
Lebanon’s Native Oenological Saccharomyces cerevisiae Flora: Assessment of Different Aspects of Genetic Diversity and Evaluation of Winemaking Potential

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Analysis Scheme
2.2. Strains Identification
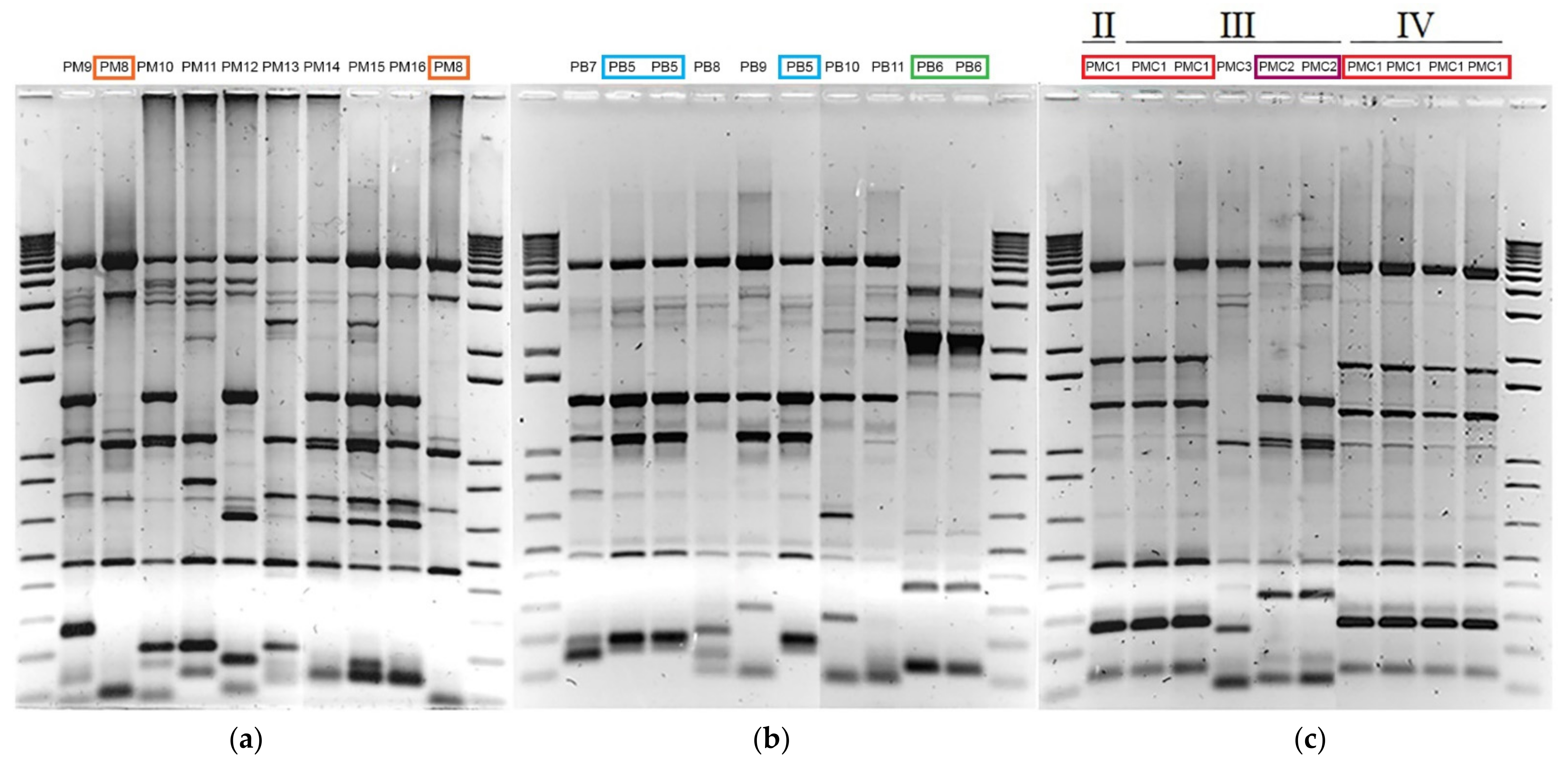
2.3. Inter-Delta Fingerprinting
2.4. Microsatellites Analysis
2.5. Strains Choice for Technological Characterization and Fermentations
2.6. Lab-Scale Fermentations and Chemical Analysis of Wines
2.7. Fermentation Results Analysis and Sensory Paired Comparison Test
3. Results
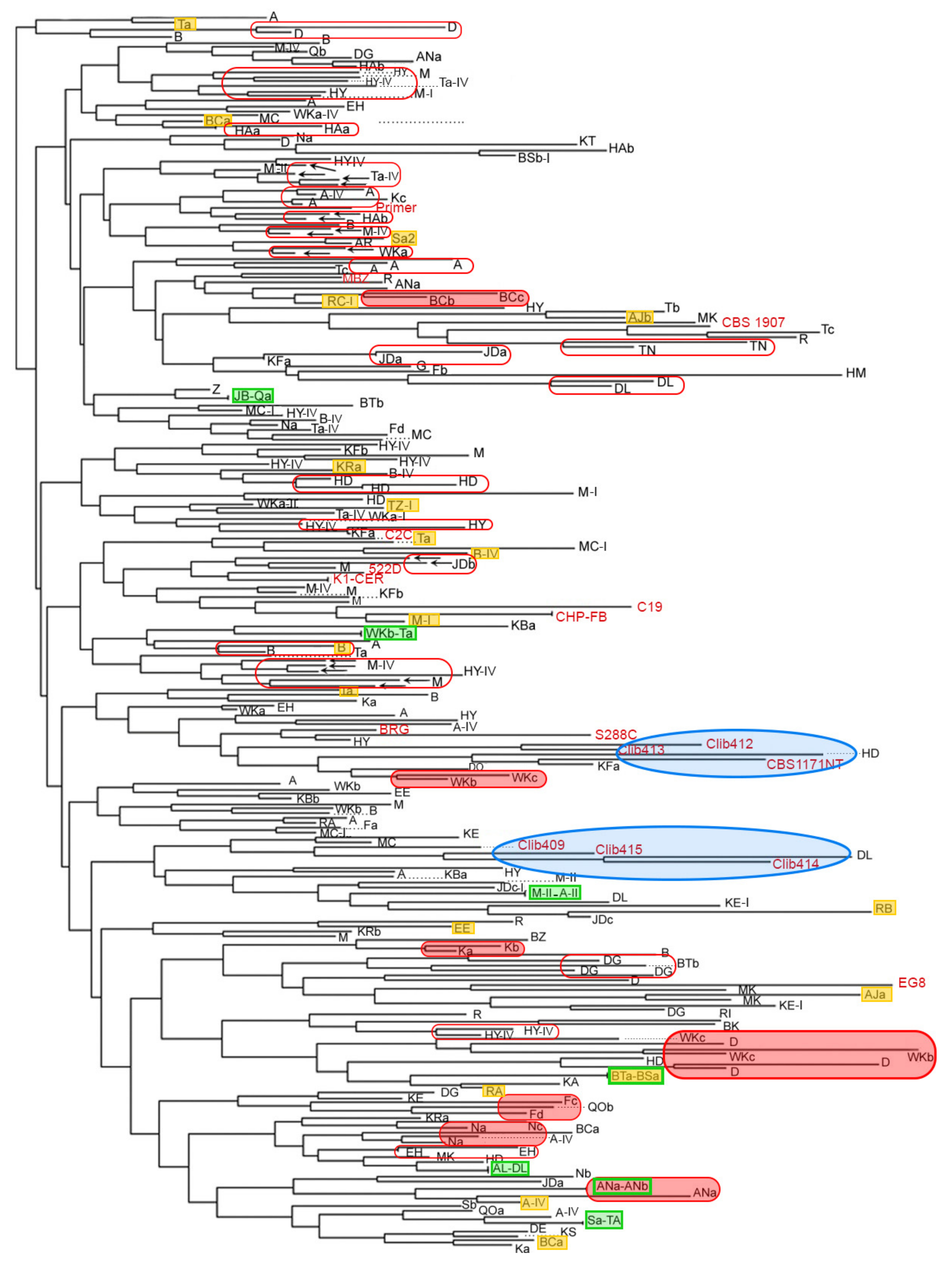
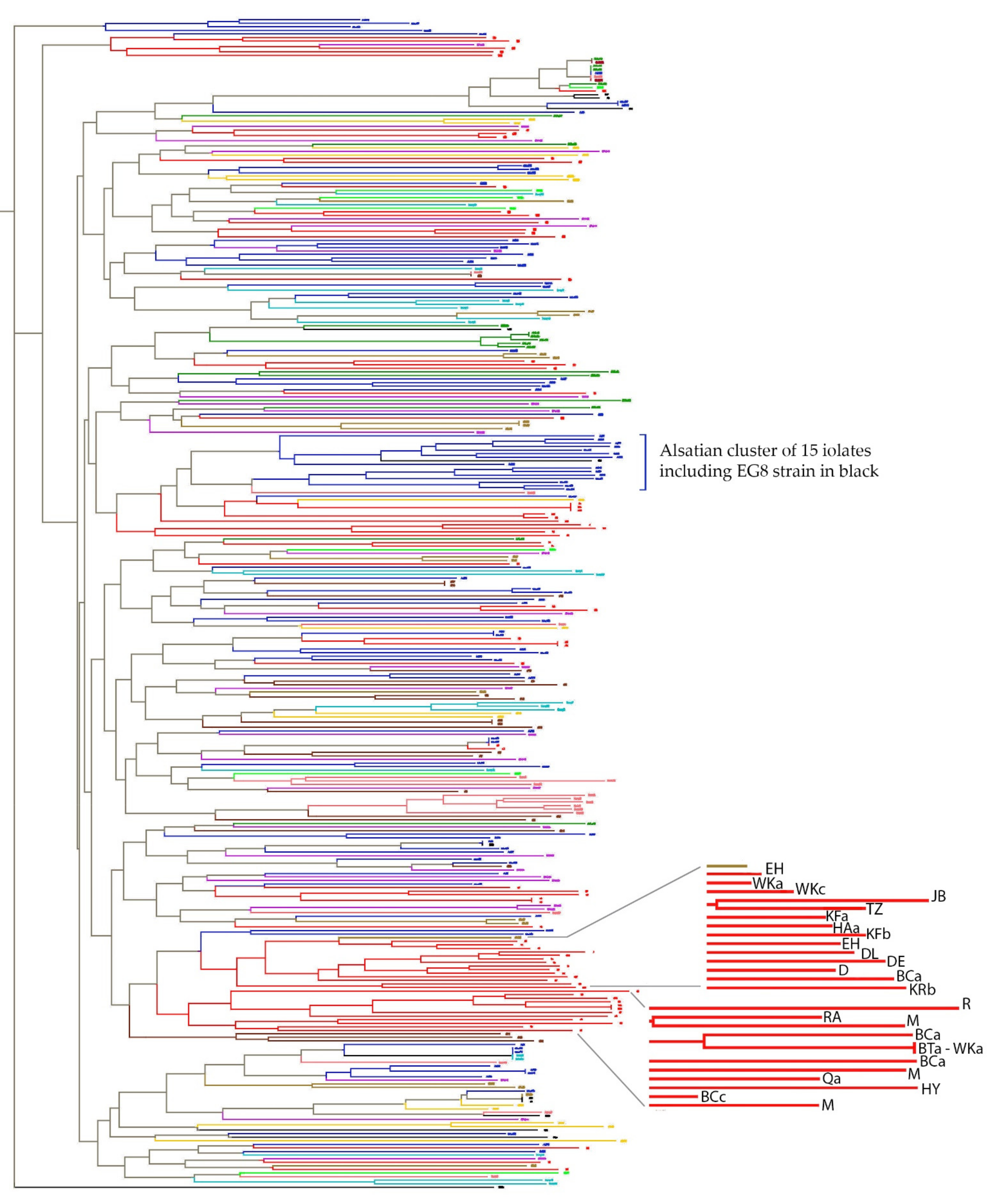
3.1. Diversity of the Natural Lebanese Oenological Flora across the Territory
3.2. Diversity of the Oenological Flora in Fermenting Vats and Wineries
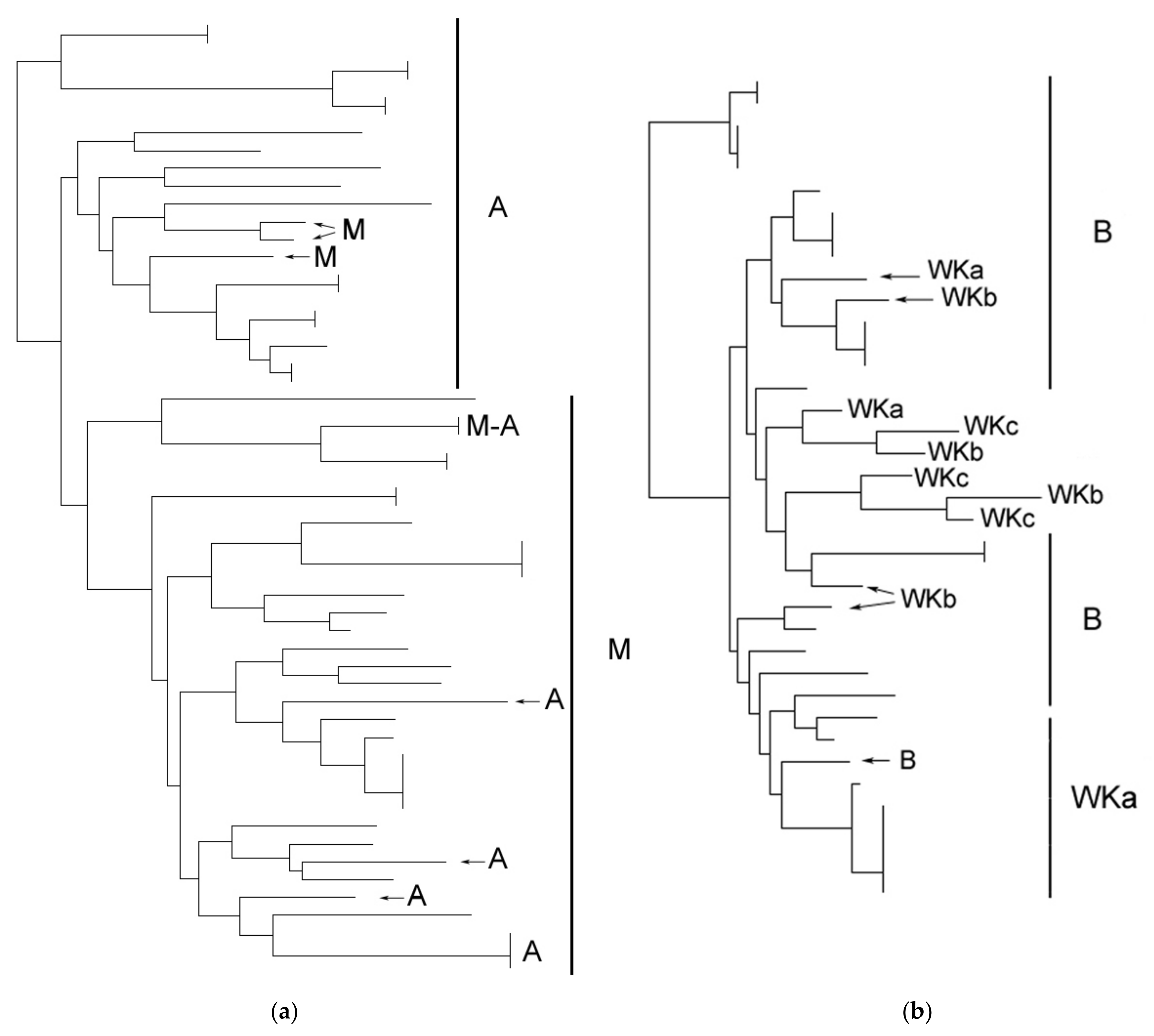
3.3. Stability of Strains in Wineries
3.4. Winery and Geographic Vicinity Effects on Strain Relatedness
3.5. Comparison of Lebanese Isolates with Floras of Diverse Origins
3.6. Lab-Scale Fermentations and Technological Screening
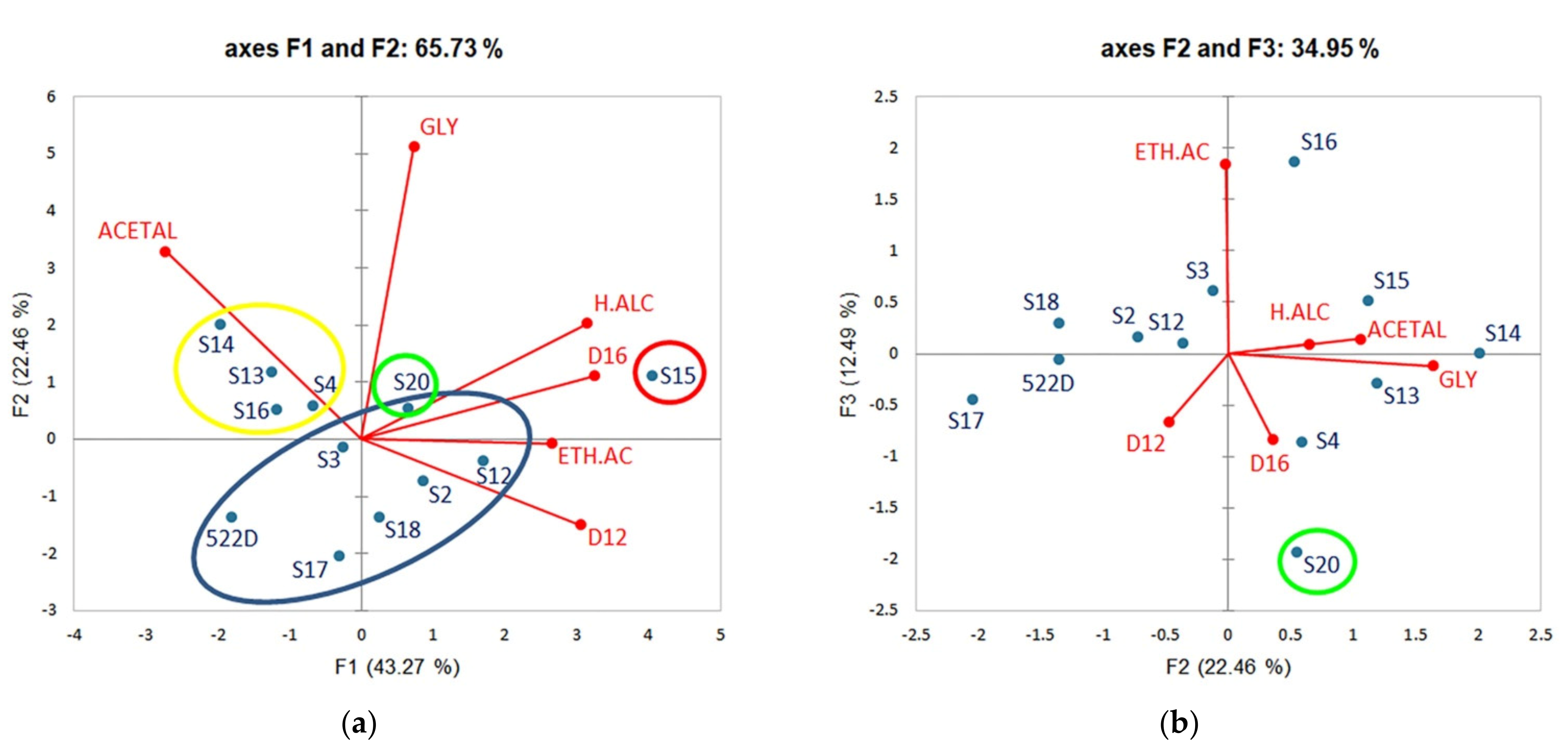
3.7. PCA Distribution of the Strains According to Oenological Parameters
3.8. Proposition of Strains for Selection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hosea. The Bible. English Standard Version ESV; 14:7; Crossway: Wheaton, IL, USA, 2016. [Google Scholar]
- Parker, S. Ugaritic Narrative Poetry; Scholars Press: Atlanta, GA, USA, 1997; p. 265. [Google Scholar]
- Ballard, R.D.; Stager, L.E.; Master, D.; Yoerger, D.; Mindell, D.; Whitcomb, L.; Singh, H.; Piechota, D. Iron Age Shipwrecks in Deep Water Off Ashkelon. Am. J. Archaeol. 2002, 106, 151–168. [Google Scholar] [CrossRef]
- Orsingher, A.; Amicone, S.; Kamlah, J.; Sader, H.; Berthold, C. Phoenician lime for Phoenician wine: Iron Age plaster from a wine press at Tell el-Burak, Lebanon. Antiquity 2020, 94, 377, 1224–1244. [Google Scholar] [CrossRef]
- Legras, J.L.; Merdinoglu, D.; Cornuet, J.M.; Karst, F. Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Botto, M. The Phoenicians and the spread of wine in the central West Mediterranean. In Patrimonio Cultural de la Vid y el Vino: Conferencia Internacional; Pérez, S.C., Pérez, J.B., Eds.; UAM Ediciones: Madrid, Spain, 2013; pp. 103–131. [Google Scholar]
- Sicard, D.; Legras, J.-L. Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. C. R. Biol. 2011, 334, 229–236. [Google Scholar] [CrossRef]
- Schacherer, J.; Shapiro, J.A.; Ruderfer, D.M.; Kruglyak, L. Comprehensive polymorphism survey elucidates population structure of Saccharomyces cerevisiae. Nature 2009, 458, 342–345. [Google Scholar] [CrossRef] [Green Version]
- Schutz, M.; Gafner, J. Analysis of yeast diversity during spontaneous and induced alcoholic fermentations. J. Appl. Bacteriol. 1993, 75, 551–558. [Google Scholar] [CrossRef]
- Constanti, M.; Poblet, M.; Arola, L.; Mas, A.; Guillamon, J. Analysis of yeast populations during alcoholic fermentation in a newly established winery. Am. J. Enol. Vitic. 1997, 48, 339–344. [Google Scholar]
- Guillamon, J.M.; Sabate, J.; Barrio, E.; Cano, J.; Querol, A. Rapid identification of wine yeast species based on RFLP analysis of the ribosomal internal transcribed spacer (ITS) region. Arch. Microbiol. 1998, 169, 387–392. [Google Scholar] [CrossRef]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Garofalo, C.; Tristezza, M.; Grieco, F.; Spano, G.; Capozzi, V. From grape berries to wine: Population dynamics of cultivable yeasts associated to “Nero di Troia” autochthonous grape cultivar. World J. Microbiol. Biotechnol. 2016, 32, 59. [Google Scholar] [CrossRef]
- Cavalieri, D.; McGovern, P.E.; Hartl, D.L.; Mortimer, R.; Polsinelli, M. Evidence for S. cerevisiae fermentation in ancient wine. J. Mol. Evol. 2003, 57, S226–S232. [Google Scholar] [CrossRef] [Green Version]
- Schutz, M.; Gafner, J. Dynamics of the yeast strain population during spontaneous alcoholic fermentation determined by CHEF gel electrophoresis. Lett. Appl. Microbiol. 1994, 19, 253–257. [Google Scholar] [CrossRef]
- Sabate, J.; Cano, J.; Querol, A.; Guillamon, J.M. Diversity of Saccharomyces strains in wine fermentations: Analysis for two consecutive years. Lett. Appl. Microbiol. 1998, 28, 452–455. [Google Scholar] [CrossRef] [Green Version]
- Pramateftaki, P.V.; Lanaridis, P.; Typas, M.A. Molecular identification of wine yeasts at species or strain level: A case study with strains from two vine-growing areas of Greece. J. Appl. Microbiol. 2000, 89, 236–248. [Google Scholar] [CrossRef] [Green Version]
- Lopes, C.A.; van Broock, M.; Querol, A.; Caballero, A.C. Saccharomyces cerevisiae wine yeast populations in a cold region in Argentinean Patagonia. A study at different fermentation scales. J. Appl. Microbiol. 2002, 93, 608–615. [Google Scholar] [CrossRef]
- Börlin, M.; Venet, P.; Claisse, O.; Salin, F.; Legras, J.-L.; Masneuf-Pomarède, I. Cellar associated Saccharomyces cerevisiae population structure revealed high diversity and perennial persistence in Sauternes wine estates. Appl. Environ. Microbiol. 2016, 82, 2909–2918. [Google Scholar] [CrossRef] [Green Version]
- Vigentini, I.; Maghradze, D.; Petrozziello, M.; Bonello, F.; Mezzapelle, V.; Valdetara, F.; Failla, O.; Foschino, R. Indigenous Georgian wine-associated yeasts and grape cultivars to edit the wine quality in a precision oenology perspective. Front. Microbiol. 2016, 7, 352. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, P.; López, R.; Garijo, M.; Escribano, R.; González-Arenzana, L.; López-Alfaro, I.; Gutiérrez, A.R. Biodiversity of Saccharomyces cerevisiae Yeasts in Spontaneous Alcoholic Fermentations: Typical Cellar or Zone Strains? In Advances in Grape and Wine Biotechnology; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Versavaud, A.; Courcoux, P.; Roulland, C.; Dulau, L.; Hallet, J.N. Genetic Diversity and Geographical Distribution of Wild Saccharomyces cerevisiae Strains from the Wine-Producing Area of Charentes, France. Appl. Environ. Microbiol. 1995, 61, 3521–3529. [Google Scholar] [CrossRef] [Green Version]
- Torija, M.J.; Rozes, N.; Poblet, M.; Guillamon, J.M.; Mas, A. Yeast population dynamics in spontaneous fermentations: Comparison between two different wine-producing areas over a period of three years. Antonie Van Leeuwenhoek 2001, 79, 345–352. [Google Scholar] [CrossRef]
- Querol, A.; Barrio, E.; Huerta, T.; Ramon, D. Molecular monitoring of wine fermentations conducted by active dry yeast strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef] [Green Version]
- Egli, C.M.; Edinger, W.D.; Mitrakul, C.M.; Henick-Kling, T. Dynamics of indigenous and inoculated yeast populations and their effect on the sensory character of Riesling and Chardonnay wines. J. Appl. Microbiol. 1998, 85, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Ganga, M.A.; Martínez, C. Effect of wine yeast monoculture practice on the biodiversity of non-Saccharomyces yeasts. J. Appl. Microbiol. 2004, 96, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Le Jeune, C.; Erny, C.; Demuyter, C.; Lollier, M. Evolution of the population of Saccharomyces cerevisiae from grape to wine in a spontaneous fermentation. Food Microbiol. 2006, 23, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Vezinhet, F.; Hallet, J.-N.; Valade, M.; Poulard, A. Ecological survey of wine yeast strains by molecular methods of identification. Am. J. Enol. Vitic. 1992, 43, 83–86. [Google Scholar]
- Beltran, G.; Torija, M.J.; Novo, M.; Ferrer, N.; Poblet, M.; Guillamón, J.; Rozès, N.; Mas, A. Analysis of yeast populations during alcoholic fermentation: A six year follow-up study. Syst. Appl. Microbiol. 2002, 5, 287–293. [Google Scholar] [CrossRef]
- Sangorrín, M.; Zajonskovsky, I.; van Broock, M.; Caballero, A. The use of killer biotyping in an ecological survey of yeast in an old patagonian winery. World J. Microbiol. Biotechnol. 2002, 18, 115–120. [Google Scholar] [CrossRef]
- Frezier, V.; Duburdieu, D. Ecology of yeast strain Saccharomyces cerevisiae during spontaneous fermentation in a Bordeaux winery. Am. J. Enol. Vitic. 1992, 43, 375–380. [Google Scholar]
- Gutiérrez, A.R.; Santamaría, P.; Epifanio, S.; Garijo, P.; López, R. Ecology of spontaneous fermentation in one winery during 5 consecutive years. Lett. Appl. Microbiol. 1999, 29, 411–415. [Google Scholar] [CrossRef]
- Schuller, D.; Alves, H.; Dequin, S.; Casal, M. Ecological survey of Saccharomyces cerevisiae strains from vineyards in the Vinho Verde Region of Portugal. FEMS Microbiol. Ecol. 2005, 51, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Valero, E.; Schuller, D.; Cambon, B.; Casal, M.; Dequin, S. Dissemination and survival of commercial wine yeast in the vineyard: A large-scale, three-years study. FEMS Yeast Res. 2005, 5, 959–969. [Google Scholar] [CrossRef] [Green Version]
- Schuller, D.; Cardoso, F.; Sousa, S.; Gomes, P.; Gomes, A.C.; Santos, M.; Casal, M. Genetic Diversity and Population Structure of Saccharomyces cerevisiae Strains Isolated from Different Grape Varieties and Winemaking Regions. PLoS ONE 2012, 7, e32507. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Zarraonaindia, I.; Perisin, M.; Palacios, A.; Acedo, A. From vineyard soil to wine fermentation: Microbiome approximations to explain the “terroir” concept. Front. Microbiol. 2017, 8, 821. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; van der Lelie, D.; Zarraonaindia, I. Microbial terroir for wine grapes. Proc. Nat. Acad. Sci. USA 2014, 111, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Pinto, C.; Pinho, D.; Cardoso, R.; Custódio, V.; Fernandes, J.; Susana, S.; Pinheiro, M.; Egas, C.; Gomes, A. Wine fermentation microbiome: A landscape from different Portuguese wine appellations. Front. Microbiol. 2015, 6, 905. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.W.; Tsai, P.; Anfang, N.; Ross, H.A.; Goddard, M.R. Pyrosequencing reveals regional differences in fruit-associated fungal communities. Environ. Microbiol. 2014, 16, 2848–2858. [Google Scholar] [CrossRef] [Green Version]
- Morrison-Whittle, P.; Goddard, M.R. From vineyard to winery: A source map of microbial diversity driving wine fermentation. Environ. Microbiol. 2018, 20, 75. [Google Scholar] [CrossRef] [Green Version]
- Gayevskiy, V.; Goddard, M.R. Geographic delineations of yeast communities and populations associated with vines and wines in New Zealand. ISME J. 2012, 6, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Knight, S.; Goddard, M.R. Quantifying separation and similarity in a Saccharomyces cerevisiae metapopulation. ISME J. 2015, 9, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Viel, A.; Legras, J.-L.; Nadai, C.; Carlot, M.; Lombardi, A.; Crespan, M.; Migliaro, D.; Giacomini, A.; Corich, V. The Geographic Distribution of Saccharomyces cerevisiae Isolates within three Italian Neighboring Winemaking Regions Reveals Strong Differences in Yeast Abundance, Genetic Diversity and Industrial Strain Dissemination. Front. Microbiol. 2017, 8, 1595. [Google Scholar] [CrossRef] [Green Version]
- Drumonde-Neves, J.; Franco-Duarte, R.; Vieira, E.; Mendes, I.; Lima, T.; Schuller, D.; Pais, C. Differentiation of Saccharomyces cerevisiae populations from vineyards of the Azores Archipelago: Geography vs. Ecology. Food Microbiol. 2018, 74, 151–162. [Google Scholar] [CrossRef]
- Knight, S.; Klaere, S.; Fedrizzi, B.; Goddard, M.R. Regional microbial signatures positively correlate with differential wine phenotypes: Evidence for a microbial aspect to terroir. Sci. Rep. 2015, 5, 14233. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. mBio 2016, 7, e00631-16. [Google Scholar] [CrossRef] [Green Version]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Lopes, C.A.; Rodríguez, M.E.; Sangorrín, M.; Querol, A.; Caballero, A.C. Patagonian wines: The selection of an indigenous yeast starter. J. Ind. Microbiol. Biotechnol. 2007, 34, 539–546. [Google Scholar] [CrossRef]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romaniello, R.; Condelli, N.; Romano, P. Selected Indigenous Saccharomyces cerevisiae Strains as Profitable Strategy to Preserve Typical Traits of Primitivo Wine. Fermentation 2019, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Grieco, F.; Tristezza, M.; Vetrano, C.; Bleve, G.; Panico, E.; Mita, G.; Logrieco, A. Exploitation of autochthonous micro-organism potential to enhance the quality of Apulian wines. Annal. Microbiol. 2011, 61, 67–73. [Google Scholar] [CrossRef]
- Capozzi, V.; Garofalo, C.; Chiriatti, M.A.; Grieco, F.; Spano, G. Microbial terroir and food innovation: The case of yeast biodiversity in wine. Microbiol. Res. 2015, 181, 75–83. [Google Scholar] [CrossRef]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Capece, A.; Granchi, L.; Guerrini, S.; Mangani, S.; Romaniello, R.; Vincenzini, M.; Romano, P. Diversity of Saccharomyces cerevisiae strains isolated from two Italian wine-producing regions. Front. Microbiol. 2016, 7, 1018. [Google Scholar] [CrossRef] [Green Version]
- Csoma, H.; Zakany, N.; Capece, A.; Romano, P.; Sipiczki, M. Biological diversity of Saccharomyces yeasts of spontaneously fermenting wines in four wine regions: Comparative genotypic and phenotypic analysis. Int. J. Food Microbiol. 2010, 140, 239–248. [Google Scholar] [CrossRef]
- Feghali, N.; Albertin, W.; Tabet, E.; Rizk, Z.; Bianco, A.; Zara, G.; Masneuf-Pomarede, I.; Budroni, M. Genetic and Phenotypic Characterisation of a Saccharomyces cerevisiae Population of ‘Merwah’ White Wine. Microorganisms 2019, 7, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legras, J.-L.; Karst, F. Optimisation of Inter-Deltas analysis for Saccharomyces cerevisiae strain characterisation. FEMS Microbiol. Lett. 2003, 221, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Legras, J.-L.; Ruh, O.; Merdinoglu, D.; Karst, F. Selection of hypervariable microsatellite loci for the characterization of Saccharomyces cerevisiae strains. Int. J. Food Microbiol. 2005, 102, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Fell, J.W. The Yeasts: A Taxonomic Study; Elsevier Science: Amsterdam, The Netherlands, 1998; p. 1076. [Google Scholar]
- Nguyen, H.V.; Gaillardin, C. Two subgroups within the Saccharomyces bayanus species evidenced by PCR amplification and restriction polymorphism of the non-transcribed spacer 2 in the ribosomal DNA unit. Syst. Appl. Microbiol. 1997, 20, 286–294. [Google Scholar] [CrossRef]
- Hoffman, C.S. Preparation of Yeast DNA. Curr. Protoc. Mol. Biol. 2001, 39, 11–14. [Google Scholar] [CrossRef]
- PHYLIP Home Page—University of Washington. Available online: https://evolution.genetics.washington.edu/phylip.html (accessed on 13 May 2020).
- Jiranek, V.; Langridge, P.; Henschke, A. Validation of Bismuth containing indicator media for predicting H2S production potential of Saccharomyces cerevisiae wine yeasts under oenological conditions. Am. J. Enol. Vitic. 1995, 46, 269–273. [Google Scholar]
- OIV. Guidelines for the Characterization of Wine Yeasts of the Genus Saccharomyces Isolated from Vitivinicultural Environments. Resolution OIV-OENO 370. 2012. Available online: https://www.oiv.int/public/medias/1429/oiv-oeno-370-2012-en.pdf (accessed on 10 May 2021).
- Weast, R.C. Concentrative Properties of Aqueous Solutions: Conversion Tables, Table 88. In Handbook of Chemistry and Physics, 69th ed.; CRC Press: Boca Raton, FL, USA, 1988. [Google Scholar]
- ITV. Choix et Emploi des Microorganismes en Oenologie; ITV Services Publication: Paris, France, 2003; p. 422. [Google Scholar]
- OIV. International Standard for the Labelling of Wines. Edition 2015. Available online: https://www.oiv.int/public/medias/4776/oiv-wine-labelling-standard-en-2015.pdf (accessed on 10 May 2021).
- Siebert, T.; Bramley, B.; Solomon, M. Hydrogen sulfide: Aroma detection threshold study in white and red wines. Technol. Rev. 2009, 183, 14–16. [Google Scholar]
- Rauhut, D. Yeast production of sulfur compounds. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers: Chur, Switzerland, 1993; pp. 183–223. [Google Scholar]
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavour. Adv. Appl. Microbiol. 2005, 57, 131–175. [Google Scholar] [CrossRef]
- Liu, S.Q.; Pilone, G.J. An overview of formation and roles of acetaldehyde in winemaking with emphasis on microbiological implications. Int. J. Food Sci. Technol. 2000, 35, 49–61. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P. Influence de la nature de la souche de levure sur l’arôme des vins. C. R. L’acad. Agric. Fr. 1993, 79, 73–80. [Google Scholar]
- Rojas, V.; Gil, J.V.; Pinaga, J.; Manzanares, P. Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int. J. Food Microbiol. 2003, 86, 181–188. [Google Scholar] [CrossRef]
- Gardner, N.; Rodrigue, N.; Champagne, C. Combined effects of sulfites, temperature and agitation time on production of glycerol in grape juice by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1993, 59, 2022–2028. [Google Scholar] [CrossRef] [Green Version]
- Fleet, G. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Van der Westhuizen, T.J.; Augustyn, O.H.P.; Pretorius, I.S. Geographical distribution of indigenous Saccharomyces cerevisiae strains isolated from vineyards in the coastal regions of the Western Cape in South Africa. S. Afr. J. Enol. Vitic. 2000, 21, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Povhe Jemec, K.; Cadez, N.; Zagorc, T.; Bubic, V.; Zupec, A.; Raspor, P. Yeast population dynamics in five spontaneous fermentations of Malvasia must. Food Microbiol. 2001, 18, 247–259. [Google Scholar] [CrossRef]
- Clemente-Jimenez, J.M.; Mingorance-Cazorla, L.; Martínez-Rodríguez, S.; Javier Las Heras-Vázquez, F.; Rodríguez-Vico, F. Molecular characterization and oenological properties of wine yeasts isolated during spontaneous fermentation of six varieties of grape must. Food Microbiol. 2004, 21, 149–155. [Google Scholar] [CrossRef]
- Capece, A.; Siesto, G.; Romaniello, R.; Lagreca, V.M.; Pietrafesa, R.; Calabretti, A.; Romano, P. Assessment of competition in wine fermentation among wild Saccharomyces cerevisiae strains isolated from Sangiovese grapes in Tuscany region. LWT Food Sci. Technol. 2013, 54, 485–492. [Google Scholar] [CrossRef]
- Cheng, E.; Martiniuk, J.T.; Hamilton, J.; McCarthy, G.; Castellarin, S.D.; Measday, V. Characterization of Sub-Regional Varition in Saccharomyces Populations and Grape Phenolic Composition in Pinot Noir Vineyards of a Canadian Wine Region. Front. Genet. 2020, 11, 908. [Google Scholar] [CrossRef]
- Martiniuk, J.T.; Pacheco, B.; Russell, G.; Tong, S.; Backstrom, I.; Measday, V. Impact of commercial strain use on Saccharomyces cerevisiae population structure and dynamics in pinot noir vineyards and spontaneous fermentations of a Canadian winery. PLoS ONE 2016, 11, e0160259. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Ciani, M. Survival of inoculated Saccharomyces cerevisiae strain on wine grapes during two vintages. Lett. Appl. Microbiol. 2006, 42, 248–253. [Google Scholar] [CrossRef]
- Mercado, L.; Dalcero, A.; Masuelli, R.; Combina, M. Diversity of Saccharomyces strains on grapes and winery surfaces: Analysis of their contribution to fermentative flora of Malbec wine from Mendoza (Argentina) during two consecutive years. Food Microbiol. 2007, 24, 403–412. [Google Scholar] [CrossRef]
- Börlin, M. Diversité et Structure de Population des Levures Saccharomyces cerevisiae à L’échelle du Vignoble Bordelais: Impact de Différents Facteurs sur la Diversité. Ph.D. Thesis, Université de Bordeaux, Bordeaux, France, 2015. [Google Scholar]
- Redzepovic, S.; Orlic, S.; Sikora, S.; Majdak, A.; Pretorius, I.S. Identification and characterization of Saccharomyces cerevisiae and Saccharomyces paradoxus strains isolated from Croatian vineyards. Lett. Appl. Microbiol. 2002, 35, 305–310. [Google Scholar] [CrossRef]
- Sabate, J.; Cano, J.; Esteve-Zarzoso, B.; Guillamon, J.M. Isolation and identification of yeasts associated with vineyard and winery by RFLP analysis of ribosomal genes and mitochondrial DNA. Microbiol. Res. 2002, 157, 267–274. [Google Scholar] [CrossRef]
- Abdo, H.; Catacchio, C.; Ventura, M.; D’Addabbo, P.; Alexandre, H.; Guilloux-Bénatier, M.; Rousseaux, S. The establishment of a fungal consortium in a new winery. Sci. Rep. 2020, 10, 7962. [Google Scholar] [CrossRef]
- Goddard, M.R.; Anfang, N.; Tang, R.; Gardner, R.C.; Jun, C. A distinct population of Saccharomyces cerevisiae in New Zealand: Evidence for local dispersal by insects and human aided global dispersal in oak barrels. Environ. Microbiol. 2010, 12, 63–73. [Google Scholar] [CrossRef]
- Stefanini, I.; Dapporto, L.; Berná, L.; Polsinelli, M.; Turillazzi, S.; Cavalieri, D. Social wasps are a Saccharomyces mating nest. Proc. Natl. Acad. Sci. USA 2016, 113, 2247–2251. [Google Scholar] [CrossRef] [Green Version]
- Francesca, N.; Canale, D.E.; Settanni, L.; Moschetti, G. Dissemination of wine-related yeasts by migratory birds. Environ. Microbiol. Rep. 2012, 4, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Fay, J.; Benavides, J. Evidence for Domesticated and Wild Populations of Saccharomyces cerevisiae. PLoS Genet. 2005, 1, e5. [Google Scholar] [CrossRef]
- Ayoub, M.-J.; Legras, J.-L.; Saliba, R.; Gaillardin, C. Application of Multi Locus Sequence Typing to the analysis of the biodiversity of indigenous Saccharomyces cerevisiae wine yeasts from Lebanon. J. Appl. Microbiol. 2006, 100, 699–711. [Google Scholar] [CrossRef]
- Majdak, A.; Herjavec, S.; Orlic, S.; Redzepovic, S.; Nikola Mirozevic, N. Comparison of Wine Aroma Compounds Produced by Saccharomyces paradoxus and Saccharomyces cerevisiae Strains. Food Technol. Biotechnol. 2002, 40, 103–109. [Google Scholar]
- Comi, G.; Maifreni, M.; Manzano, M.; Lagazio, C.; Cocolin, L. Mitochondrial DNA restriction enzyme analysis and evaluation of the enological characteristics of Saccharomyces cerevisiae strains isolated from grapes of the wine-producing area of Collio (Italy). Int. J. Food Microbiol. 2000, 58, 117–121. [Google Scholar] [CrossRef]
- Blanco, P.; Miras-Avalos, J.M.; Pereira, E.; Orriols, I. Fermentative aroma compounds and sensory profiles of Godello and Albarino wines as influenced by Saccharomyces cerevisiae yeast strains. J. Sci. Food Agric. 2013, 93, 2849–2857. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inter-Diversity Aspect | Number of Years | Number of Villages | Number of Wineries | Number of Vats per Winery and per Year | Number of Isolates per Winery and per Year | Winery Name | Year Considered |
|---|---|---|---|---|---|---|---|
| Biogeographical diversity survey | 4 | 51 | 79 | 1 to 6 | 1 to 14 | See Figure S1 | Years I, II, III, IV |
| Diversity within wineries ‡ | 2 | 9 | 9 * | 1 | 9 or 10 | Winery M | Years III and IV |
| Winery MC | Year IV | ||||||
| Winery WKa | Year IV | ||||||
| Winery A | Year IV | ||||||
| Winery T | Year IV | ||||||
| Winery B | Years III and IV | ||||||
| Winery HY | Years III and IV | ||||||
| Winery DG | Year III | ||||||
| Winery D | Year III | ||||||
| Perenniality in wineries ‡ | 2 to 4 | 7 | 7 * | 1 to 6 | 4 to 14 ** | Winery M | Years I, II, III, IV |
| Winery MC | Years I, III, IV | ||||||
| Winery WKa | Years III and IV | ||||||
| Winery A | Years III and IV | ||||||
| Winery T | Years III and IV | ||||||
| Winery B | Years III and IV | ||||||
| Winery HY | Years III and IV |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayoub, M.-J.; Legras, J.-L.; Abi-Nakhoul, P.; Nguyen, H.-V.; Saliba, R.; Gaillardin, C. Lebanon’s Native Oenological Saccharomyces cerevisiae Flora: Assessment of Different Aspects of Genetic Diversity and Evaluation of Winemaking Potential. J. Fungi 2021, 7, 678. https://doi.org/10.3390/jof7080678
Ayoub M-J, Legras J-L, Abi-Nakhoul P, Nguyen H-V, Saliba R, Gaillardin C. Lebanon’s Native Oenological Saccharomyces cerevisiae Flora: Assessment of Different Aspects of Genetic Diversity and Evaluation of Winemaking Potential. Journal of Fungi. 2021; 7(8):678. https://doi.org/10.3390/jof7080678
Chicago/Turabian StyleAyoub, Marie-José, Jean-Luc Legras, Pierre Abi-Nakhoul, Huu-Vang Nguyen, Rachad Saliba, and Claude Gaillardin. 2021. "Lebanon’s Native Oenological Saccharomyces cerevisiae Flora: Assessment of Different Aspects of Genetic Diversity and Evaluation of Winemaking Potential" Journal of Fungi 7, no. 8: 678. https://doi.org/10.3390/jof7080678