
The Culturable Mycobiome of Mesophotic Agelas oroides: Constituents and Changes Following Sponge Transplantation to Shallow Water


Abstract
:1. Introduction
2. Materials and Methods
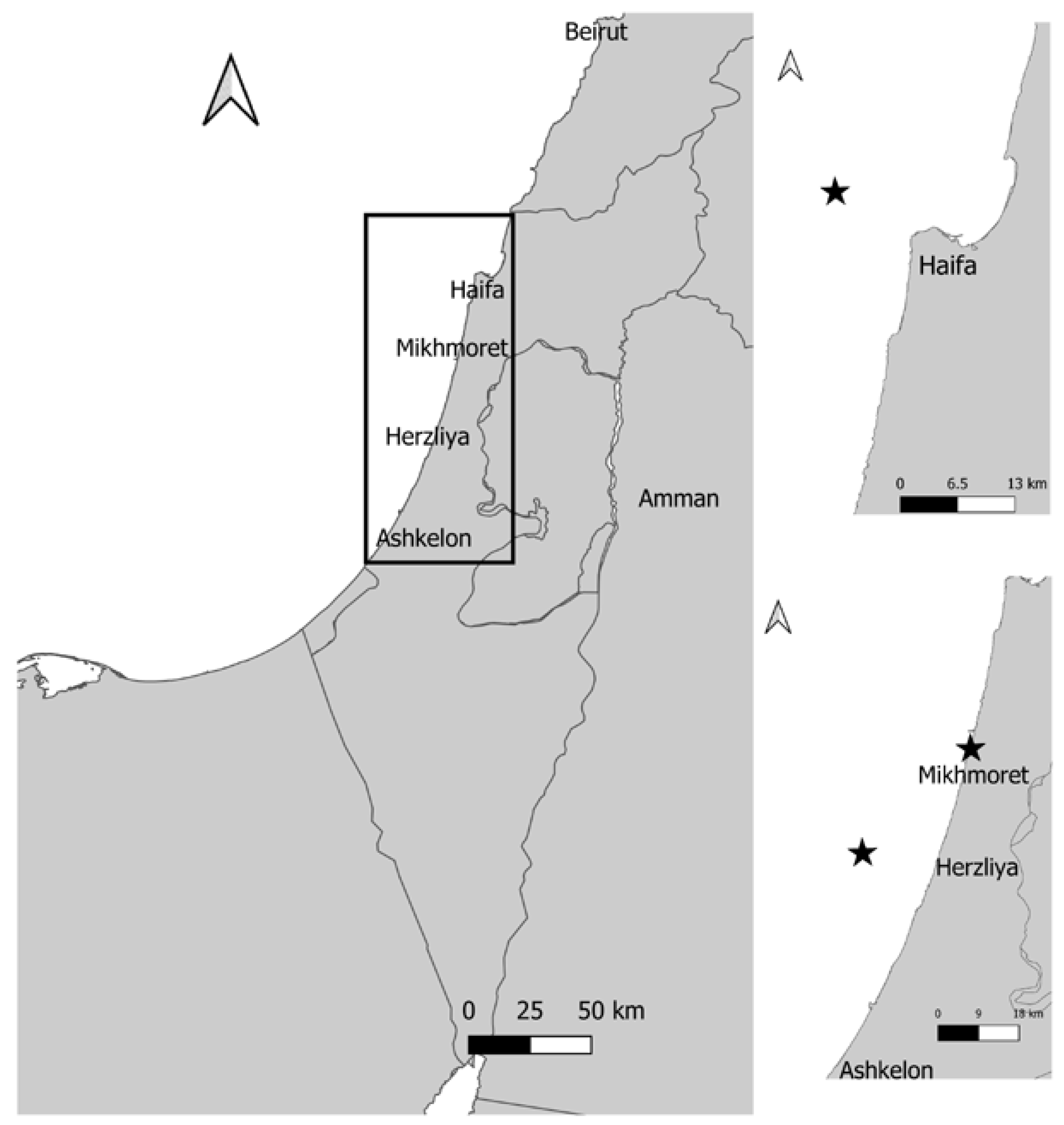
2.1. Sponge Sample Collection
2.2. Isolation and Identification of Agelas oroides-Derived Fungi
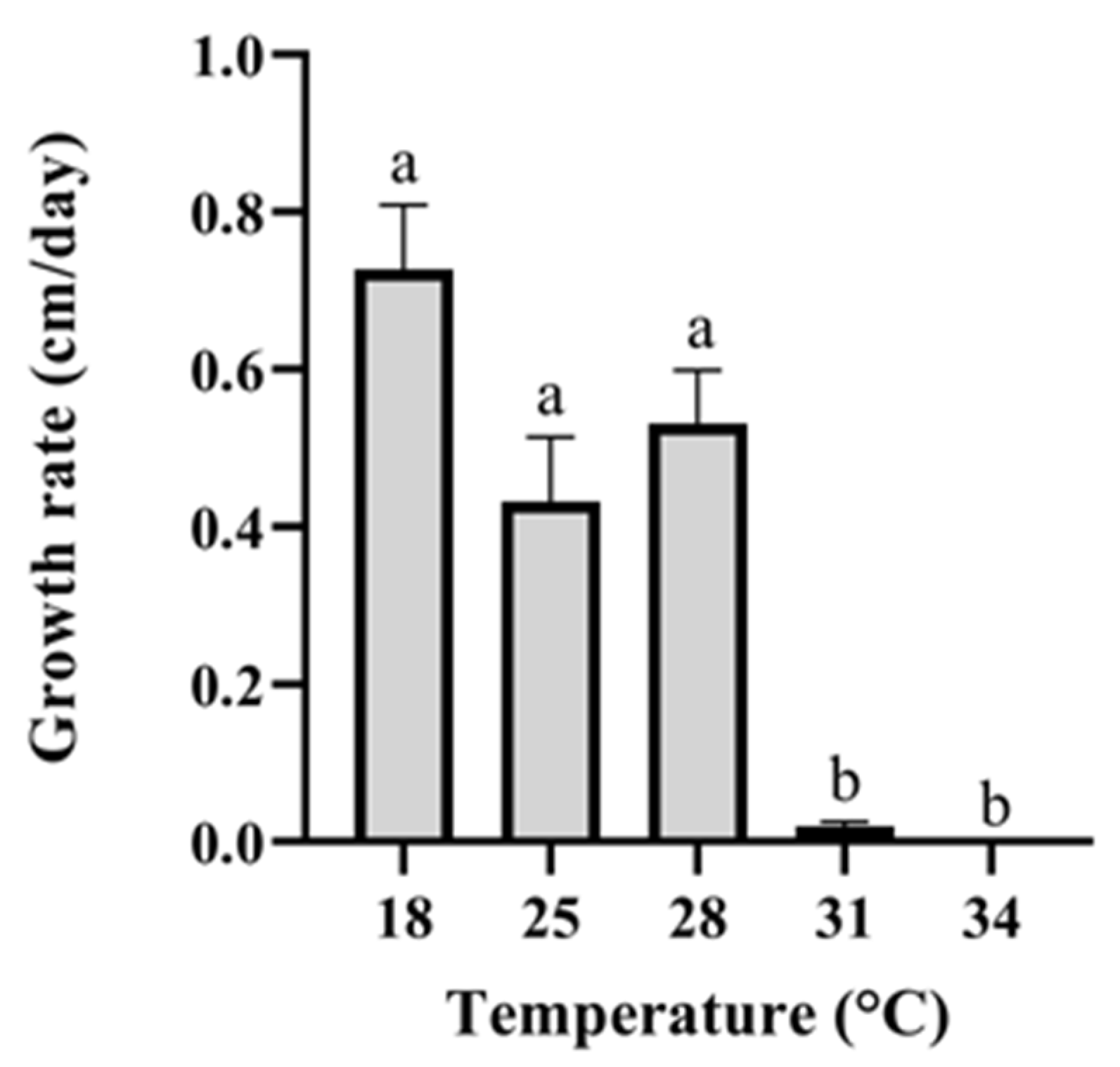
2.3. Fungal-Fungal Interactions and Assays of Growth on Sponge Tissue
3. Results
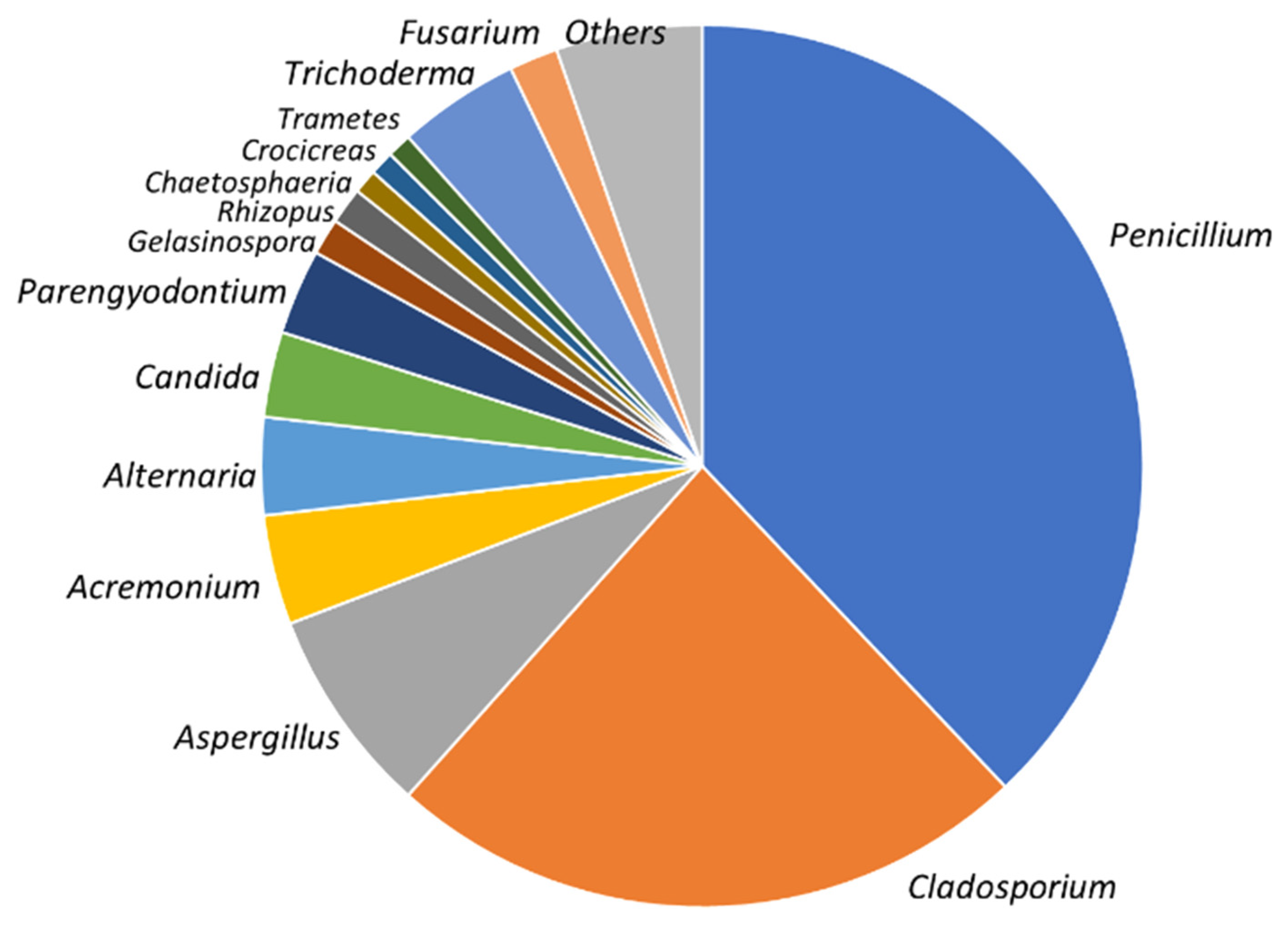
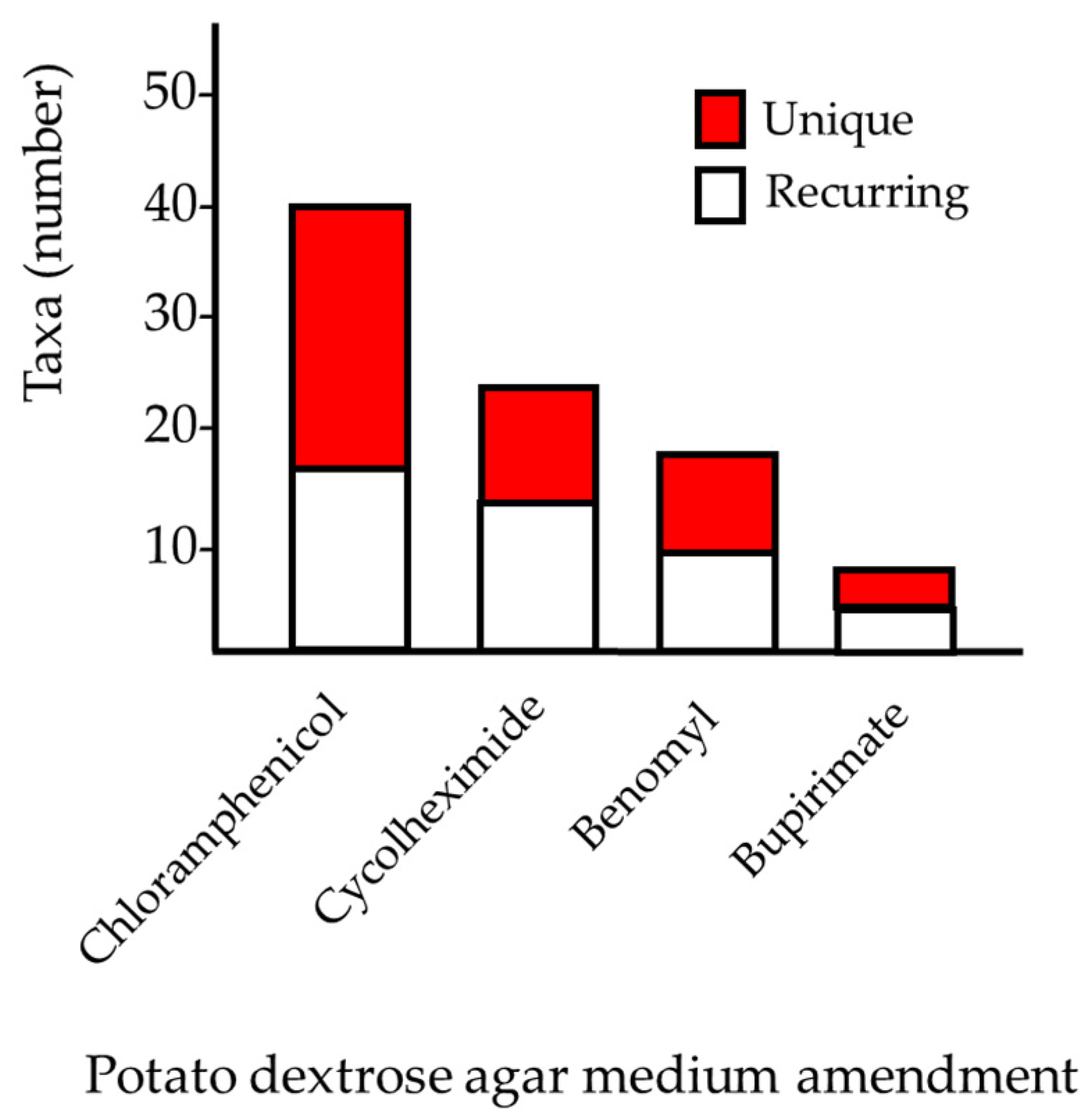
3.1. Fungal Diversity in Agelas oroides
3.2. Members of the Agelas oroides Mycobiome Exhibit a Variety of Fungus-Fungus Interactions
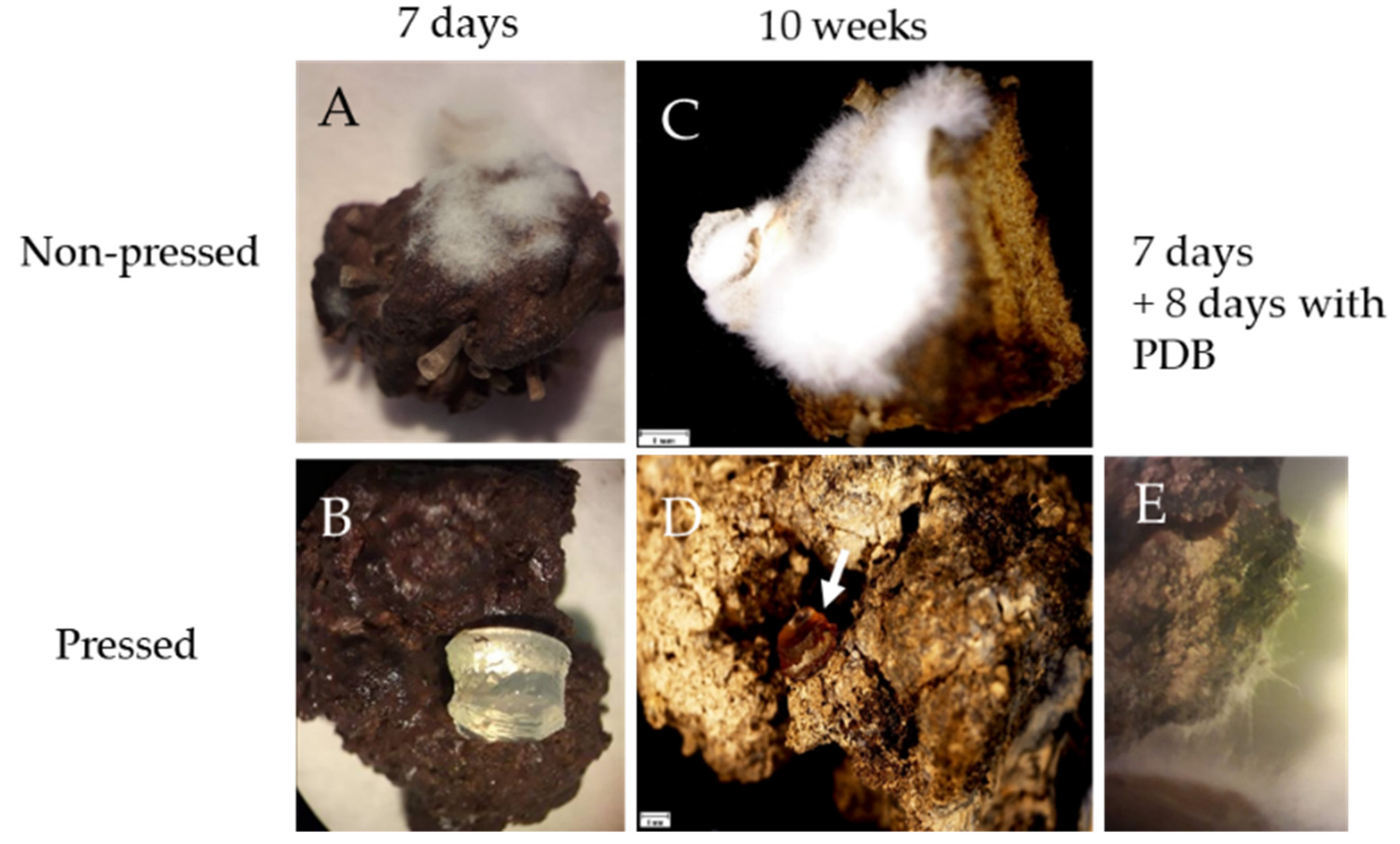
3.3. Sponge Holo-Biont Constituents Can Support Growth of Parengyodontium album
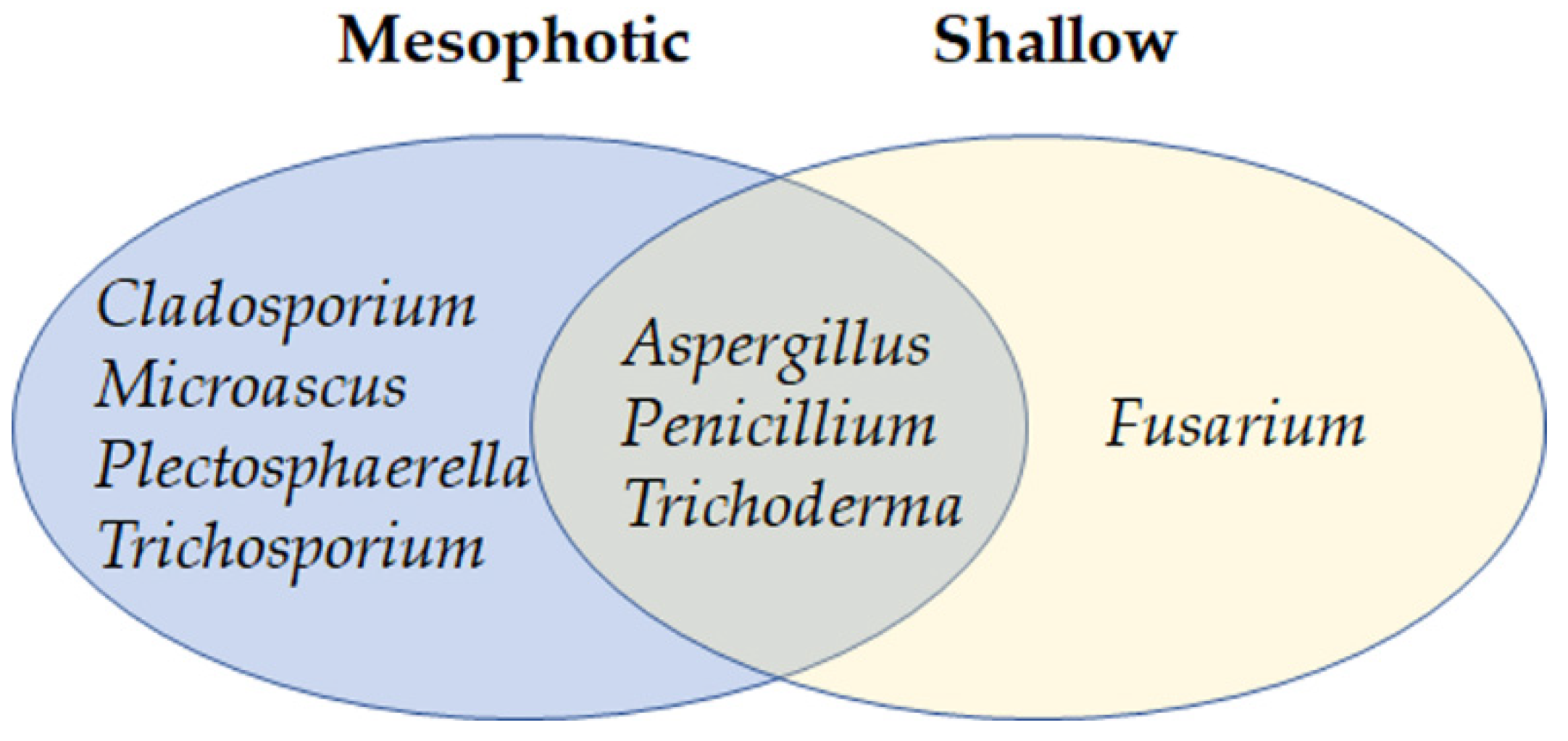
3.4. Transfer of Agelas oroides from Mesophotic to Shallow Water Confers Changes in the Mycobiome
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Soest, R.W.M.; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez, B.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schönberg, C.; et al. World Porifera Database. Available online: www.marinespecies.org/porifera (accessed on 8 March 2021).
- Hickman, C.P.; Roberts, L.S.; Keen, S.L.; Larson, A.; L’Anson, H.; Eisenhour, D.J. Integrated Principles of Zoology; McGraw-Hill: New York, NY, USA, 2008; pp. 246–259. [Google Scholar]
- Pita, L.; Rix, L.; Slaby, B.M.; Franke, A.; Hentschel, U. The Sponge Holobiont in a Changing Ocean: From Microbes to Ecosystems. Microbiome 2018, 6, 46. [Google Scholar] [CrossRef]
- Beazley, L.I.; Kenchington, E.L.; Murillo, F.J.; Sacau, M.D.M. Deep-Sea Sponge Grounds Enhance Diversity and Abundance of Epibenthic Megafauna in the Northwest Atlantic. ICES J. Mar. Sci. 2013, 70, 1471–1490. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Chintiroglou, C.C.; Konstantinou, D.; Voultsiadou, E. Sponges as “Living Hotels” in Mediterranean Marine Caves. Sci. Mar. 2016, 80, 279–289. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative Stress in Marine Environments: Biochemistry and Physiological Ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoukatou, M.; Hellio, C.; Vagias, C.; Harvala, C.; Roussis, V. Chemical Defense and Antifouling Activity of Three Mediterranean Sponges. Z. Naturforsch. 2014, 57, 161–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.; Hill, R.; Piel, J.; Thacker, R.; Hentschel, U. Soaking it Up: The Complex Lives of Marine Sponges and Their Microbial Associates. ISME J. 2007, 1, 187–190. [Google Scholar] [CrossRef]
- Taylor, M.; Radax, R.; Steger, D.; Wagner, M. Sponge-Associated Microorganisms: Evolution, Ecology and Biotechnological Potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [CrossRef] [Green Version]
- Cohen, E.; Koch, L.; Myint, T.K.; Rahamim, Y.; Aluma, Y.; Ilan, M.; Yarden, O.; Carmeli, S. Novel Terpenoids of the Fungus Aspergillus insuetus Isolated from the Mediterranean Sponge Psammocinia sp. Collected along the Coast of Israel. Bioorg. Med. Chem. 2011, 19, 6587–6593. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.M.; Tang, X.L.; Sun, Y.T.; Song, D.Y.; Cheng, Y.J.; Liu, H.; Li, P.L.; Li, G.Q. Biological and Chemical Diversity of Marine Sponge-Derived Microorganisms over the Last Two Decades from 1998 to 2017. Molecules 2020, 25, 853. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, S.; Tsai, P.; Bell, J.; Fromont, J.; Ilan, M.; Lindquist, N.; Perez, T.; Rodrigo, A.; Schupp, P.J.; Vacelet, J.; et al. Assessing the Complex Sponge Microbiota: Core, Variable and Species-Specific. ISME J. 2012, 6, 564–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.H.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; et al. Fungi in the Marine Environment: Open Questions and Unsolved Problems. MBio 2019, 10, e01189-18. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.T.H.D.; Thomas, T. Diversity, Host-Specificity and Stability of Sponge-Associated Fungal Communities of Co-Occurring Sponges. PeerJ 2018, 6, e4965. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Li, B.; Zheng, C.; Wang, G. Molecular Detection of Fungal Communities in the Hawaiian Marine Sponges Suberites zeteki and Mycale armata. Appl. Environ. Microbiol. 2008, 74, 6091–6101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naim, M.A.; Smidt, H.; Sipkema, D. Fungi Found in Mediterranean and North Sea Sponges: How Specific are They? PeerJ 2017, 5, e3722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz, Z.; Komon-Zelazowska, M.; Druzhinina, I.S.; Aveskamp, M.M.; Shnaiderman, A.; Aluma, Y.; Carmeli, S.; Ilan, M.; Yarden, O. Diversity and Potential Antifungal Properties of Fungi Associated with a Mediterranean Sponge. Fungal Divers. 2010, 42, 17–26. [Google Scholar] [CrossRef]
- Yarden, O. Fungal Association with Sessile Marine Invertebrates. Front. Microbiol. 2014, 5, 1–6. [Google Scholar] [CrossRef]
- Easson, C.G.; Chaves-Fonnegra, A.; Thacker, R.W.; Lopez, J.V. Host Population Genetics and Biogeography Structure the Microbiome of the Sponge Cliona Delitrix. Ecol Evol. 2020, 10, 2007–2020. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, S.M.; Antwis, R.E.; Lenzi, L.; Lucaci, A.; Behringer, D.C.; Butler, M.J.; Preziosi, R.F. Host Genetics and Geography Influence Microbiome Composition in the Sponge Ircinia Campana. J. Anim. Ecol. 2019, 88, 1684–1695. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, C.; Marengo, B.; De Ciucis, C.; Nitti, M.; Pronzato, M.; Marinari, U.; Pronzato, R.; Manconi, R.; Domenicotti, C. Effects of Agelas oroides and Petrosia ficiformis Crude Extracts on Human Neuroblastoma Cell Survival. Int. J. Oncol. 2007, 30, 161–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferretti, C.; Vacca, S.; De Ciucis, C.; Marengo, B.; Duckworth, A.R.; Manconi, R.; Pronzato, R.; Domenicotti, C. Growth Dynamics and Bioactivity Variation of the Mediterranean Demosponge Agelas oroides (Agelasida, Agelasidae) and Petrosia ficiformis (Haplosclerida, Petrosiidae). Mar. Ecol. 2009, 30, 327–336. [Google Scholar] [CrossRef]
- Idan, T.; Shefer, S.; Feldstein, T.; Yahel, R.; Huchon, D.; Ilan, M. Shedding Light on an East-Mediterranean Mesophotic Sponge Ground Community and the Regional Sponge Fauna. Medit. Mar. Sci. 2018, 19, 84–106. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Zapata, F.L.; Bongaerts, P.; Ramírez-Portilla, C.; Adu-Oppong, B.; Walljasper, G.; Reyes, A.; Sanchez, J.A. Holobiont Diversity in a Reef-Building Coral Over its Entire Depth Range in the Mesophotic Zone. Front. Mar. Sci. 2018, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Kahng, S.E.; Copus, J.M.; Wagner, D. Recent Advances in the Ecology of Mesophotic Coral Ecosystems (MCEs). Curr. Opin. Environ. Sustain. 2014, 7, 72–81. [Google Scholar] [CrossRef]
- Lesser, M.P.; Slattery, M.; Leichter, J.J. Ecology of Mesophotic Coral Reefs. J. Exp. Mar. Biol. Ecol. 2009, 375, 1–8. [Google Scholar] [CrossRef]
- Bongaerts, P.; Riginos, C.; Brunner, R.; Englebert, N.; Smith, S.R.; Hoegh-Guldberg, O. Deep Reefs Are Not Universal Refuges: Reseeding Potential Varies Among Coral Species. Sci. Adv. 2017, 3, e1602373. [Google Scholar] [CrossRef] [Green Version]
- Tsurnamal, M. Studies on the Porifera of the Mediterranean Littoral of Israel. Ph.D. Thesis, The Hebrew University of Jerusalem, Jerusalem, Israel, 1968. [Google Scholar]
- Idan, T.; Goren, L.; Shefer, S.; Ilan, M. Sponges in a Changing Climate: Survival of Agelas oroides in a Warming Mediterranean Sea. Front. Mar. Sci. 2020, 7, 1–11. [Google Scholar] [CrossRef]
- Gloeckner, V.; Wehrl, M.; Moitinho-Silva, L.; Gernert, C.; Schupp, P.; Pawlik, J.R.; Lindquist, N.L.; Erpenbeck, D.; Wörheide, G.; Hentschel, U. The HMA-LMA Dichotomy Revisited: An Electron Microscopical Survey of 56 Sponge Species. Biol. Bull. 2005, 277, 78–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poppell, E.; Weisz, J.; Spicer, L.; Massaro, A.; Hill, A.; Hill, M. Sponge Heterotrophic Capacity and Bacterial Community Structure in High- and Low-Microbial Abundance Sponges. Mar. Ecol. 2013, 35, 414–424. [Google Scholar] [CrossRef]
- Abdel-Wahab, N.M.; Scharf, S.; Özkaya, F.C.; Kurtán, T.; Mándi, A.; Fouad, M.A.; Kamel, M.S.; Müller, W.E.G.; Kalscheuer, R.; Lin, W.; et al. Induction of Secondary Metabolites from the Marine-Derived Fungus Aspergillus Versicolor through Co-Cultivation with Bacillus subtilis. Planta Med. 2019, 85, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Frank, M.; Özkaya, F.C.; Müller, W.E.G.; Hamacher, A.; Kassack, M.U.; Lin, W.H.; Liu, Z.; Proksch, P. Cryptic Secondary Metabolites from the Sponge-Associated Fungus Aspergillus Ochraceus. Mar. Drugs. 2019, 17, 99. [Google Scholar] [CrossRef] [Green Version]
- Bell, T.G.; Kramvis, A. Fragment Merger: An Online Tool to Merge Overlapping Long Sequence Fragments. Viruses 2013, 5, 824–833. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Banerjee, D. Proteases from Endophytic Fungi with Potential Industrial Applications. In Recent Advancement in White Biotechnology through Fungi. Fungal Biology; Yadav, A., Mishra, S., Singh, S., Gupta, A., Eds.; Springer: Cham, Switzerland, 2019; pp. 319–359. ISBN 978-3-030-10480-1. [Google Scholar]
- Le Goeff, G.; Lopes, P.; Arcile, G.; Vlachou, P.; Van Elslande, E.; Retailleau, P.; Gallard, J.F.; Weis, M.; Benayahu, Y.; Fokialakis, N.; et al. Impact of the Cultivation Technique on the Production of Secondary Metabolites by Chrysosporium Lobatum TM-237-S5, Isolated from the Sponge Acanthella Cavernosa. Mar. Drugs. 2019, 17, 678. [Google Scholar] [CrossRef] [Green Version]
- Frank, M.; Hartmann, R.; Plenker, M.; Mandi, A.; Kurtan, T.; Ozkaya, F.C.; Muller, W.E.G.; Kassack, M.U.; Hamacher, A.; Lin, W.H.; et al. Azaphilones from the Sponge-Associated Fungus Penicillium Canescens Strain 4.14.6a. J. Nat. Prod. 2019, 82, 2159–2166. [Google Scholar] [CrossRef] [PubMed]
- Kavruk, E.; Karabey, B.; Temel, H.Y.; Tuna, E.E.H. Phylogenetic Diversity of Mediterranean Sponge Microbiome Using Denaturing Gradient Gel Electrophoresis. Fres. Environ. Bull. 2019, 28, 10011–10020. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.B.G.; Suetrong, S.; Sakayaroj, J.; Bahkali, A.H.; Abdel-Wahab, M.A.; Boekhout, T.; Pang, K.L. Classification of Marine Ascomycota, Basidiomycota, Blastocladiomycota and Chytridiomycota. Fungal Divers. 2015, 73, 1–72. [Google Scholar] [CrossRef]
- Orellana, S.C. Assessment of Fungal Diversity in the Environment Using Metagenomics: A Decade in Review. Fungal Genom. Biol. 2013, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Wang, N.F.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and Distribution of Fungal Communities in the Marine Sediments of Kongsfjorden, Svalbard (High Arctic). Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Passarini, M.R.Z.; Santos, C.; Lima, N.; Berlinck, R.G.S.; Sette, L.D. Filamentous Fungi from the Atlantic Marine Sponge Dragmacidon Reticulatum. Arch. Microbiol. 2013, 195, 99–111. [Google Scholar] [CrossRef]
- Du Plessis, I.L.; Druzhinina, I.S.; Atanasova, L.; Yarden, O.; Jacobs, K. The Diversity of Trichoderma Species from Soil in South Africa, with Five New Additions. Mycologia 2018, 110, 559–583. [Google Scholar] [CrossRef]
- Gal-Hemed, I.; Atanasova, L.; Komon-Zelazowska, M.; Druzhinina, I.S.; Viterbo, A.; Yarden, O. Marine Isolates of Trichoderma spp. as Potential Halotolerant Agents of Biological Control for Arid-Zone Agriculture. Appl. Environ. Microbiol. 2011, 77, 5100–5109. [Google Scholar] [CrossRef] [Green Version]
- Gesner, S.; Cohen, N.; Ilan, M.; Yarden, O.; Carmeli, S. Pandangolide 1a, a New Metabolite of the Sponge-Associated Fungus Cladosporium sp. and the Absolute Stereochemistry of Pandangolide 1 and iso-Cladospolide B. J. Nat. Prod. 2005, 68, 1350–1353. [Google Scholar] [CrossRef] [PubMed]
- Panizel, I.; Yarden, O.; Ilan, M.; Carmeli, S. Eight Novel Peptaibols from Sponge-Associated Trichoderma Atroviride. Mar. Drugs 2013, 11, 4937–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, L.; Lodin, A.; Herold, I.; Ilan, M.; Carmeli, S.; Yarden, O. Sensitivity of Neurospora Crassa to a Marine-Derived Aspergillus Tubingensis Anhydride Exhibiting Antifungal Activity that Is Mediated by the MAS1 Protein. Mar. Drugs 2014, 12, 4713–4731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sempere, F.; Santamarina, M.P. Suppression of Nigrospora Oryzae (Berk. & Broome) Petch by an Aggressive Mycoparasite and Competitor, Penicillium Oxalicum Currie & Thom. Int. J. Food Microbiol. 2008, 122, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Horn, B.W.; Peterson, S.W. Host Specificity of Eupenicillium Ochrosalmoneum, E. Cinnamopurpureum and Two Penicillium Species Associated with the Conidial Heads of Aspergillus. Mycologia 2008, 100, 12–19. [Google Scholar] [CrossRef]
- Hu, X.J.; Qin, L.; Roberts, D.P.; Lakshman, D.K.; Gong, Y.M.; Maul, J.E.; Xie, L.H.; Yu, C.B.; Li, Y.S.; Hu, L.; et al. Characterization of Mechanisms Underlying Degradation of Sclerotia of Sclerotinia Sclerotiorum by Aspergillus Aculeatus Asp-4 Using a Combined qRT-PCR and Proteomic Approach. BMC Genom. 2017, 18, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Todd, J.; Ward, T.J.; Robert, V.A.R.G.; Crous, P.W.; Geiser, D.M.; Kang, S. DNA Sequence-Based Identification of Fusarium: Current Status and Future Directions. Phytoparasitica 2015, 43, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Frisvad Jens, C.; Larsen Thomas, O. Extrolites of Aspergillus Fumigatus and Other Pathogenic Species in Aspergillus Section Fumigati. Front. Microbiol. 2016, 6, 1485. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, S.S.; Stchigel, A.M.; Cano, J.F.; Godoy-Martinez, P.C.; Colombo, A.L.; Guarro, J. Aspergillus Novoparasiticus: A New Clinical Species of the Section Flavi. Med. Mycol. 2012, 50, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Gourama, H.; Bullerman, L.B. Aspergillus flavus and Aspergillus Parasiticus: Aflatoxigenic Fungi of Concern in Foods and Feeds. J. Food Prot. 1995, 58, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Ein-Gil, N.; Ilan, M.; Carmeli, S.; Smith, G.W.; Pawlik, J.R.; Yarden, O. Presence of Aspergillus Sydowii, a Pathogen of Gorgonian Sea Fans in the Marine Sponge Spongia Obscura. ISME J. 2009, 3, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.G.; Liu, Q.; Zhu, G.L.; Liu, H.S.; Zhu, W.M. Marine Natural Products Sourced from Marine-Derived Penicillium Fungi. J. Asian Nat. Prod. Res. 2016, 18, 92–115. [Google Scholar] [CrossRef]
- Nicoletti, R.; Trincone, A. Bioactive Compounds Produced by Strains of Penicillium and Talaromyces of Marine Origin. Mar. Drugs. 2016, 14, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Famurewa, O.; Olutiola, P.O. Comparison of Growth and Cellulolytic Enzyme Production in Aspergillus Chevalieri and Penicillium Steckii from Mouldy Cacao Beans. Folia Microbiol. 1991, 36, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Lifshitz, N.; Hazanov, L.; Fine, M.; Yarden, O. Seasonal Variations in the Culturable Mycobiome of Acropora Loripes along a Depth Gradient. Microorganisms 2020, 8, 1139. [Google Scholar] [CrossRef]
- Chellappan, S.; Jasmin, C.; Basheer, S.M.; Elyas, K.K.; Bhat, S.G.; Chandrasekaran, M. Production, Purification and Partial Characterization of a Novel Protease from Marine Engyodontium Album BTMFS10 under Solid State Fermentation. Process. Biochem. 2006, 41, 956–961. [Google Scholar] [CrossRef]
- Tsang, C.C.; Chan, J.F.W.; Pong, W.M.; Chen, J.H.K.; Ngan, A.H.Y.; Cheung, M.; Lai, C.K.C.; Tsang, D.N.C.; Lau, S.K.P.; Woo, P.C.Y. Cutaneous Hyalohyphomycosis Due to Parengyodontium Album gen. et comb. nov. Med. Mycol. 2016, 54, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.; Wang, J.; Zhang, X.; Nong, X.; Xu, X.; Qi, S. Cytotoxic Polyketides from the Deep Sea Derived Fungus Engyodontium Album DFFSCS021. Mar. Drugs. 2014, 12, 5902–5915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, P.A.; Webster, N.S.; Miller, D.J.; Bourne, D.G. Host-Microbe Coevolution: Applying Evidence from Model Systems to Complex Marine Invertebrate Holobionts. MBio 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kelly, J.B.; Carlson, D.E.; Low, J.S.; Rice, T.; Thacker, R.W. The Relationship between Microbiomes and Selective Regimes in the Sponge Genus Ircinia. Front. Microbiol. 2021, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kumala, L.; Riisgård, H.U.; Canfield, D.E. Osculum Dynamics and Filtration Activity in Small Single-Osculum Explants of the Demosponge Halichondria Panicea. Mar. Ecol. Prog. Ser. 2017, 572, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, F.; Røy, H.; Bayer, K.; Hentschel, U.; Pfannkuchen, M.; Brümmer, F.; de Beer, D. Oxygen Dynamics and Transport in the Mediterranean Sponge Aplysina Aerophoba. Mar. Biol. 2008, 153, 1257–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavy, A.; Keren, R.; Yahel, G.; Ilan, M. Intermittent Hypoxia and Prolonged Suboxia Measured in situ in a Marine Sponge. Front. Mar. Sci. 2016, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name and Targeted Templates | Primer Designation (Annealing Temp.) | Sequence |
|---|---|---|
| Internal Transcribed Spacer (ITS) (all fungal templates) | ITS-5F ITS-4R (62 °C) | GGAAGTAAAAGTCGTAACAAGG TCCTCCGCTTATTGATATGC |
| ACTINa (ACT) (Cladosporium spp.) | ACT-512F ACT-783R (60 °C) | ATGTGCAAGGCCGGTTTCGC TACGAGTCCTTCTGGCCCAT |
| β-tubulin (Tub2) (Acremonium spp.; Chaetomium sp.; Nigrospora sp.; Parengydontium spp.; Penicillium spp.; Talaromyces sp.; Zygosporium spp.) (benA) (Aspergillus spp.; Penicillium spp.) | Bt2a-F Bt2b-R (68.9 °C) Ben2f Bt2b-R (55 °C) | GGTAACCAAATCGGTGCTGCTTTC ACCCTCAGTGTAGTGACCCTTGGC TCCAGACTGGTCAGTGTGTAA ACCCTCAGTGTAGTGACCCTTGGC |
| Calmodulin (CaM) (Aspergillus spp.) | CMD 5 CMD 6 (55 °C) | CCGAGTACAAGGAGGCCTTC CCGATAGAGGTCATAACGTGG |
| Glyceraldehyde-3-phosphate dehydrogenase (GPD) (Alternaria spp.) | gpd1-F gpd2-R (58 °C) | CAACGGCTTCGGTCGCATTG GCCAAGCAGTTGGTTGTGC |
| RNA polymerase II subunit 2 (RPB2) (Acremonium spp.; Penicillium sp.; Pichia sp.) | fRPB2-5F fRPB2-7cR (58.5 °C) | GATGATAGAGATCATTTTGG ATGGGTAAACAAGCTATGGG |
| Large Subunit (D1/D2) of rRNA (Crocicreas spp.; Arthrographis sp.; Sarocladium sp.) | NL-1F NL-4R (55 °C) | GCATATCAATAAGCGGAGGAAAAG GGTCCGTGTTTCAAGACGG |
| Translation elongation factor 1-alpha (TEF1) (Trichoderma spp.) (Fusarium spp.) | EF1-728F TEF1LLEr (62 °C) ef1 ef2 (55 °C) | CATCGAGAAGTTCGAGAAGG AACTTGCAGGCAATGTGG ATGGGTAAGGARGACAAGAC GGARGTACCAGTSATCATGTT |
| Taxon | Number of Isolated Strains | Genes, in Addition to ITS, Used for Molecular Identification (“-“ Indicates No Additional Genes Were Analyzed) |
|---|---|---|
| Penicillium steckii | 47 | Tub2 |
| Penicillium sp. | 12 | - |
| Cladosporium limoniforme | 10 | ACT |
| Aspergillus sp. | 8 | - |
| Cladosporium halotolerans | 8 | ACT |
| Cladosporium sp. | 8 | - |
| Cladosporium sphaerospermum | 8 | ACT |
| Penicillium brevicompactum | 8 | Tub2 |
| Acremonium sclerotigenum | 7 | RPB2 |
| Candida sp. | 7 | - |
| Parengyodontium album | 7 | Tub2 |
| Cladosporium ramotenellum | 6 | ACT |
| Alternaria alternata | 5 | GPD |
| Aspergillus niger | 5 | Cam |
| Aspergillus flavus | 4 | Cam |
| Penicillium chrysogenum | 4 | Tub2 |
| Penicillium citrinum | 4 | Tub2 |
| Alternaria sp. | 3 | - |
| Aspergillus tubingensis | 3 | Cam |
| Cladosporium aciculare | 3 | ACT |
| Cladosporium perangustum | 3 | ACT |
| Gelasinospora sp. | 3 | - |
| Rhizopus sp. | 3 | - |
| Trichoderma sp. | 3 | - |
| Acremonium sp. | 2 | - |
| Aspergillus protuberus | 2 | Cam |
| Chaetosphaeria sp. | 2 | - |
| Cladosporium dominicanum | 2 | ACT |
| Cladosporium tenellum | 2 | ACT |
| Crocicreas coronatum | 2 | D1/D2 |
| Fusarium acuminatum | 2 | TEF1 |
| Penicillium adametzioides | 2 | Tub2 |
| Penicillium capsulatum | 2 | Tub2 |
| Trametes sp. | 2 | - |
| Trichoderma atroviride | 2 | TEF1 |
| Trichoderma orientale | 2 | TEF1 |
| Arthrographis kalrae | 1 | D1/D2 |
| Aspergillus fumigatiaffinis | 1 | Cam |
| Aspergillus insulicola | 1 | benA |
| Aspergillus sydowii | 1 | Cam |
| Aspergillus novoparasiticus | 1 | Cam |
| Chaetomium longiciliata | 1 | Tub2 |
| Cladosporium aggregatocicatricatum | 1 | ACT |
| Cladosporium angustisporum | 1 | ACT |
| Cladosporium longicatenatum | 1 | ACT |
| Cystidiodontia sp. | 1 | - |
| Exophiala sp. | 1 | - |
| Fusarium brachygibbosum | 1 | TEF1 |
| Fusarium equiseti | 1 | TEF1 |
| Microascus sp. | 1 | - |
| Monocillium sp. | 1 | - |
| Nigrospora osmanthi | 1 | Tub2 |
| Penicillium astrolabium | 1 | benA |
| Penicillium coffeae | 1 | Tub2 |
| Penicillium digitatum | 1 | Tub2 |
| Penicillium simile | 1 | benA |
| Penicillium sizovae | 1 | Tub2 |
| Penicillium wotroi | 1 | Tub2 |
| Pichia guilliermondii | 1 | RPB2 |
| Pichia sp. | 1 | - |
| Plectosphaerella sp. | 1 | - |
| Rhodotorula sp. | 1 | - |
| Sarocladium bacillisporum | 1 | D1/D2 |
| Talaromyces funiculosus | 1 | Tub2 |
| Trichoderma atrobrunneum | 1 | TEF1 |
| Trichoderma gamsii | 1 | TEF1 |
| Trichoderma guizhouense | 1 | TEF1 |
| Zygosporium masonii | 1 | Tub2, D1/D2 |
| Zygosporium pseudogibbum | 1 | Tub2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben-Dor Cohen, E.; Ilan, M.; Yarden, O. The Culturable Mycobiome of Mesophotic Agelas oroides: Constituents and Changes Following Sponge Transplantation to Shallow Water. J. Fungi 2021, 7, 567. https://doi.org/10.3390/jof7070567
Ben-Dor Cohen E, Ilan M, Yarden O. The Culturable Mycobiome of Mesophotic Agelas oroides: Constituents and Changes Following Sponge Transplantation to Shallow Water. Journal of Fungi. 2021; 7(7):567. https://doi.org/10.3390/jof7070567
Chicago/Turabian StyleBen-Dor Cohen, Eyal, Micha Ilan, and Oded Yarden. 2021. "The Culturable Mycobiome of Mesophotic Agelas oroides: Constituents and Changes Following Sponge Transplantation to Shallow Water" Journal of Fungi 7, no. 7: 567. https://doi.org/10.3390/jof7070567