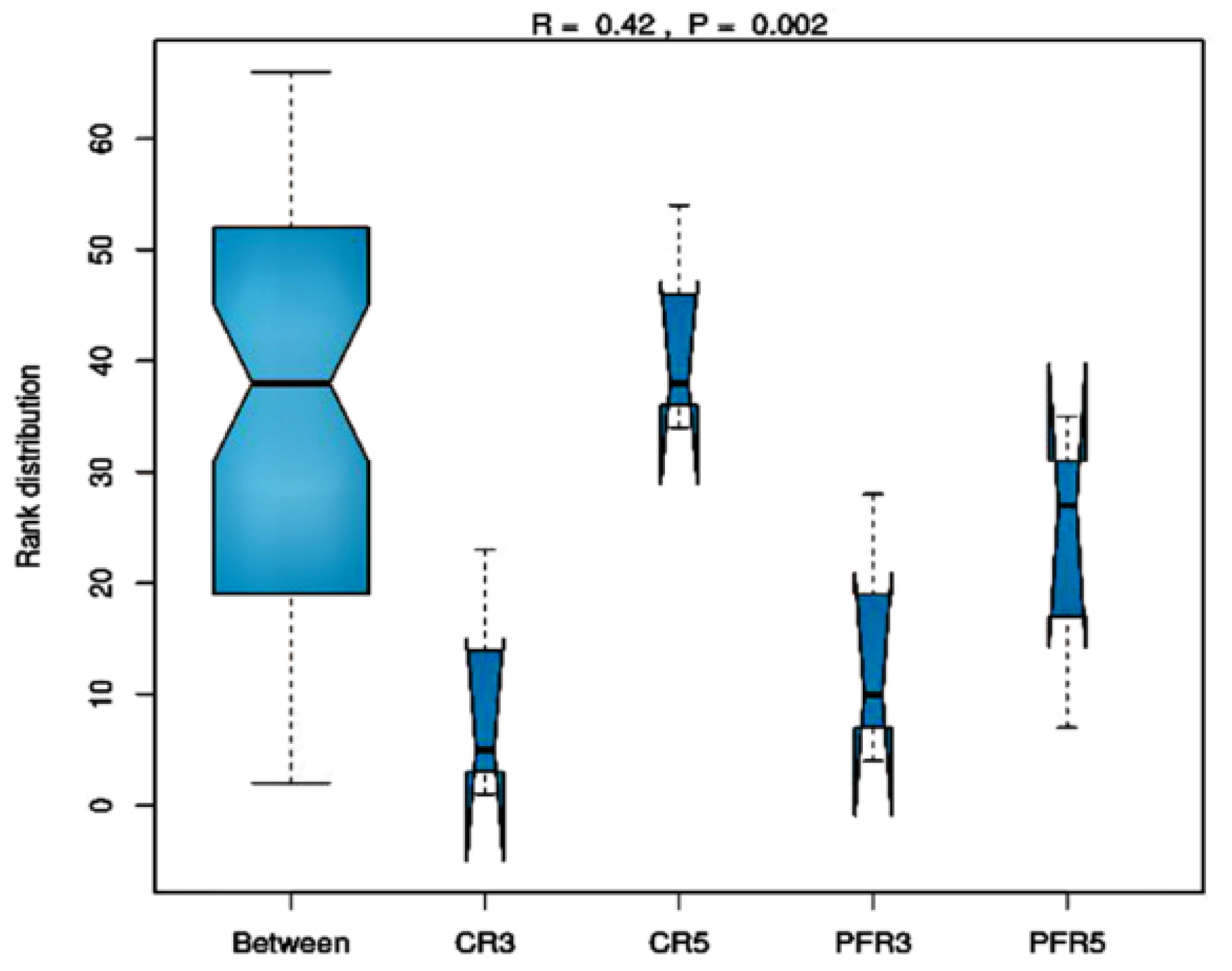
3.3. Fungal Richness and Diversity among Apple Fruit Samples
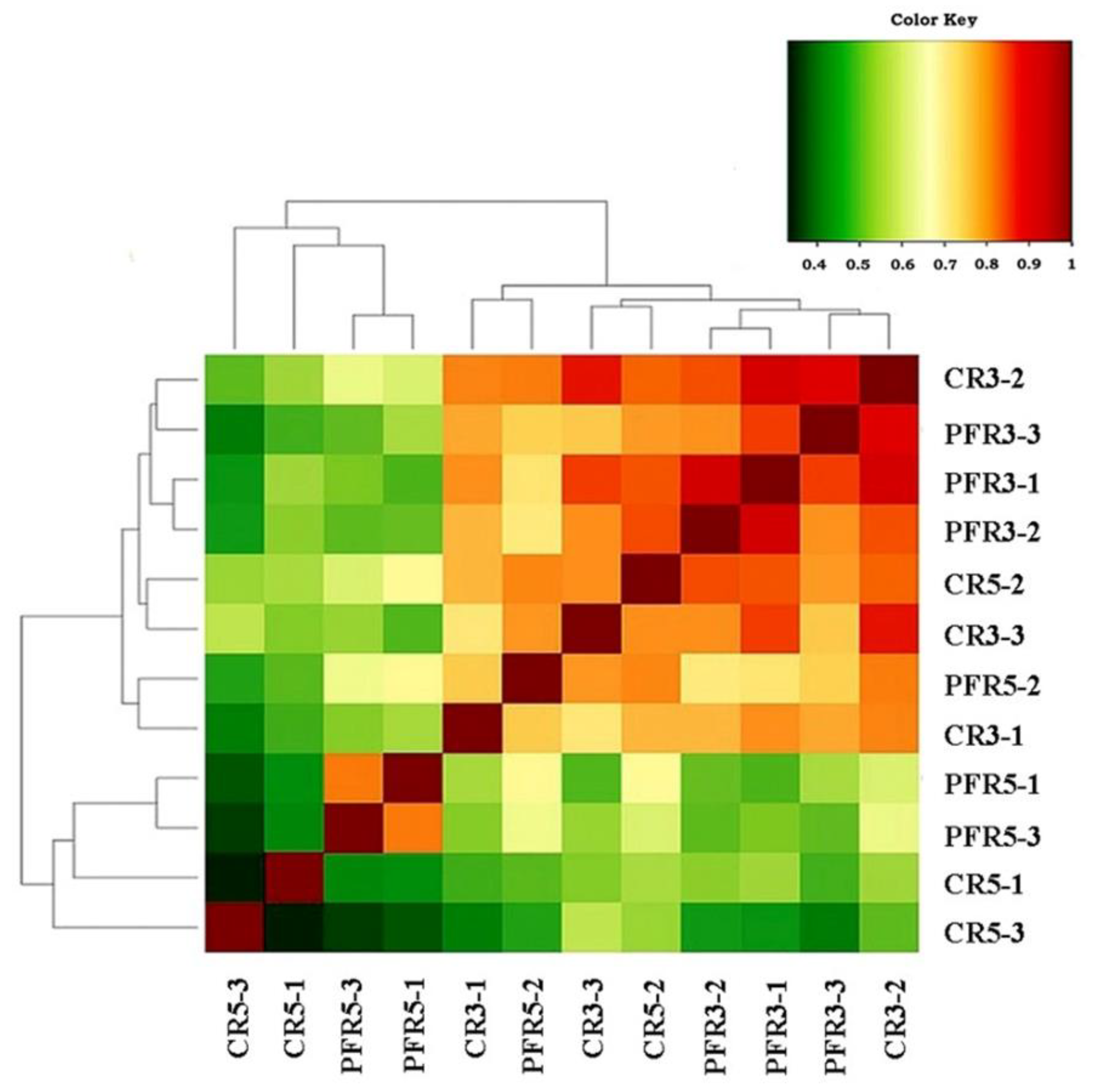
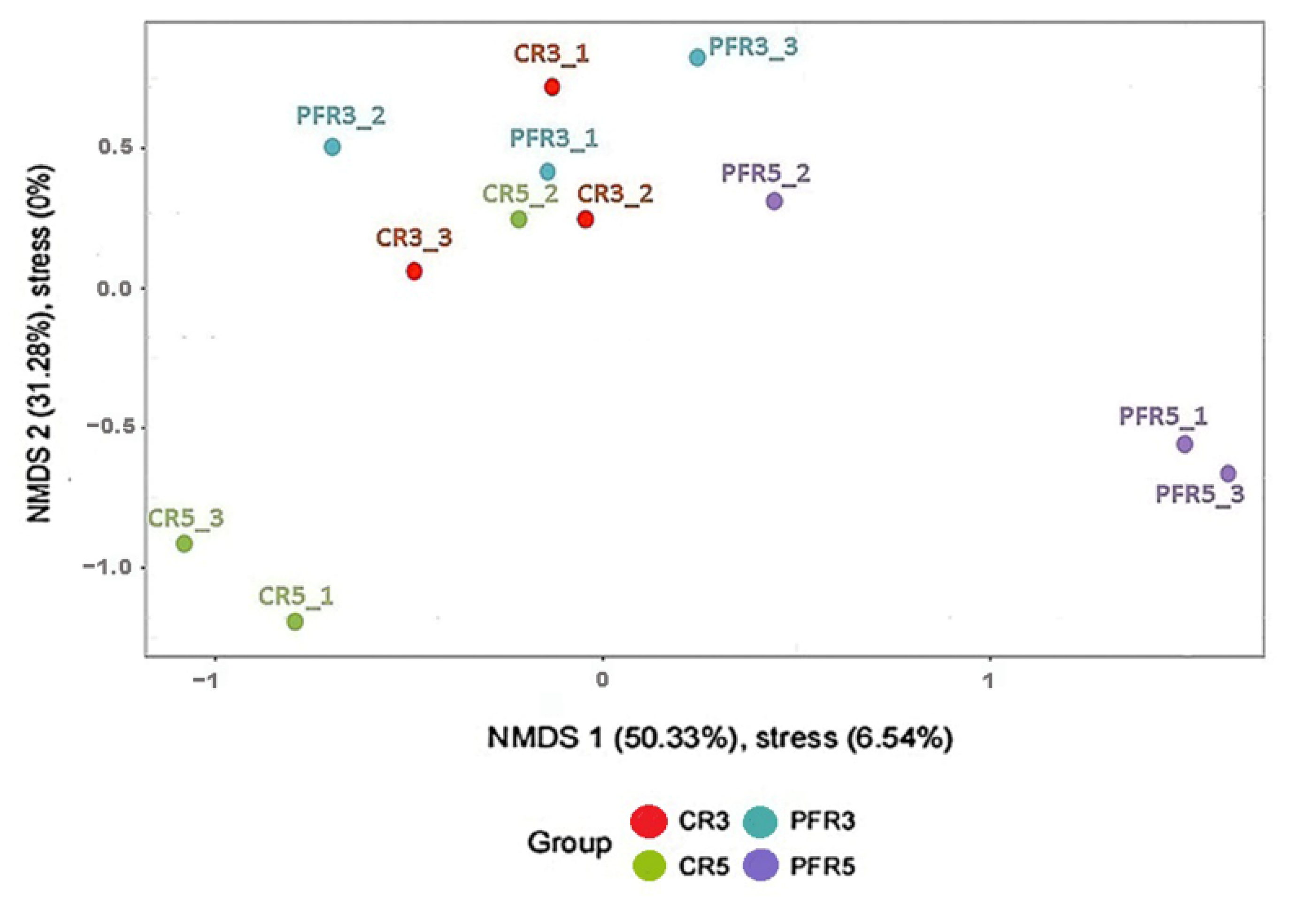
3.4. Analysis of Fungal Variation between Calyx-End and Peel from Bagging- and Non-Bagging-Based Practices
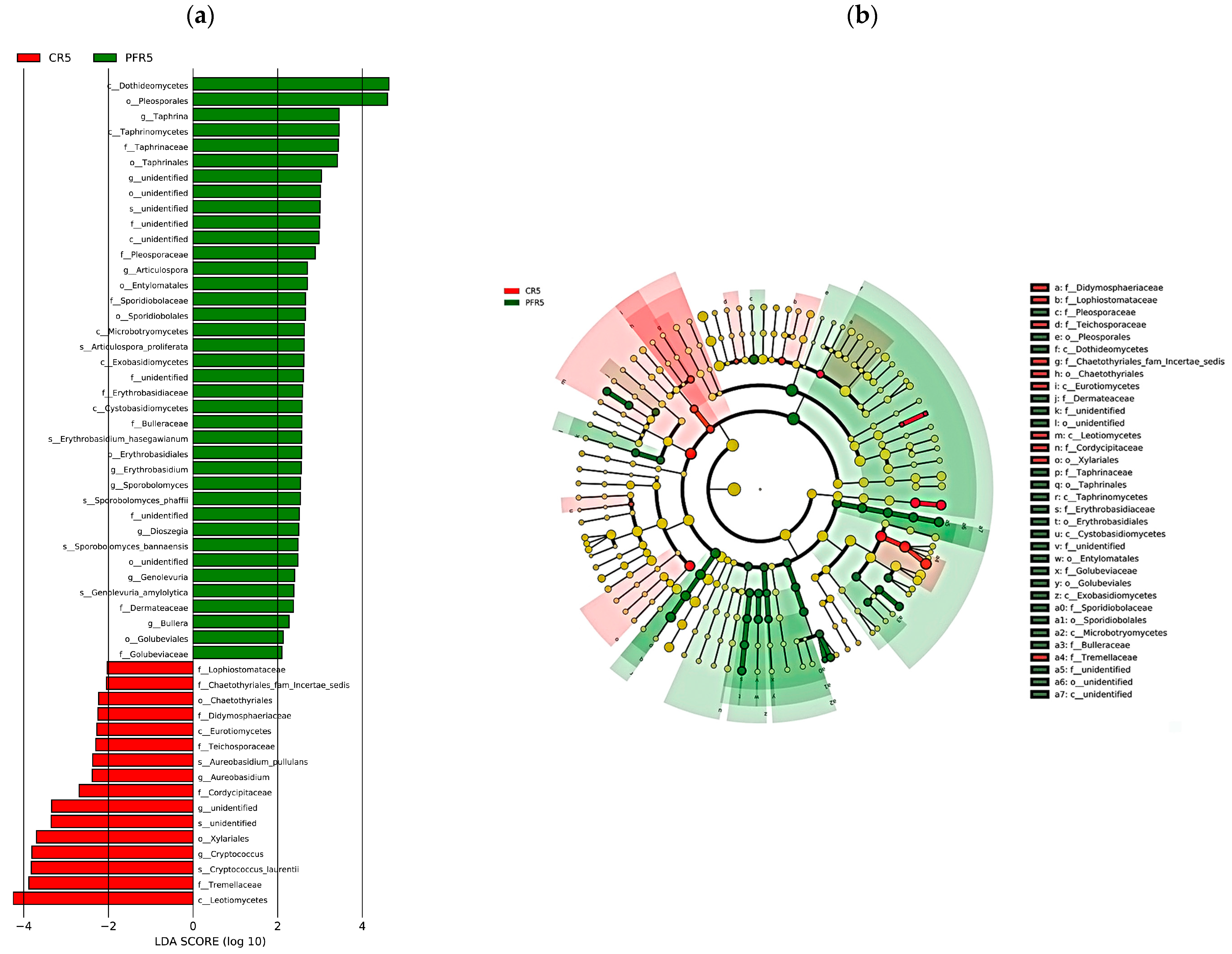
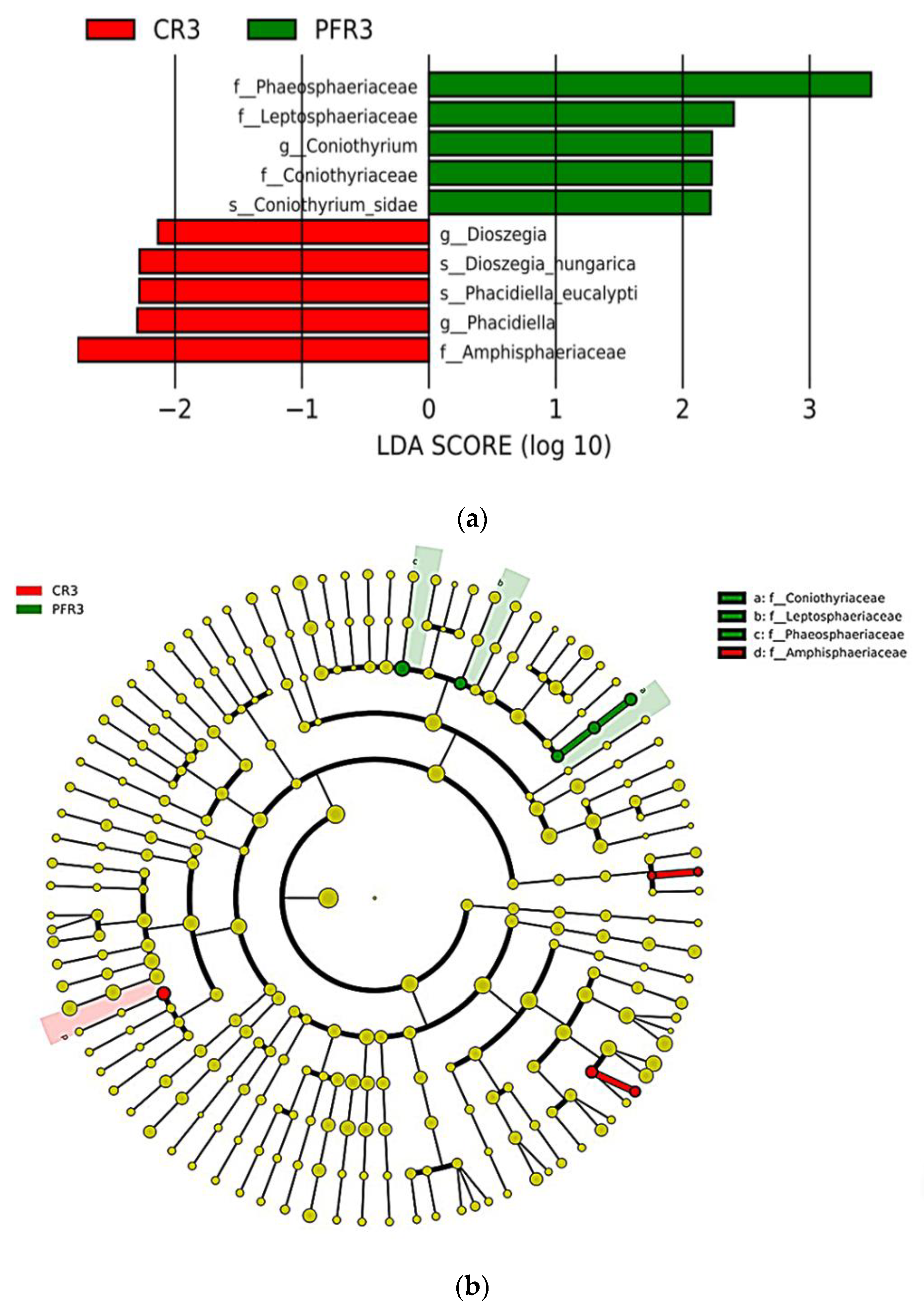
3.5. Fungal Traits of Bagged Apple Microbiome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forte, A.V.; Ignatov, A.N.; Ponomarenko, V.V.; Dorokhov, D.B.; Savelyev, N.I. Phylogeny of the Malus (apple tree) species, inferred from the morphological traits and molecular DNA analysis. Russ. J. Genet. 2002, 38, 1150–1160. [Google Scholar] [CrossRef]
- U. S. Departement of Agriculture. Index of Plant Diseases in the United States; Agriculture handbook no. 165; U. S. Departement of Agriculture: Washington, DC, USA, 1960.
- Wang, X.; Shi, C.-M.; Gleason, M.L.; Huang, L. Fungal species associated with apple Valsa canker in East Asia. Phytopathol. Res. 2020, 2, 35. [Google Scholar] [CrossRef]
- Sharma, R.R.; Reddy, S.V.R.; Jhalegar, M.J. Pre-harvest fruit bagging: A useful approach for plant protection and improved post-harvest fruit quality—A review. J. Hortic. Sci. Biotechnol. 2014, 89, 101–113. [Google Scholar] [CrossRef]
- Sharma, R.R. Fruit Production: Problems and Solutions; International Book Distributing Company: New Delhi, India, 2006. [Google Scholar]
- Kitagawa, H.; Manabe, K.; Esguerra, E.B. Bagging of fruit on the tree to control disease. Acta Hortic. 1992, 321, 871–875. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, S.K.; Lim, S.C.; Lee, C.H.; Youn, C.K.; Kim, H.H.; Choi, K.S. Effects of bagging material on coloration, maturity, and quality of peach fruits. J. Korean Soc. Hortic. Sci. 2000, 41, 395–400. [Google Scholar]
- Jia, H.-J.; Araki, A.; Okamoto, G. Influence of fruit bagging on aroma volatiles and skin coloration of ’Hakuho’ peach (Prunus persica Batsch). Postharvest Biol. Technol. 2005, 35, 61–68. [Google Scholar] [CrossRef]
- Faci, J.M.; Medina, E.T.; Martínez-Cob, A.; Alonso, J.M. Fruit yield and quality response of a late season peach orchard to different irrigation regimes in a semi-arid environment. Agric. Water Manag. 2014, 143, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.R.; Pal, R.K.; Asrey, R.; Sagar, V.R.; Dhiman, M.R.; Rana, M.R. Pre-harvest fruit bagging influences fruit color and quality of apple cv. Delicious. Agric. Sci. 2013, 4, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Bai, S.; Tuan, P.A.; Saito, T.; Honda, C.; Hatsuyama, Y.; Ito, A.; Moriguchi, T. Epigenetic regulation of MdMYB1 is associated with paper bagging-induced red pigmentation of apples. Planta 2016, 244, 573–586. [Google Scholar] [CrossRef]
- Xu, G.; Nie, J.; Wu, Y.; Yan, Z.; Ye, M. The effects of fruit bagging on residue behavior and dietary risk for four pesticides in apple. Sci. Rep. 2018, 8, 14348. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Chang, Y.; Yan, Z.; Li, X. Effects of bagging on the quality of pear fruit and pesticide residues. Acta Hortic. 2008, 772, 315–318. [Google Scholar] [CrossRef]
- Bentley, W.J.; Viveros, M. Brown-bagging Granny Smith apples on trees stops codling moth damage. Calif. Agric. 1992, 46, 30–32. [Google Scholar] [CrossRef]
- Wang, J.Y.; Wang, S.M.; Gao, H.J. Review on research progress on the control of apple diseases and insect pests by bagging. Chin. Agric. Bull. 2006, 22, 423–426. (In Chinese) [Google Scholar]
- Hofman, P.J.; Smith, L.G.; Joyce, D.C.; Johnson, G.I.; Meiburg, G.F. Bagging of mango (Mangifera indica cv. ’Keitt’) fruit influences fruit quality and mineral composition. Postharvest Biol. Technol. 1997, 12, 83–91. [Google Scholar] [CrossRef]
- Joyce, D.C.; Beasley, D.R.; Shorter, A.J. Effect of preharvest bagging on fruit calcium levels, and storage and ripening characteristics of ‘Sensation’ mangoes. Aust. J. Exp. Agric. 1997, 37, 383–389. [Google Scholar]
- Dai, P.; Liang, X.; Wang, Y.; Gleason, M.L.; Zhang, R.; Sun, G. High humidity and age-dependent fruit susceptibility promote development of Trichothecium black spot on apple. Plant Dis. 2019, 103, 259–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granado, J.; Thürig, B.; Kieffer, E.; Petrini, L.; Fließbach, A.; Tamm, L.; Weibel, F.P.; Wyss, G.S. Culturable fungi of stored ‘golden delicious’ apple fruits: A one-season comparison study of organic and integrated production systems in Switzerland. Microb. Ecol. 2008, 56, 720–732. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, A.; Wisniewski, M.; Droby, S.; Schena, L. Spatial and compositional variation in the fungal communities of organic and conventionally grown apple fruit at the consumer point-of-purchase. Hortic. Res. 2016, 3, 16047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Abdelfattah, A.; Norelli, J.; Burchard, E.; Schena, L.; Droby, S.; Wisniewski, M. Apple endophytic microbiota of different rootstock/scion combinations suggests a genotype-specific influence. Microbiome 2018, 6, 18. [Google Scholar] [CrossRef]
- Vepštaitė-Monstavičė, I.; Lukša, J.; Stanevičienė, R.; Strazdaitė-Žielienė, Ž.; Yurchenko, V.; Serva, S.; Servienė, E. Distribution of apple and blackcurrant microbiota in Lithuania and the Czech Republic. Microbiol. Res. 2018, 206, 1–8. [Google Scholar] [CrossRef]
- Shen, Y.; Nie, J.; Li, Z.; Li, H.; Wu, Y.; Dong, Y.; Zhang, J. Differentiated surface fungal communities at point of harvest on apple fruits from rural and peri-urban orchards. Sci. Rep. 2018, 8, 2165. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D. Endophyte—The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Mukhtar, I.; Mushtaq, S.; Ali, A.; Khokhar, I. Epiphytic and endophytic phyllosphere microflora of Cassytha Filiformis L. and its hosts. Ecoprint Int. J. Ecol. 2010, 17, 1–8. [Google Scholar] [CrossRef]
- Lata, R.; Chowdhury, S.; Gond, S.K.; White, J.F. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 2018, 66, 268–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Viret, O.; Keller, M.; Jaudzems, V.G.; Cole, F.M. Botrytis cinerea infection of grape flowers: Light and electron microscopical studies of infection sites. Phytopathology 2004, 94, 850–857. [Google Scholar] [CrossRef] [Green Version]
- Andrews, J.H.; Kenerley, C.M. Microbial populations associated with buds and young leaves of apple. Can. J. Bot. 1980, 58, 847–855. [Google Scholar] [CrossRef]
- Chand-Goyal, T.; Spotts, R.A. Enumeration of bacterial and yeast colonists of apple fruits and identification of epiphytic yeasts on pear fruits in the Pacific Northwest United States. Microbiol. Res. 1996, 151, 427–432. [Google Scholar] [CrossRef]
- Serdani, M.; Crous, P.W.; Holz, G.; Petrini, O. Endophytic fungi asso- ciated with core rot of apples in South Africa, with specific reference to Alternaria species. Sydowia 1998, 50, 257–271. [Google Scholar]
- Woody, S.T.; Spear, R.N.; Nordheim, E.V.; Ives, A.R.; Andrews, J.H. Single-leaf resolution of the temporal population dynamics of Aureobasidium pullulans on apple leaves. Appl. Environ. Microbiol. 2003, 69, 4892–4900. [Google Scholar] [CrossRef] [Green Version]
- Manici, L.M.; Kelderer, M.; Franke-whittle, I.H.; Rühmer, T.; Baab, G.; Nicoletti, F.; Caputo, F.; Topp, A.; Insam, H.; Naef, A. Relationship between root-endophytic microbial communities and replant disease in specialized apple growing areas in Europe. Appl. Soil Ecol. 2013, 72, 207–214. [Google Scholar] [CrossRef]
- Kellenberger, E. Exploring the unknown: The silent revolution of microbiology. EMBO Rep. 2001, 2, 5–7. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Purahong, W.; Wubet, T.; Hyde, K.D.; Zhang, W.; Xu, H.; Zhang, G.; Fu, C.; Liu, M.; Xing, Q.; et al. Direct comparison of culture-dependent and culture-independent molecular approaches reveal the diversity of fungal endophytic communities in stems of grapevine (Vitis vinifera). Fungal Divers. 2018, 90, 85–107. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Purahong, W.; Zhang, W.; Wubet, T.; Li, X.H.; Liu, M.; Zhao, W.; Hyde, K.D.; Liu, J.H.; Yan, J. Biodiversity of fungi on Vitis vinifera L. revealed by traditional and high-resolution culture-independent approaches. Fungal Divers. 2018, 90, 1–84. [Google Scholar] [CrossRef] [Green Version]
- Buée, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S.; Martin, F. 454 Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, A.; Whitehead, S.R.; Macarisin, D.; Liu, J.; Burchard, E.; Freilich, S.; Dardick, C.; Droby, S.; Wisniewski, M. Effect of washing, waxing and low-temperature storage on the postharvest microbiome of apple. Microorganisms 2020, 8, 944. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, A.; Wisniewski, M.; Li Destri Nicosia, M.G.; Destri, L.; Cacciola, S.O.; Schena, L. Metagenomic analysis of fungal diversity on strawberry plants and the effect of management practices on the fungal community structure of aerial organs. PLoS ONE 2016, 11, e0160470. [Google Scholar] [CrossRef]
- Kou, R.; Lam, H.; Duan, H.; Ye, L.; Jongkam, N.; Chen, W.; Zhang, S.; Li, S. Benefits and challenges with applying unique molecular identifiers in next generation sequencing to detect low frequency mutations. PLoS ONE 2016, 11, e0146638. [Google Scholar] [CrossRef] [Green Version]
- Canzoniero, J.V.; Cravero, K.; Park, B.H. The impact of collisions on the ability to detect rare mutant alleles using barcode-type next-generation sequencing techniques. Cancer Inform. 2017, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kinde, I.; Wu, J.; Papadopoulos, N.; Kinzler, K.W.; Vogelstein, B. Detection and quantification of rare mutations with massively parallel sequencing. Proc. Natl. Acad. Sci. USA 2011, 108, 9530–9535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duong, L.M.; Jeewon, R.; Lumyong, S.; Hyde, K.D. DGGE coupled with ribosomal DNA gene phylogenies reveal uncharacterized fungal phylotypes. Fungal Divers. 2006, 23, 121–138. [Google Scholar]
- McKinnon, A.C. Plant tissue preparation for the detection of an endophytic fungus in planta. Methods Mol. Biol. 2016, 1477, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Tejesvi, M.V.; Picart, P.; Kajula, M.; Hautajärvi, H.; Ruddock, L.; Kristensen, H.H.; Tossi, A.; Sahl, H.G.; Ek, S.; Mattila, S.; et al. Identification of antibacterial peptides from endophytic microbiome. Appl. Microbiol. Biotechnol. 2016, 100, 9283–9293. [Google Scholar] [CrossRef]
- Shade, A.; McManus, P.S.; Handelsman, J. Unexpected diversity during community succession in the apple flower microbiome. mBio 2013, 4, e00602-12. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.L. Evaluation of fruit bagging as a pest management option for direct pests of apple. Insects 2018, 9, 178. [Google Scholar] [CrossRef] [Green Version]
- Grasswitz, T.R.; Fimbres, O. Efficacy of a physical method for control of direct pests of apples and peaches. J. Appl. Entomol. 2013, 137, 790–800. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus (Madison) 1990, 12, 13–15. [Google Scholar]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identifification of fungi-recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 623–656. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity, 2nd ed.; Blackwell Science Ltd: Oxford, UK, 2004. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial (Plymouth Routines in Multivariate Ecological Research); PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Kim, B.-R.; Shin, J.; Guevarra, R.B.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Lee, J.-H.; Kim, H.B.; Isaacson, R.E. Deciphering diversity indices for a better understanding of microbial communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [Green Version]
- Kirk, P.M.; Ainsworth, G.C.; Grossbritannien, P.; Bisby, G.R. Ainsworth and Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Lutzoni, F.; Kauff, F.; Cox, C.J.; McLaughlin, D.; Celio, G.; Dentinger, C.; Padamsee, M.; Hibbett, D.; James, T.Y.; Baloch, E.; et al. Assembling the fungal tree of life: Progress, classification, and evolution of subcellular traits. Am. J. Bot. 2004, 91, 1446–1480. [Google Scholar] [CrossRef]
- Chen, Y.L.; Xu, T.L.; Veresoglou, S.D.; Hu, H.W.; Hao, Z.P.; Hu, Y.J.; Liu, L.; Deng, Y.; Rillig, M.C.; Chen, B.D. Plant diversity represents the prevalent determinant of soil fungal community structure across temperate grasslands in northern China. Soil Biol. Biochem. 2017, 110, 12–21. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Arafat, Y.; Lin, W. Studies on fungal communities and functional guilds shift in tea continuous cropping soils by high-throughput sequencing. Ann. Microbiol. 2020, 70, 7. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Liu, W.; Zhu, C.; Luo, G.; Kong, Y.; Ling, N.; Wang, M.; Dai, J.; Shen, Q.; Guo, S. Bacterial rather than fungal community composition is associated with microbial activities and nutrient-use efficiencies in a paddy soil with short-term organic amendments. Plant Soil 2018, 424, 335–349. [Google Scholar] [CrossRef]
- Michailides, T.J.; Morgan, D.P.; Mitcham, E.; Crisosto, C. Occurrence of mouldy core and core rot of Fuji apple in California. KAC Plant Prot. Q. 1994, 3, 4–6. [Google Scholar]
- Gao, L.L.; Zhang, Q.; Sun, X.Y.; Jiang, L.; Zhang, R.; Sun, G.Y.; Zha, Y.L.; Biggs, A.R. Etiology of moldy core, core browning, and core rot of Fuji apple in China. Plant Dis. 2013, 97, 510–516. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.Z.; Sun, G.Y.; Gao, B.W.; Li, C.Y.; Zhang, P.L.; Lei, X.L. Studies on the identification of pathogen and the biological characteristics of black-dot disease of bagged apple. Acta Agric. Boreali-Occident. Sinia 2005, 14, 18–21. (In Chinese) [Google Scholar]
- Hou, Y.M.; Zhang, X.; Zhang, N.N.; Naklumpa, W.; Zhao, W.Y.; Liang, X.F.; Zhang, R.; Sun, G.Y.; Gleason, M.L. Genera Acremonium and Sarocladium cause brown spot on bagged apple fruit in China. Plant Dis. 2019, 103, 1889–1901. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Huang, Y.; Liu, J.; Zhang, Q.; Wang, C. First report of shoot dieback on apple caused by Diaporthe nobilis in China. Plant Dis. 2020, 104, 1–2. [Google Scholar] [CrossRef]
- Gutierrez, M.; Teoduloz, C.; Rodriguez, J.; Lolas, M.; Schmeda-Hirschmann, G. Bioactive metabolites from the fungus Nectria galligena, the main apple canker agent in Chile. J. Agric. Food Chem. 2005, 53, 7701–7708. [Google Scholar] [CrossRef]
- Elfar, K.; Zoffoli, J.P.; Latorre, B.A. Identification and characterization of Alternaria species associated with moldy core of apple in Chile. Plant Dis. 2018, 102, 2158–2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munir, M.; Amsden, B.; Dixon, E.; Vaillancourt, L.; Ward Gauthier, N.A. Characterization of Colletotrichum species causing bitter rot of apple in Kentucky orchards. Plant Dis. 2016, 100, 2194–2203. [Google Scholar] [CrossRef] [Green Version]
- Martins, N.; Ferreira, I.C.; Barros, L.; Silva, S.; Henriques, M. Candidiasis: Predisposing factors, prevention, diagnosis and alternative treatment. Mycopathologia 2014, 177, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Golubev, W.; Ikeda, R.; Shinoda, T.; Nakase, T. Antifungal activity of Bullera alba (Hanna) derx. Mycoscience 1997, 38, 25–29. [Google Scholar] [CrossRef]
- Teixidó, N.; Usall, J.; Gutierrez, O.; Viñas, I. Effect of the antagonist Candida sake on apple surface microflora during cold and ambient (shelf life) storage. Eur. J. Plant Pathol. 1998, 104, 387–398. [Google Scholar] [CrossRef]
- Hoppe, B.; Purahong, W.; Wubet, T.; Kahl, T.; Bauhus, J.; Arnstadt, T.; Hofrichter, M.; Buscot, F.; Krüger, D. Linking molecular deadwood-inhabiting fungal diversity and community dynamics to ecosystem functions and processes in Central European forests. Fungal Divers. 2016, 77, 367–379. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, W.P. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [Green Version]
- Filonow, A.B.; Vishniac, H.S.; Anderson, J.A.; Janisiewicz, W.J. Biological control of Botrytis cinerea in apple by yeasts from various habitats and their putative mechanisms of antagonism. Biol. Control. 1996, 7, 212–220. [Google Scholar] [CrossRef]
- Raspor, P.; Miklič-Milek, D.; Avbelj, M.; Čadež, N. Biocontrol of grey mould disease on grape caused by Botrytis cinerea with autochthonous wine yeasts. Food Technol. Biotechnol. 2010, 48, 336–343. [Google Scholar]
- Gramisci, B.R.; Lutz, M.C.; Lopes, C.A.; Sangorrín, M.P. Enhancing the efficacy of yeast biocontrol agents against postharvest pathogens through nutrient profiling and the use of other additives. Biol. Control. 2018, 121, 151–158. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Review: Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef]
- Havenga, M.; Gatsi, G.M.; Halleen, F.; Spies, C.F.J.; van der Merwe, R.; Mostert, L. Canker and wood rot pathogens present in young apple trees and propagation material in the Western Cape of South Africa. Plant Dis. 2019, 103, 3129–3141. [Google Scholar] [CrossRef]
- Brooks, F.T. On the occurrence of Phacidiella discolor (Mout. & Sacc.) Potebnia in England. Trans. Br. Mycol. Soc. 1928, 13, 75–81. [Google Scholar] [CrossRef]
- De Hoog, G.S. Evolution of black yeasts: Possible adaptation to the human host. Antonie Van Leeuwenhoek 1993, 63, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Voigt, O.; Knabe, N.; Nitsche, S.; Erdmann, E.A.; Schumacher, J.; Gorbushina, A.A. An advanced genetic toolkit for exploring the biology of the rock-inhabiting black fungus Knufia Petricola. Sci. Rep. 2020, 10, 22021. [Google Scholar] [CrossRef]
- Lin, Y.Y.; Shiau, S.; Fang, C.T. Risk factors for invasive Cryptococcus neoformans diseases: A case-control study. PLoS ONE 2015, 10, e0119090. [Google Scholar] [CrossRef] [Green Version]
- Eisenman, H.C.; Casadevall, A.; McClelland, E.E. New insights on the pathogenesis of invasive Cryptococcus neoformans infection. Curr. Infect. Dis. Rep. 2007, 9, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Barnes, E.H. Bacteria on leaf surfaces and in intercellular leaf spaces. Science 1965, 147, 1151–1152. [Google Scholar] [CrossRef]
- Henis, Y.; Bashan, Y. Epiphytic survival of bacterial leaf pathogens. In Microbiology of the Phyllosphere; Fokkema, N., van den Heuvel, J., Eds.; Cambridge Univ. Press: New York, NY, USA, 1986; Volume 3, pp. 252–268. [Google Scholar]
- Talley, S.M.; Coley, P.D.; Kursar, T.A. The effects of weather on fungal abundance and richness among 25 communities in the Intermountain West. BMC Ecol. 2002, 2, 7. [Google Scholar] [CrossRef]
- Willis, A.D. Rarefaction, alpha diversity, and statistics. Front. Microbiol. 2019, 10, 2407. [Google Scholar] [CrossRef] [Green Version]
- Li, E.M.; Shi, D.C.; Xu, Y.H.; Chen, F.; Zhai, H. Changing characteristics of temperature and humidity in different type bags for bagging apple and their effects on fruit appearance quality. Chin. J. Appl. Ecol. 2008, 19, 208–212. [Google Scholar]
- Andrews, J.H.; Kenerley, C.M. The effects of a pesticide program on non-target epiphytic microbial populations of apple leaves. Can. J. Microbiol. 1978, 24, 1058–1072. [Google Scholar] [CrossRef]
- Zaller, J.G.; Brühl, C.A. Editorial: Non-target effects of pesticides on organisms inhabiting agroecosystems. Front. Environ. Sci. 2019, 7, 75. [Google Scholar] [CrossRef] [Green Version]
- Leff, J.W.; Fierer, N. Bacterial communities associated with the surfaces of fresh fruits and vegetables. PLoS ONE 2013, 8, e59310. [Google Scholar] [CrossRef] [Green Version]
- Sugahara, H.; Kondo, T.; Okada, M.; Ikeda, Y.; Kaida, K.; Fudou, R.; Mizuno, T.; Sakagami, Y. Articulospora sp. produces Art1, an inhibitor of bacterial histidine kinase. Biosci. Biotechnol. Biochem. 2008, 72, 2521–2525. [Google Scholar] [CrossRef] [Green Version]
- Renker, C.; Blanke, V.; Börstler, B.; Heinrichs, J.; Buscot, F. Diversity of Cryptococcus and Dioszegia yeasts (Basidiomycota) inhabiting arbuscular mycorrhizal roots or spores. FEMS Yeast Res. 2004, 4, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Janisiewicz, W.J.; Kurtzman, C.P.; Buyer, J.S. Yeasts associated with nectarines and their potential for biological control of brown rot. Yeast 2010, 27, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Khosla, K.; Bhardwaj, S.S. Control of peach leaf curl by pre-bloom and post infection applications of fungicides. Plant Dis. Res. 2010, 25, 66–68. [Google Scholar]
- Cissé, O.H.; Almeida, J.M.G.C.F.; Fonseca, Á.; Kumar, A.A.; Salojärvi, J.; Overmyer, K.; Hauser, P.M.; Pagni, M. Genome sequencing of the plant pathogen Taphrina deformans, the causal agent of peach leaf curl. mBio 2013, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Köhl, J. Biological Control of Botrytis spp. by Ulocladium Atrum through Competitive Colonisation of Necrotic Plant Tissues; Plant Research International. B.V.: Wageningen, The Netherlands, 2004. [Google Scholar]
- Li, Z.F.; Wang, L.F.; Feng, Z.L.; Zhao, L.H.; Shi, Y.Q.; Zhu, H.Q. Diversity of endophytic fungi from different Verticillium-wilt-resistant Gossypium hirsutum and evaluation of antifungal activity against Verticillium dahliae in vitro. J. Microbiol. Biotechnol. 2014, 24, 1149–1161. [Google Scholar] [CrossRef] [Green Version]
- Pandey, R.R.; Arora, D.K.; Dubey, R.C. Antagonistic interactions between fungal pathogens and phylloplane fungi of guava. Mycopathologia 1993, 124, 31–39. [Google Scholar] [CrossRef]
- Elmer, P.A.G.; Reglinski, T. Biosuppression of Botrytis cinerea in grapes. Plant Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Bosshard, E.; Chüepp, H.; Siegfried, W. Concepts and methods in biological control of diseases in apple orchards. EPPO Bull. 1987, 17, 655–663. [Google Scholar] [CrossRef]
- Alexander, B.J.R.; Stewart, A. Glasshouse screening for biological control agents of Phytophthora cactorum on apple (Malus domestica). N. Z. J. Crop. Hortic. Sci. 2001, 29, 159–169. [Google Scholar] [CrossRef]
- Schena, L.; Ippolito, A.; Zahavi, T.; Cohen, L.; Nigro, F.; Droby, S. Genetic diversity and biocontrol activity of Aureobasidium pullulans isolates against postharvest rots. Postharvest Biol. Technol. 1999, 17, 189–199. [Google Scholar] [CrossRef]
- Ippolito, A.; El Ghaouth, A.; Wilson, C.L.; Wisniewski, M. Control of postharvest decay of apple fruit by Aureobasidium pullulans and induction of defense responses. Postharvest Biol. Technol. 2000, 19, 265–272. [Google Scholar] [CrossRef]
- Kheireddine, A.; Essghaier, B.; Hedi, A.; Dhieb, C.; Sadfi-Zouaoui, N. New epiphytic yeasts able to reduce grey mold disease on apples. Plant Prot. Sci. 2018, 54, 248–257. [Google Scholar]
- Meador, D.B.; Taylor, B.H. Effect of early season foliar sprays of GA4+7 on russeting and return bloom of Golden Delicious apple. HortScience 1987, 22, 412–415. [Google Scholar]
- Matteson Heidenreich, M.C.; Corral-Garcia, M.R.; Momol, E.A.; Burr, T.J. Russet of apple fruit caused by Aureobasidium pullulans and Rhodotorula glutinis. Plant Dis. 1997, 81, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, A.; Grimm, E.; Knoche, M. Late-season surface water induces skin spot in apple. HortScience 2014, 49, 1324–1327. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Straube, J.; Khanal, B.P.; Knoche, M.; Debener, T. Russeting in apple is initiated after exposure to moisture ends—I. histological evidence. Plants 2020, 9, 1293. [Google Scholar] [CrossRef] [PubMed]
- Tukey, L.D. Observations on the russeting of apples growing in plastic bags. Proc. Am. Soc. Hortic. Sci. 1969, 74, 30–39. [Google Scholar]
- Dimakopoulou, M.; Tjamos, S.E.; Antoniou, P.P.; Pietri, A.; Battilani, P.; Avramidis, N.; Markakis, E.A.; Tjamos, E.C. Phyllosphere grapevine yeast Aureobasidium pullulans reduces Aspergillus carbonarius ( sour rot ) incidence in wine-producing vineyards in Greece. Biol. Control. 2008, 46, 158–165. [Google Scholar] [CrossRef]
- Hashem, M.; Alamri, S.A.; Hesham, A.E.-L.; Al-Qahtani, F.M.H.; Kilany, M. Biocontrol of apple blue mould by new yeast strains: Cryptococcus albidus KKUY0017 and Wickerhamomyces anomalus KKUY0051 and their mode of action. Biocontrol. Sci. Technol. 2014, 24, 1137–1152. [Google Scholar] [CrossRef]
- Moller, L.; Kessler, K.D.; Steyn, A.; Valentine, A.J.; Botha, A. The role of Cryptococcus laurentii and mycorrhizal fungi in the nutritional physiology of Lupinus angustifolius L. hosting N2-fixing nodules. Plant Soil 2016, 409, 345–360. [Google Scholar] [CrossRef]
- Verkley, G.J.M.; Da Silva, M.; Wicklow, D.T.; Crous, P.W. Paraconiothyrium, a new genus to accommodate the mycoparasite Coniothyrium minitans, anamorphs of Paraphaeosphaeria, and four new species. Stud. Mycol. 2004, 50, 323–335. [Google Scholar]
- Damm, U.; Verkley, G.J.M.; Crous, P.W.; Fourie, P.H.; Haegi, A.; Riccioni, L. Novel Paraconiothyrium species on stone fruit trees and other woody hosts. Persoonia 2008, 20, 9–17. [Google Scholar] [CrossRef]
- Gama, D.D.S.; Santos, Í.A.F.M.; De Abreu, L.M.; De Medeiros, F.H.V.; Duarte, W.F.; Cardoso, P.G. Endophytic fungi from Brachiaria grasses in Brazil and preliminary screening of Sclerotinia sclerotiorum antagonists. Sci. Agric. 2020, 77, e20180210. [Google Scholar] [CrossRef]
- Khokhar, I.; Bajwa, R. Prevalence of post-harvest rot of fruits and vegetables by Penicillium species. Int. J. Adv. Res. Biol. Sci. 2014, 1, 14–19. [Google Scholar]
- Garibaldi, A.; Bertetti, D.; Ortu, G.; Gullino, M.L. A leaf spot caused by Stagonosporopsis trachelii on Campanula medium in Italy. J. Plant Pathol. 2015, 97, 546. [Google Scholar]
- Costamilan, L.M. Variability of the wheat powdery mildew pathogen Blumeria graminis f. sp. tritici in the 2003 crop season. Fitopatol. Bras. 2005, 30, 420–422. [Google Scholar] [CrossRef]
- O’Gorman, D.T.; Sholberg, P.L.; Stokes, S.C.; Ginns, J. DNA sequence analysis of herbarium specimens facilitates the revival of Botrytis mali, a postharvest pathogen of apple. Mycologia 2008, 100, 227–235. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Kristiansson, E.; Ryberg, M.; Hallenberg, N.; Larsson, K.H. Intraspecific ITS variability in the Kingdom Fungi as expressed in the international sequence databases and its implications for molecular species identification. Evol. Bioinform. 2008, 2008, 193–201. [Google Scholar] [CrossRef]
- Gezahgne, A.; Cortinas, M.N.; Wingfield, M.J.; Roux, J. Characterisation of the Coniothyrium stem canker pathogen on Eucalyptus camaldulensis in Ethiopia. Australas. Plant Pathol. 2005, 34, 85–90. [Google Scholar] [CrossRef]
- Ali, S.; Abbasi, P.; Rehman, S.; Ellouze, W. First Report of Moldy Core of Sweet Tango Apples from New Zealand Caused by Alternaria arborescens. Plant Dis. 2021. [Google Scholar] [CrossRef]
- De Vrije, T.; Antoine, N.; Buitelaar, R.M.; Bruckner, S.; Dissevelt, M.; Durand, A.; Gerlagh, M.; Jones, E.E.; Lüth, P.; Oostra, J.; et al. The fungal biocontrol agent Coniothyrium minitans: Production by solid-state fermentation, application and marketing. Appl. Microbiol. Biotechnol. 2001, 56, 58–68. [Google Scholar] [CrossRef]
- Grove, G.G.; Eastwell, K.C.; Jones, A.L.; Sutton, T.B. Diseases of apple. In Apples: Botany, Production and Uses; Ferree, D.C., Warrington, I.J., Eds.; CAB International: Wallingford, CT, USA, 2003; pp. 459–488. [Google Scholar]
- Pinto, C.; Pinho, D.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Unravelling the diversity of grapevine microbiome. PLoS ONE 2014, 9, e85622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Ichinose, H. Cytochrome P450 of wood-rotting basidiomycetes and biotechnological applications. Biotechnol. Appl. Biochem. 2013, 60, 71–81. [Google Scholar] [CrossRef]
- Trdá, L.; Barešová, M.; Šašek, V.; Nováková, M.; Zahajská, L.; Dobrev, P.I.; Motyka, V.; Burketová, L. Cytokinin metabolism of pathogenic fungus Leptosphaeria maculans involves isopentenyltransferase, adenosine kinase and cytokinin oxidase/dehydrogenase. Front. Microbiol. 2017, 8, 1374. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L.; Kannangara, S.D.; Promputtha, I. Fungi vs. Fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef]
- Farzaneh, V.; Carvalho, I.S. Modelling of microwave assisted extraction (MAE) of anthocyanins (TMA). J. Appl. Res. Med. Aromat. Plants 2017, 6, 92–100. [Google Scholar] [CrossRef]
- Farzaneh, V.; Gominho, J.; Pereira, H.; Carvalho, I.S. Screening of the antioxidant and enzyme inhibition potentials of portuguese Pimpinella anisum L. Seeds by GC-MS. Food Anal. Methods 2018, 11, 2645–2656. [Google Scholar] [CrossRef]
- Zola, F.G.; Rodrigues, A.C.; Oliveira, B.D.; Sacramento, N.T.B.; Taylor, J.G.; Pinto, U.M.; Bertoldi, M.C. Mineral and centesimal contents, antioxidant activity and antimicrobial action of phenolic compounds from Eugenia brasiliensis Lam. pulp. Food Sci. Technol. 2019, 39, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E.; Kutty-Amma, S.; Allan, A.C. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007, 49, 414–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzaneh, V.; Carvalho, I.S. A review of the health benefit potentials of herbal plant infusions and their mechanism of actions. Ind. Crops Prod. 2015, 65, 247–258. [Google Scholar] [CrossRef]
- Schaefer, H.M.; Rentzsch, M.; Breuer, M. Anthocyanins reduce fungal growth in fruits. Nat. Prod. Commun. 2008, 3, 1267–1272. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.C.; Yu, M.; Liu, F.; Qu, J.Z.; Pan, X.X.; Zhang, H.B.; Yang, M.Z. Diversity distributions and the anthocyanin associations of fungal endophytes in different colored grapevine leaves. J. Plant Biol. 2020, 63, 107–116. [Google Scholar] [CrossRef]
- Diskin, S.; Feygenberg, O.; Maurer, D.; Droby, S.; Prusky, D.; Alkan, N. Microbiome alterations are correlated with occurrence of postharvest stem-end rot in mango fruit. Phytobiomes J. 2017, 1, 117–127. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Application Date in 2019 | Associated Pesticides |
|---|---|
| 10 May | 30% pyraclostrobin • tebuconazole + 23% lambda-cyhalothrin |
| 25 May | 30% pyraclostrobin • tebuconazole + 12% deltamethrin • thiamethoxam |
| 20 June | 30% hexaconazole + 70% propsenzine + 2.3% emamectin benzoate + 5% potassium dihydrogen phosphate |
| 10 July | 43% tebuconazole + 80% mancozeb + 23% lambda cyhalothrin + 5% potassium dihydrogen phosphate |
| 5 August | 25% propiconazole + thiophanate methyl + 12% deltamethrin • thiamethoxam + 5% potassium dihydrogen phosphate |
| 20 August | 27% thiophenone • tebuconazole + 2.3% emamectin benzoate |
| CR3 | CR5 | PFR3 | PFR5 | |
|---|---|---|---|---|
| CR3 | 2100 ± 427 | 0.191 | 0.603 | 0.05 |
| CR5 | - | 2510 ± 149 | 0.13 | 0.0005 |
| PFR3 | - | - | 1870 ± 565 | 0.223 |
| PFR5 | - | - | - | 1390 ± 118 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khwantongyim, P.; Wansee, S.; Lu, X.; Zhang, W.; Sun, G. Variations in the Community Structure of Fungal Microbiota Associated with Apple Fruit Shaped by Fruit Bagging-Based Practice. J. Fungi 2021, 7, 764. https://doi.org/10.3390/jof7090764
Khwantongyim P, Wansee S, Lu X, Zhang W, Sun G. Variations in the Community Structure of Fungal Microbiota Associated with Apple Fruit Shaped by Fruit Bagging-Based Practice. Journal of Fungi. 2021; 7(9):764. https://doi.org/10.3390/jof7090764
Chicago/Turabian StyleKhwantongyim, Punda, Somying Wansee, Xi Lu, Wei Zhang, and Guangyu Sun. 2021. "Variations in the Community Structure of Fungal Microbiota Associated with Apple Fruit Shaped by Fruit Bagging-Based Practice" Journal of Fungi 7, no. 9: 764. https://doi.org/10.3390/jof7090764