
Five Novel Taxa from Freshwater Habitats and New Taxonomic Insights of Pleurotheciales and Savoryellomycetidae

 , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Herbarium Material, Isolation and Morphology
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Molecular Phylogenetic Analyses
2.3.1. Sequence Selection and Phylogenetic Analyses Construction
2.3.2. Maximum Likelihood Analyses
2.3.3. Bayesian Inference Analyses
3. Results
3.1. Phylogenetic Analyses
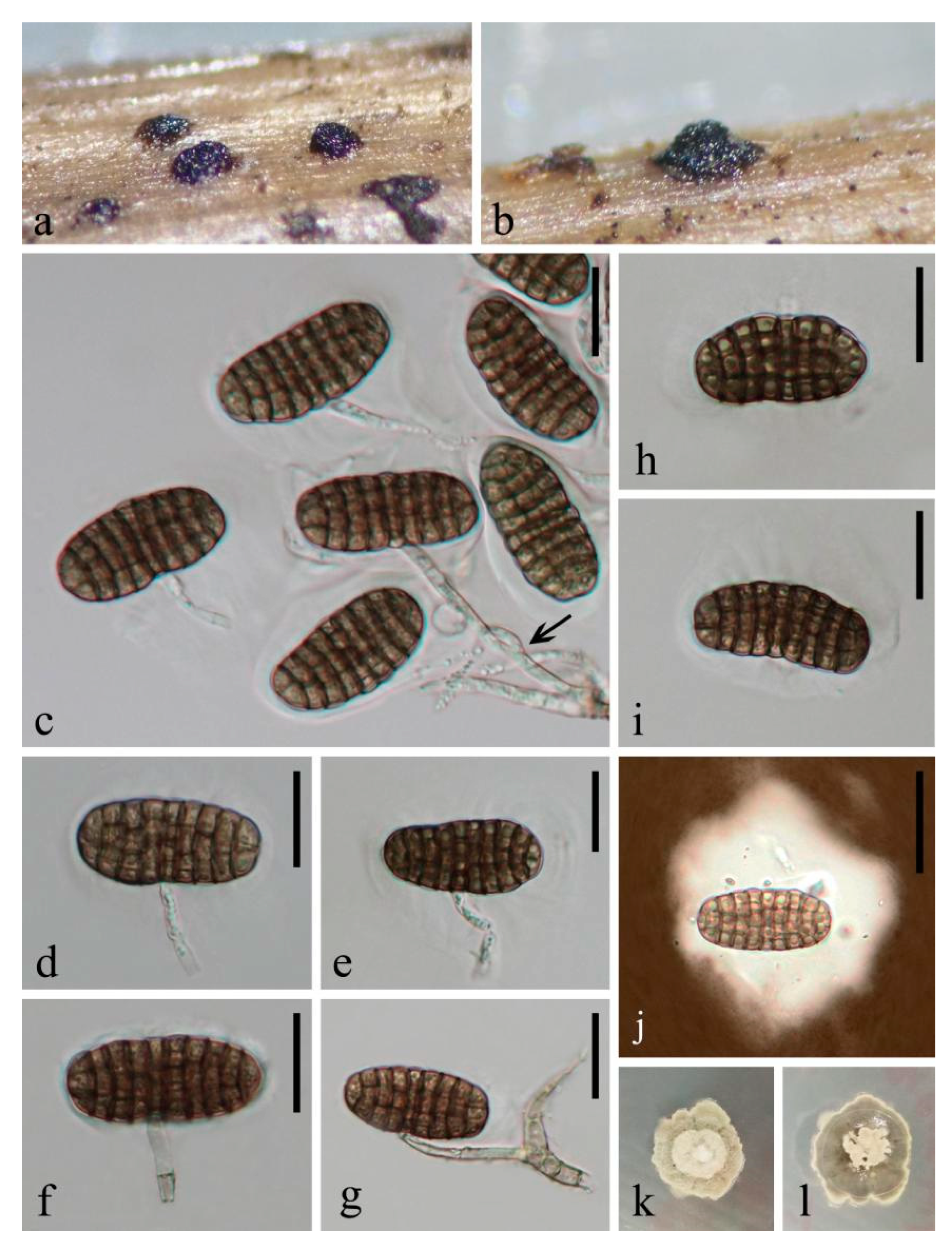
3.2. Taxonomy of Fungi Colonising Decaying Submerged Wood
3.2.1. Novel Taxa in Pleurotheciaceae
3.2.2. Novel Taxa in Savoryellomycetidae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hongsanan, S.; Maharachchikumbura, S.; Hyde, K.D.; Samarakoon, M.C.; Jeewon, R.; Zhao, Q.; Al-Sadi, A.; Bahkali, A.H. An updated phylogeny of Sordariomycetes based on phylogenetic and molecular clock evidence. Fungal Divers. 2017, 84, 25–41. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Maharachchikumbura, S.; Jones, E.B.G.; Dong, W.; Devadatha, B.; Yang, J.; Ekanayaka, A.H.; De Silva, W.; Sarma, V.V.; Al-Sadi, A.; et al. Phylogenetic Revision of Savoryellaceae and Evidence for Its Ranking as a Subclass. Front. Microbiol. 2019, 10, 840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Réblová, M.; Restrepo, M.H.; Fournier, J.; Nekvindová, J. New insights into the systematics of Bactrodesmium and its allies and introducing new genera, species and morphological patterns in the Pleurotheciales and Savoryellales (Sordariomycetes). Stud. Mycol. 2020, 95, 415–466. [Google Scholar] [CrossRef]
- Réblová, M.; Seifert, K.; Fournier, J.; Štěpánek, V. Newly recognised lineages of perithecial ascomycetes: The new orders Conioscyphales and Pleurotheciales. Pers. Mol. Phylogeny Evol. Fungi 2016, 37, 57–81. [Google Scholar] [CrossRef] [Green Version]
- Guarro, J.; Vieira, L.A.; De Freitas, D.; Gené, J.; Zaror, L.; Hofling-Lima, A.L.; Fischman, O.; Zorat-Yu, C.; Figueras, M.J. Phaeoisaria clematidis as a Cause of Keratomycosis. J. Clin. Microbiol. 2000, 38, 2434–2437. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.-L.; Hyde, K.D.; Liu, J.-K.; Maharachchikumbura, S.; Jeewon, R.; Bao, D.-F.; Bhat, D.J.; Lin, C.-G.; Li, W.-L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Restrepo, M.; Gené, J.; Castañeda-Ruiz, R.; Mena-Portales, J.; Crous, P.; Guarro, J. Phylogeny of saprobic microfungi from Southern Europe. Stud. Mycol. 2017, 86, 53–97. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.-G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Réblová, M.; Seifert, K.A.; Fournier, J.; Štěpánek, V. Phylogenetic classification of Pleurothecium and Pleurotheciella gen. nov. and its dactylaria-like anamorph (Sordariomycetes) based on nuclear ribosomal and protein-coding genes. Mycologia 2012, 104, 1299–1314. [Google Scholar] [CrossRef]
- Luo, Z.-L.; Hyde, K.D.; Bhat, D.J.; Jeewon, R.; Maharachchikumbura, S.S.N.; Bao, D.-F.; Li, W.-L.; Su, X.-J.; Yang, X.-Y.; Su, H.-Y. Morphological and molecular taxonomy of novel species Pleurotheciaceae from freshwater habitats in Yunnan, China. Mycol. Prog. 2018, 17, 511–530. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1–367. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Chethana, K.W.T.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.-Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Hyde, K.D.; Chaiwan, N.; Norphanphoun, C.; Boonmee, S.; Camporesi, E.; Chethana, K.W.T.; Dayarathne, M.C.; de Silva, N.I.; Dissanayake, A.J.; Ekanayaka, A.H.; et al. Mycosphere notes 169–224. Mycosphere 2018, 9, 271–430. [Google Scholar] [CrossRef]
- Su, H.-Y.; Udayanga, D.; Luo, Z.-L.; Manamgoda, D.; Zhao, Y.-C.; Yang, J.; Liu, X.-Y.; McKenzie, E.; Zhou, D.-Q.; Hyde, K. Hyphomycetes from aquatic habitats in Southern China: Species of Curvularia (Pleosporaceae) and Phragmocephala (Melannomataceae). Phytotaxa 2015, 226, 201–216. [Google Scholar] [CrossRef]
- Fallah, P.M.; Crane, J.L.; A Shearer, C. Freshwater ascomycetes: Two new species of Ascotaiwania from North America. Can. J. Bot. 1999, 77, 87–92. [Google Scholar]
- Fernández, F.A.; Lutzoni, F.M.; Huhndorf, S.M. Teleomorph-anamorph connections: The new pyrenomycetous genus Carpoligna and its Pleurothecium anamorph. Mycologia 1999, 91, 251–262. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Persoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.; Liu, X.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.-C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Hyde, K.D.; Doilom, M.; Yu, X.-D.; Bhat, D.J.; Jeewon, R.; Boonmee, S.; Wang, G.-N.; Nalumpang, S.; Zhang, H. Pseudobactrodesmium (Dactylosporaceae, Eurotiomycetes, Fungi) a Novel Lignicolous Genus. Front. Microbiol. 2020, 11, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Zhang, H.; Doilom, M.; Yu, X.-D.; Wang, G.-N.; Nalumpang, S. Rhexodenticula aquatica (Sordariomycetidae genera incertae sedis), a novel hyphomycete from freshwater in Thailand. Phytotaxa 2021, 483, 129–138. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. A simple method to control over-alignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar] [CrossRef]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop, New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Schwartz, T.; Pickett, B.; He, S.; Klem, E.B.; Scheuermann, R.H.; Passarotti, M.; Kaufman, S.; O’Leary, M.A. A RESTful API for Access to Phylogenetic Tools via the CIPRES Science Gateway. Evol. Bioinform. 2015, 11, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Larget, B.; Simon, D.L. Markov Chasin Monte Carlo Algorithms for the Bayesian Analysis of Phylogenetic Trees. Mol. Biol. Evol. 1999, 16, 750–759. [Google Scholar] [CrossRef]
- Doilom, M.; Dissanayake, A.J.; Wanasinghe, D.; Boonmee, S.; Liu, J.-K.; Bhat, D.J.; Taylor, J.E.; Bahkali, A.H.; McKenzie, E.H.C.; Hyde, K.D. Microfungi on Tectona grandis (teak) in Northern Thailand. Fungal Divers. 2017, 82, 107–182. [Google Scholar] [CrossRef]
- Charles, V.K. Coledictyospora, a new genus of dematiaceae. Phytopathology 1929, 19, 1051–1053. [Google Scholar]
- Matsushima, T. Matsushima mycological memoirs n. 5. Matsushima Fungus: Kobe. 1987. [Google Scholar]
- Fryar, S.; Davies, J.; Booth, W.; Hodgkiss, I.; Hyde, K. Succession of fungi on dead and live wood in brackish water in Brunei. Mycologia 2004, 96, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Nakagiri, A.; Ito, T. Some dematiaceous hyphomycetes on decomposing leaves of Satakentia liukiuensis from Ishigaki Island, Japan. IFO Res. Comm. 1995, 17, 75–98. [Google Scholar]
- Ho, W.H.; Yanna; Hyde, K.D.; Hodgkiss, I.J. Seasonality and sequential occurrence of fungi on wood submerged in Tai Po Kau Forest Stream, Hong Kong. Fungal Divers. 2002, 10, 21–43. [Google Scholar]
- Hernández, A.; Mena, J. Hifomicetos asociados a Coccothrinax (Palmae) en diferentes localidades de la Provincia de Camagüey (Cuba). Bol. Soc. Micol. Madr. 1995, 20, 25–33. [Google Scholar]
- Delgado, G. South Florida microfungi: Veramycella bispora, a new palmicolous anamorphic genus and species, with some new records for the continental USA. Mycotaxon 2009, 107, 357–373. [Google Scholar] [CrossRef]
- Whitton, S.R.; McKenzie, E.H.C.; Hyde, K.D. Anamorphic fungi associated with Pandanaceae. In Fungi Associated with Pandanaceae; Whitton, S.R., McKenzie, E.H.C., Hyde, K.D., Eds.; Springer: Dordrecht, The Netherland, 2012; pp. 125–353. [Google Scholar]
- Becerra Hernández, C.I.; Heredia, G.; Arias, R.M.; Mena Portales, J.; Castañeda Ruiz, R.F. Los hongos anamorfos saprobios del estado de Tabasco. III. Rev. Mex. Micol. 2008, 28, 25–39. [Google Scholar]
- Matsushima, T. Matsushima mycological memories n. 2. Matsushima Fungus: Kobe. 1981. [Google Scholar]
- Matsushima, T. Matsushima mycological memories n. 7. Matsushima Fungus: Kobe. 1993. [Google Scholar]
- Pinnoi, A.; Lumyong, S.; Hyde, K.D.; Jones, E.B.G. Biodiversity of fungi on the palm Eleiodoxa conferta in Sirindhorn peat swamp forest, Narathiwat, Thailand. Fungal Divers. 2006, 22, 205–218. [Google Scholar]
- Seifert, K.A.; Morgan-Jones, G.; Gams, W.; Kendrick, B. The genera of Hyphomycetes; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2011; pp. 1–997. [Google Scholar]
- Sun, L.Y.; Li, H.Y.; Sun, X.; Guo, L.D. Dematipyriforma aquilaria gen. et sp. nov., a new hyphomycetous taxon from Aquilaria crassna. Cryptogam. Mycol. 2017, 38, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Baker, W.A.; Partridge, E.C.; Morgan-Jones, G. Notes on hyphomycetes LXXXVII. Rhexoacrodictys, a new segregate genus to accommodate four species previously classified in Acrodictys. Mycotaxon 2002, 82, 95–113. [Google Scholar]
- Xiao, Z.-J.; Li, X.-X.; Wang, H.-D.; Song, P.-Y.; Tang, L. Rhexoacrodictys broussonetiae sp. nov. from Guizhou, China. Mycotaxon 2018, 133, 149–152. [Google Scholar] [CrossRef]
- Sivanesan, A.; Chang, H. Ascotaiwania, a new amphisphaeriaceous ascomycete genus on wood from Taiwan. Mycol. Res. 1992, 96, 481–484. [Google Scholar] [CrossRef]
- Chang, H.; Hsieh, S.-Y.; Jones, E.; Read, S.; Moss, S. New freshwater species of Ascotaiwania and Savoryella from Taiwan. Mycol. Res. 1998, 102, 709–718. [Google Scholar] [CrossRef]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Arzanlou, M.; Groenewald, J.; Gams, W.; Braun, U.; Shin, H.-D.; Crous, P. Phylogenetic and morphotaxonomic revision of Ramichloridium and allied genera. Stud. Mycol. 2007, 58, 57–93. [Google Scholar] [CrossRef]
- Boonmee, S.; Rossman, A.Y.; Liu, J.-K.; Li, W.-J.; Dai, D.-Q.; Bhat, J.D.; Jones, E.B.G.; McKenzie, E.H.C.; Xu, J.-C.; Hyde, K.D. Tubeufiales, ord. nov., integrating sexual and asexual generic names. Fungal Divers. 2014, 68, 239–298. [Google Scholar] [CrossRef]
- Lu, Y.-Z.; Liu, J.-K.; Hyde, K.D.; Jeewon, R.; Kang, J.-C.; Fan, C.; Boonmee, S.; Bhat, D.J.; Luo, Z.-L.; Lin, C.-G.; et al. A taxonomic reassessment of Tubeufiales based on multi-locus phylogeny and morphology. Fungal Divers. 2018, 92, 131–344. [Google Scholar] [CrossRef]
- Kodsueb, R.; Jeewon, R.; Vijaykrishna, D.; McKenzie, E.H.C.; Lumyong, P.; Lumyong, S.; Hyde, K.D. Systematic revision of Tubeufiaceae based on morphological and molecular data. Fungal Divers. 2006, 21, 105–130. [Google Scholar]
- Zhang, H.; Dong, W.; Hyde, K.D.; Maharachchikumbura, S.; Hongsanan, S.; Bhat, D.J.; Al-Sadi, A.; Zhang, D. Towards a natural classification of Annulatascaceae-like taxa: Introducing Atractosporales ord. nov. and six new families. Fungal Divers. 2017, 85, 75–110. [Google Scholar] [CrossRef]
- Dong, W.; Hyde, K.D.; Jeewon, R.; Doilom, M.; Yu, X.D.; Wang, G.N.; Liu, N.G.; Hu, D.M.; Nalumpang, S.; Zhang, H. Towards a natural classification of annulatascaceae-like taxa II: Introducing five new genera and eighteen new species from freshwater. Mycosphere 2021, 12, 1–88. [Google Scholar]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Xia, J.W.; Ma, Y.R.; Li, Z.; Zhang, X.G. Acrodictys-like wood decay fungi from southern China, with two new families Acrodictyaceae and Junewangiaceae. Sci. Rep. 2017, 7, 1–21. [Google Scholar] [CrossRef]
- Bucher, V.; Pointing, S.; Hyde, K.; Reddy, C. Production of Wood Decay Enzymes, Loss of Mass, and Lignin Solubilization in Wood by Diverse Tropical Freshwater Fungi. Microb. Ecol. 2004, 48, 331–337. [Google Scholar] [CrossRef]
- Hyde, K.D.; Fryar, S.; Tian, Q.; Bahkali, A.H.; Xu, J. Lignicolous freshwater fungi along a north–south latitudinal gradient in the Asian/Australian region; can we predict the impact of global warming on biodiversity and function? Fungal Ecol. 2016, 19, 190–200. [Google Scholar] [CrossRef]
- Fryar, S.C.; Booth, W.; Davies, J.; Hodgkiss, I.J.; Hyde, K.D. Distribution of fungi on wood in the Tutong River, Brunei. Fungal Divers. 2004, 17, 17–38. [Google Scholar]
- Kodsueb, R.; Lumyong, S.; McKenzie, E.; Bahkali, A.; Hyde, K. Relationships between terrestrial and freshwater lignicolous fungi. Fungal Ecol. 2016, 19, 155–168. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jeewon, R.; Chen, Y.-J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.-B.; Lin, C.-G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Ho, W.H.; Hyde, K.D.; Hodgkiss, J.I. Fungal communities on submerged wood from streams in Brunei, Hong Kong, and Malaysia. Mycol. Res. 2001, 105, 1492–1501. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Voucher/Culture | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| LSU | SSU | ITS | rpb2 | ||
| Achroceratosphaeria potamia | CBS 125414 | GQ996538 | GQ996541 | MH863679 | KM588908 |
| Adelosphaeria catenata | CBS 138679T | KT278707 | KT278692 | KT278721 | KT278743 |
| Anapleurothecium botulisporum | FMR 11490T | KY853483 | - | KY853423 | - |
| Arecophila bambusae | HKUCC 4794 | AF452038 | AY083802 | - | - |
| Ascotaiwania latericolla | ICMP 22739T | MN699407 | - | MN699390 | MN704312 |
| Ascotaiwania lignicola | NIL 00005 | HQ446364 | HQ446284 | - | HQ446419 |
| Bactrodesmiastrum obovatum | FMR 6482 | FR870266 | - | - | - |
| Bactrodesmiastrum pyriforme | FMR 11931 | HE646637 | - | - | - |
| Bactrodesmium diversum | CBS 144080 | MN699415 | MN699371 | MN699355 | MN704294 |
| Bactrodesmium obovatum | CBS 144078 | MN699425 | MN699376 | MN699396 | - |
| Bactrodesmium pallidum | CBS 145349 | MN699429 | MN699380 | MN699364 | MN704302 |
| Brachysporiella setosa | HKUCC 3713 | AF132334 | - | - | - |
| Canalisporium exiguum | SS 00809 | GQ390281 | GQ390266 | - | HQ446436 |
| Canalisporium grenadoideum | BCC 20507 | GQ390267 | GQ390252 | - | HQ446420 |
| Cercophora caudata | CBS 606.72 | AY999113 | DQ368659 | AY999135 | DQ368646 |
| Cercophora newfieldiana | SMH 3303 | AY780062 | - | - | AY780167 |
| Cercophora thailandica | MFLUCC 12-0845 | KU863127 | KU872131 | - | KU940176 |
| Cheilymenia stercorea | AFTOL 148 | AY544661 | AY544705 | - | - |
| Coleodictyospora muriformis | MFLUCC 18-1243T | MW981648 | MW981704 | MW981642 | - |
| Coleodictyospora muriformis | MFLUCC 18-1279T | MW981649 | MW981705 | MW981643 | - |
| Conioscypha japonica | CBS 387.84 | AY484514 | JQ437438 | - | - |
| Conioscypha lignicola | CBS 335.93T | AY484513 | JQ437439 | - | JQ429260 |
| Conioscypha peruviana | ILL 41202T | KF781539 | - | - | - |
| Conioscypha varia | CBS 604.70 | MH871656 | - | MH859869 | - |
| Cosmospora arxii | CBS 748.69 | MH871181 | - | NR-145062 | HQ897725 |
| Dematiosporium aquaticum | MFLU 18-1641T | MK835855 | - | - | MN194029 |
| Dematipyriforma aquilaria | CGMCC 3.17268T | KJ138623 | KJ138622 | KJ138621 | - |
| Diaporthe cyatheae | YMJ 1364 | JX570891 | JX570890 | - | JX570893 |
| Diaporthe eres | AR 3538 | AF408350 | - | - | - |
| Diaporthe xishuangbanica | LC6744 | KY011862 | - | - | - |
| Doratomyces stemonitis | AFTOL-ID 1380 | DQ836907 | DQ836901 | - | - |
| Entosordaria perfidiosa | BW3 | MF488992 | - | - | MF489002 |
| Fuscosporella pyriformis | MFLUCC 16-0570T | KX550896 | KX550900 | - | KX576872 |
| Fusicolla aquaeductuum | KUMCC 18-0015 | MH087221 | - | MH087219 | - |
| Helicoascotaiwania farinosa | DAOMC 241947 | JQ429230 | - | JQ429145 | - |
| Helicoascotaiwania farinosa | ILLS 53605T | AY094189 | - | - | - |
| Helicoascotaiwania farinosa | P2-6 | AY316357 | - | - | - |
| Helicoascotaiwania lacustris | CBS 145963T | MN699430 | MN699382 | MN699399 | MN704304 |
| Helicoascotaiwania lacustris | CBS 145964 | MN699431 | MN699383 | MN699400 | MN704305 |
| Helicoascotaiwania lacustris | CBS 146144 | MN699432 | MN699384 | MN699401 | MN704306 |
| Melanotrigonum ovale | CBS 138744 | KT278710 | KT278697 | KT278725 | KT278746 |
| Melanotrigonum ovale | CBS 138815 | KT278711 | KT278698 | KT278722 | KT278747 |
| Melanotrigonum ovale | CBS 138743T | KT278709 | KT278696 | KT278724 | KT278745 |
| Microascus trigonosporus | AFTOL-ID 914 | DQ470958 | DQ471006 | DQ491513 | - |
| Mucispora obscuriseptata | MFLUCC 15-0618T | KX550892 | KX550897 | - | KX576870 |
| Nectria nigrescens | AR 4211 | HM484720 | JN939647 | HM484707 | JQ014123 |
| Neoascotaiwania fusiformis | MFLUCC 15-0621T | KX550893 | - | MG388215 | KX576871 |
| Neoascotaiwania limnetica | CBS 126576 | KY853513 | KT278689 | KY853452 | MN704308 |
| Neoascotaiwania terrestris | CBS 142291T | KY853515 | KY853547 | KY853454 | - |
| Neomonodictys muriformis | MFLUCC 16-1136T | MN644485 | - | MN644509 | - |
| Neoroussoella alishanense | MFLUCC 11-0190 | MN028398 | - | MN028394 | - |
| Neoroussoella bambusae | MFLUCC 11-0124 | KJ474839 | - | KJ474827 | KJ474856 |
| Neotorula submersa | KUMCC 15-0280 | KX789217 | - | KX789214 | - |
| Neurospora crassa | MUCL 19026 | AF286411 | X04971 | - | - |
| Obliquifusoideum guttulatum | MFLUCC 18-1233T | MW981650 | MW981706 | MW981645 | - |
| Paracremonium binnewijzendii | CBS 143277 | MG250174 | - | NR-157491 | - |
| Parathyridaria percutanea | CBS 868.95 | KF366449 | KF366451 | KF322118 | KF366452 |
| Phaeoisaria annesophieae | MFLU 19-0531 | MT559084 | - | MT559109 | - |
| Phaeoisaria aquatica | MFLUCC 16-1298T | MF399254 | - | MF399237 | MF401406 |
| Phaeoisaria clematidis | MFLUCC 18-1017 | MW132065 | MW132063 | MW131990 | - |
| Phaeoisaria fasciculata | CBS 127885T | KT278705 | KT278693 | KT278719 | KT278741 |
| Phaeoisaria fasciculata | DAOM 230055 | KT278706 | KT278694 | KT278720 | KT278742 |
| Phaeoisaria pseudoclematidis | MFLUCC 11-0393T | KP744501 | KP753962 | KP744457 | - |
| Phaeoisaria sedimenticola | CGMCC 3.14949T | JQ031561 | - | JQ074237 | - |
| Phragmocephala stemphylioides | DAOM 673211 | KT278717 | - | KT278730 | - |
| Pisorisporium cymbiforme | PRM 924378 | KM588902 | KM588899 | - | KM588905 |
| Pleurotheciella aquatica | MFLUCC 17-0464T | MF399253 | MF399220 | MF399236 | MF401405 |
| Pleurotheciella centenaria | DAOM 229631T | JQ429234 | JQ429246 | JQ429151 | JQ429265 |
| Pleurotheciella erumpens | CBS 142447T | MN699435 | MN699387 | MN699406 | MN704311 |
| Pleurotheciella fusiformis | MFLUCC 17-0113T | MF399250 | MF399218 | MF399233 | MF401403 |
| Pleurotheciella guttulata | KUMCC 15-0442 | MF399256 | MF399222 | MF399239 | MF401408 |
| Pleurotheciella guttulata | KUMCC 15-0296T | MF399257 | MF399223 | MF399240 | MF401409 |
| Pleurotheciella krabiensis | MFLUCC 16-0852T | MG837013 | MG837023 | MG837018 | - |
| Pleurotheciella lunata | MFLUCC 17-0111T | MF399255 | MF399221 | MF399238 | MF401407 |
| Pleurotheciella rivularia | CBS 125238T | JQ429232 | JQ429244 | JQ429160 | JQ429263 |
| Pleurotheciella rivularia | CBS 125237 | JQ429233 | JQ429245 | JQ429161 | JQ429264 |
| Pleurotheciella saprophytica | MFLUCC 16-1251T | MF399258 | MF399224 | MF399241 | MF401410 |
| Pleurotheciella submersa | DLUCC 0739 | MF399259 | MF399225 | MF399242 | MF401411 |
| Pleurotheciella submersa | MFLUCC 17-1709T | MF399260 | MF399226 | MF399243 | MF401412 |
| Pleurotheciella sympodia | MFLUCC 18-1408 | MW981652 | - | MW981644 | - |
| Pleurotheciella sympodia | MFLUCC 15-0996 | MW981651 | MW981703 | MW981641 | - |
| Pleurotheciella sympodia | MFLUCC 18-0658 | MT559086 | MT559094 | MT555418 | - |
| Pleurotheciella sympodia | MFLUCC 18-0983 | MT555425 | MT555734 | MT555419 | - |
| Pleurotheciella sympodia | KUMCC 19-0213 | MT555426 | - | MT555420 | - |
| Pleurotheciella tropica | MFLUCC 16-0867T | MG837015 | MG837025 | MG837020 | - |
| Pleurotheciella uniseptata | DAOM 673210T | KT278716 | - | KT278729 | - |
| Pleurotheciella uniseptata | KUMCC 15-0407 | MF399248 | - | MF399231 | MF401401 |
| Pleurothecium aquaticum | MFLUCC 17-1331T | MF399263 | - | MF399245 | - |
| Pleurothecium aquaticum | B-27 | MK835854 | MK834786 | - | - |
| Pleurothecium floriforme | MFLUCC 15-0628 | KY697277 | KY697279 | KY697281 | - |
| Pleurothecium obovoideum | CBS 209.95 | EU041841 | - | EU041784 | - |
| Pleurothecium pulneyense | MFLUCC 16-1293 | MF399262 | MF399228 | - | MF401414 |
| Pleurothecium recurvatum | DAOM 230069 | JQ429238 | JQ429252 | JQ429157 | JQ429269 |
| Pleurothecium semifecundum | CBS 131271T | JQ429240 | JQ429254 | JQ429159 | JQ429270 |
| Podosordaria tulasnei | CBS 128.80 | KT281897 | - | - | - |
| Pseudoascotaiwania persoonii | A57-14C | AY094190 | - | - | - |
| Pseudocoleodictyospora sukhothaiensis | MFLUCC 12-0554 | KU764710 | KU712471 | KU712440 | KU712493 |
| Pseudocoleodictyospora tectonae | MFLUCC 12-0385 | KU764709 | KU712461 | KU712443 | KU712491 |
| Pseudocoleodictyospora tectonae | MFLUCC 12-0387 | KU764704 | KU712462 | KU712444 | KU712492 |
| Pseudocoleodictyospora thailandica | MFLUCC 12-0565 | KU764701 | KU712472 | KU712441 | KU712494 |
| Pseudoneurospora amorphoporcata | CBS 626.80 | FR774287 | - | - | - |
| Rhexoacrodictys erecta | HSAUPmyr4622 | KX033556 | KX033526 | KU999964 | - |
| Rhexoacrodictys erecta | HSAUPmyr6489 | KX033555 | KX033525 | KU999963 | - |
| Rhexoacrodictys erecta | KUMCC 20-0194 | MT559123 | - | MT555421 | - |
| Rhexoacrodictys fimicola | HMAS47737 | KX033553 | KX033522 | KU999960 | - |
| Rhexoacrodictys fimicola | HMAS43690 | KX033550 | KX033519 | KU999957 | - |
| Roussoella nitidula | MFLUCC 11-0182 | KJ474843 | - | KJ474835 | KJ474859 |
| Saprodesmium dematiosporum | KUMCC 18-0059T | MW981647 | MW981707 | MW981646 | - |
| Savoryella aquatica | SS 03801 | HQ446372 | HQ446290 | - | HQ446441 |
| Savoryella lignicola | NF00204 | HQ446378 | HQ446300 | HQ446357 | - |
| Savoryella longispora | SAT 00322 | HQ446380 | HQ446302 | HQ446359 | HQ446450 |
| Savoryella paucispora | SAT 00866 | HQ446381 | HQ446303 | - | HQ446451 |
| Savoryella verrucosa | SS 00052 | HQ446374 | - | HQ446353 | HQ446445 |
| Savoryella yunnanensis | MFLUCC 18-1395T | MK411422 | MK411423 | - | - |
| Sordaria fimicola | CBS 508.50 | AY681160 | - | - | - |
| Sterigmatobotrys macrocarpa | PRM 915682 | GU017317 | JQ429255 | JQ429153 | - |
| Sterigmatobotrys macrocarpa | DAOM 230059 | GU017316 | - | JQ429154 | JQ429271 |
| Sterigmatobotrys rudis | DAOM 229838 | JQ429241 | JQ429256 | JQ429152 | JQ429272 |
| Sterigmatobotrys uniseptata | MFLUCC 15-0358T | MK835850 | MK834784 | MK878379 | - |
| Subglobosporium tectonae | MFLUCC 12-0393 | KU764703 | KU712464 | KU712445 | KU712485 |
| Subglobosporium tectonae | MFLUCC 12-0390 | KU764702 | KU712463 | KU712446 | KU712495 |
| Thyridaria broussonetiae | CBS 141481 | KX650568 | - | NR-147658 | KX650586 |
| Torula aquatica | MFLUCC 16-1115 | MG208146 | - | MG208167 | MG207977 |
| Torula herbarum | CPC 24114 | KR873288 | - | KR873260 | - |
| Triadelphia uniseptata | TA06NZ-142 | KT278718 | - | - | - |
| Tubakia seoraksanensis | CBS 127490 | KP260499 | - | - | - |
| Xylaria hypoxylon | CBS 122620 | KY610495 | - | KY610407 | KY624231 |
| Zalerion maritima | FCUL280207CP1 | JN886806 | KT347203 | KT347216 | - |
| Zalerion xylestrix | 309156 | EU848592 | EU848591 | - | - |
| C. cubensis | C. micronesica | C. muriformis | |
|---|---|---|---|
| Conidiophores | 70–85 × 3.5–5 μm | Reduced, 2–8 × 3–4 μm (on CMA) | Up to 55 μm long, 3 μm wide |
| Conidiophore attaching point | Middle | End | Middle |
| Conidia | 42–50 × 20–22 μm | 30–40 × 13–16 μm | 32–44 × 15.5–19 μm |
| Conidial sheaths | 55–60 × 40–45 μm | Present, but not measured | Up to 55 μm thick in Indian Ink |
| Conidial transverse septa | 8–14 | 6–9 | (7–)8–9 |
| Conidial septa with or without dark bands | Not mentioned | Not mentioned | With dark brown bands |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, W.; Jeewon, R.; Hyde, K.D.; Yang, E.-F.; Zhang, H.; Yu, X.; Wang, G.; Suwannarach, N.; Doilom, M.; Dong, Z. Five Novel Taxa from Freshwater Habitats and New Taxonomic Insights of Pleurotheciales and Savoryellomycetidae. J. Fungi 2021, 7, 711. https://doi.org/10.3390/jof7090711
Dong W, Jeewon R, Hyde KD, Yang E-F, Zhang H, Yu X, Wang G, Suwannarach N, Doilom M, Dong Z. Five Novel Taxa from Freshwater Habitats and New Taxonomic Insights of Pleurotheciales and Savoryellomycetidae. Journal of Fungi. 2021; 7(9):711. https://doi.org/10.3390/jof7090711
Chicago/Turabian StyleDong, Wei, Rajesh Jeewon, Kevin D. Hyde, Er-Fu Yang, Huang Zhang, Xiandong Yu, Gennuo Wang, Nakarin Suwannarach, Mingkwan Doilom, and Zhangyong Dong. 2021. "Five Novel Taxa from Freshwater Habitats and New Taxonomic Insights of Pleurotheciales and Savoryellomycetidae" Journal of Fungi 7, no. 9: 711. https://doi.org/10.3390/jof7090711