
β-Aminobutyric Acid Induced Resistance against Alternaria Fruit Rot in Apple Fruits



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Pathogen
2.3. In Vitro Assays
2.3.1. Germination
2.3.2. Mycelial Growth
2.3.3. Growth in Liquid Medium
2.4. Efficacy in Detached Fruits
2.4.1. Inoculation Procedure
2.4.2. Experiment 1. Effect of Application Date on Rot Development
2.4.3. Experiment 2. Effect of BABA Concentration on Rot Development
2.4.4. Experiment 3. Efficacy of Isomers of BABA
2.5. Efficacy in the Orchard
2.5.1. Field Experiment 1 and 2
2.5.2. Field Experiment 3
2.6. Data Analysis
3. Results
3.1. In Vitro Assays
3.1.1. Germination
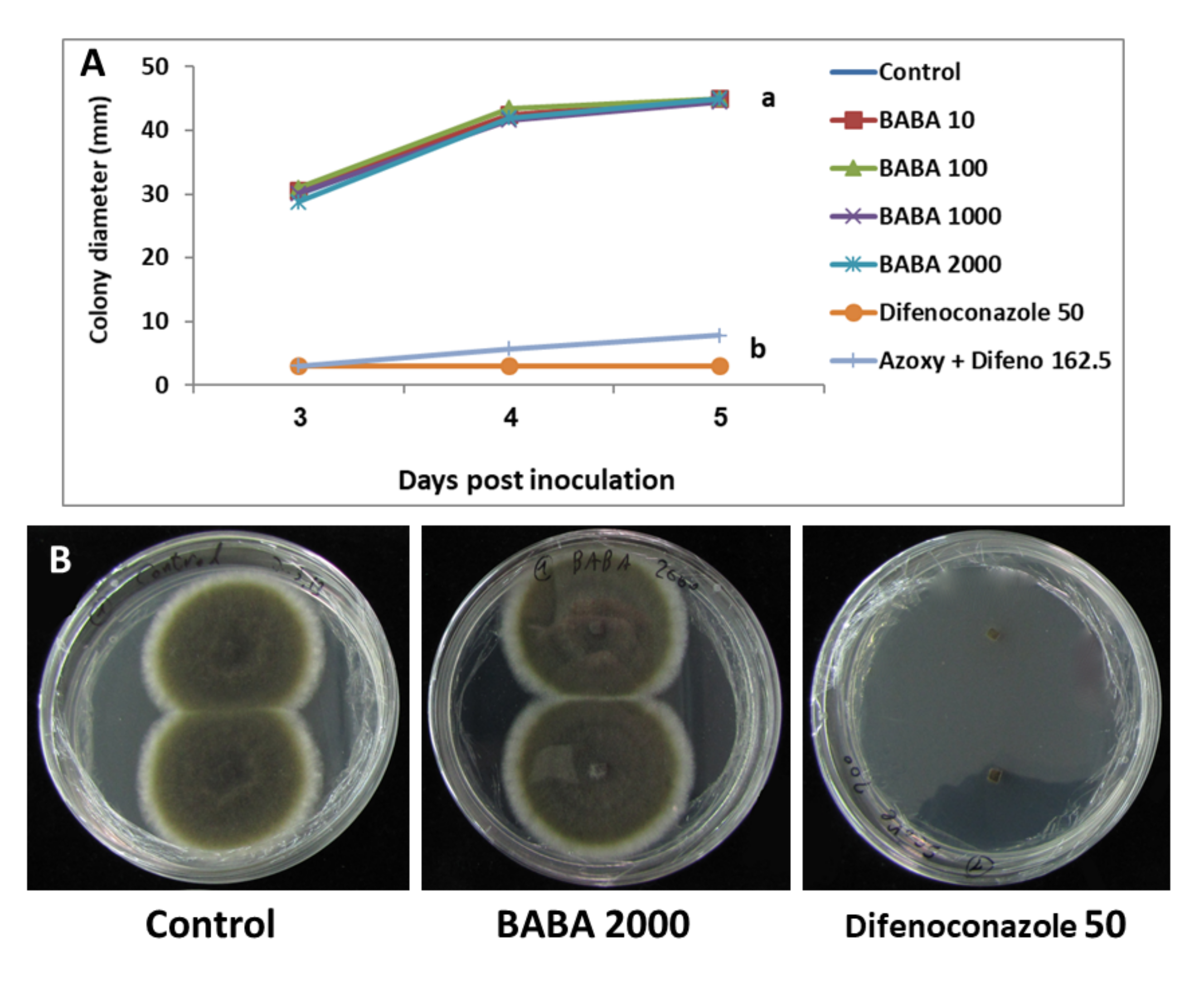
3.1.2. Mycelial Growth
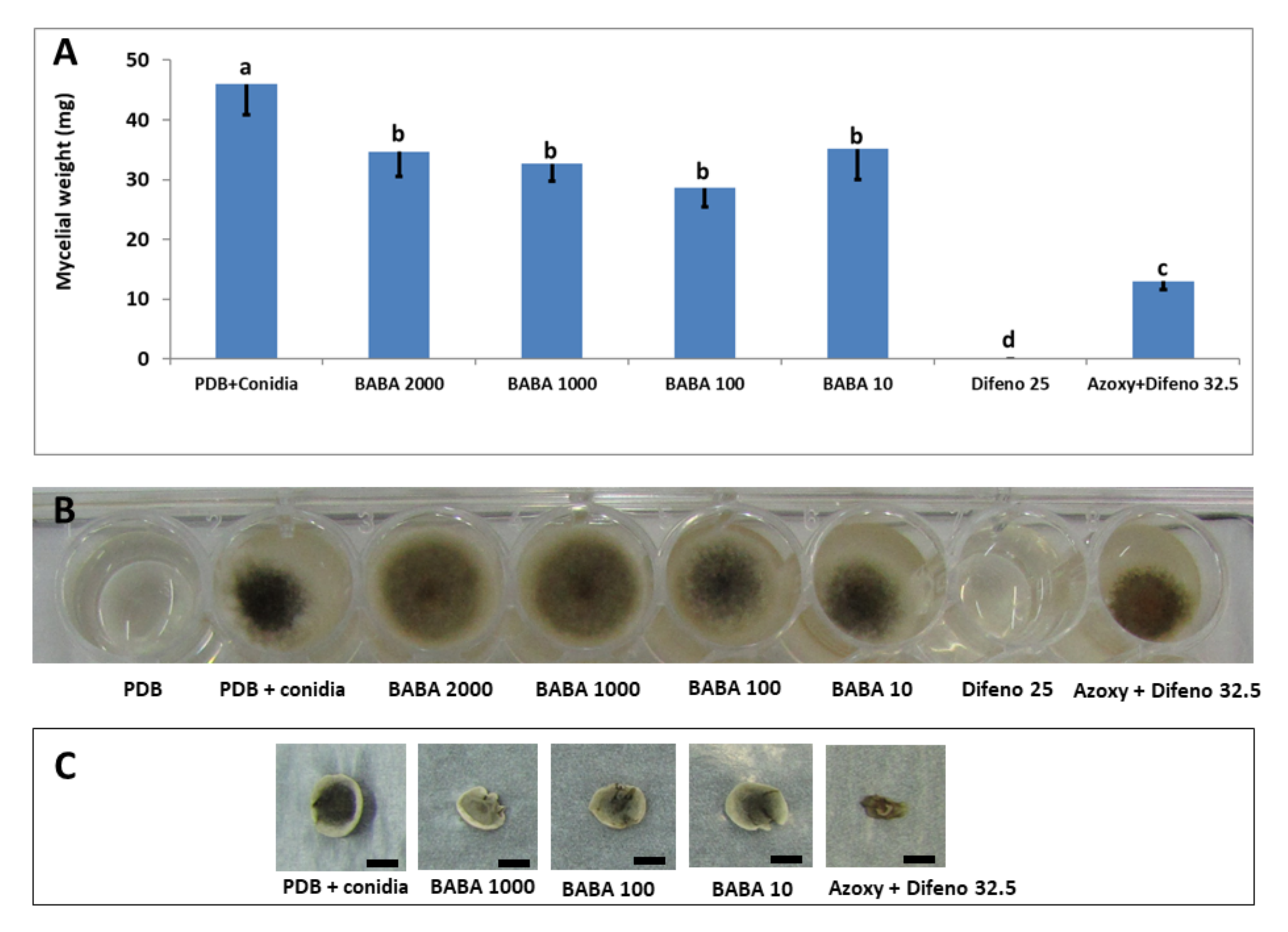
3.1.3. Growth in Liquid Medium
3.2. Experiments on Detached Fruits
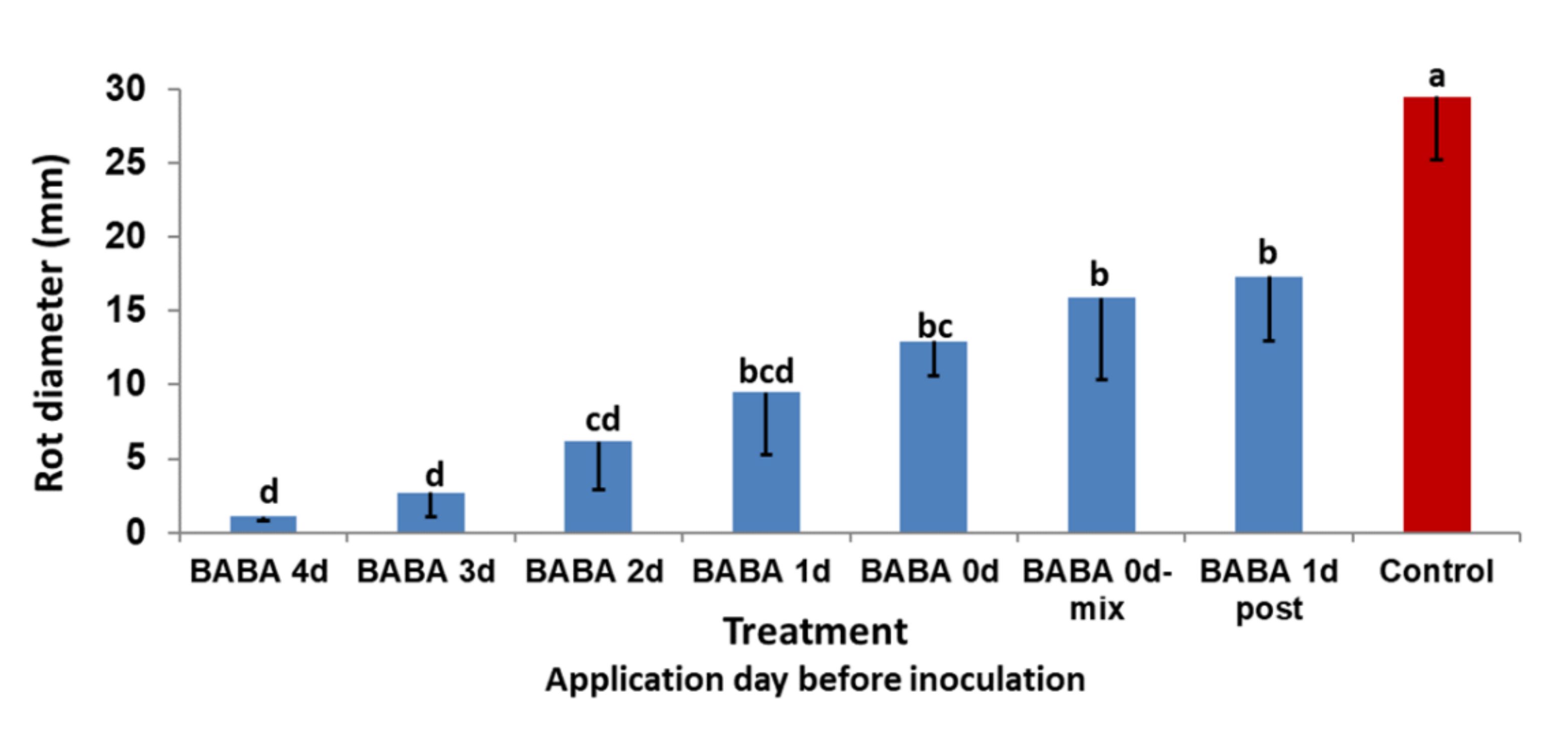
3.2.1. Effect of Application Date on Rot Development
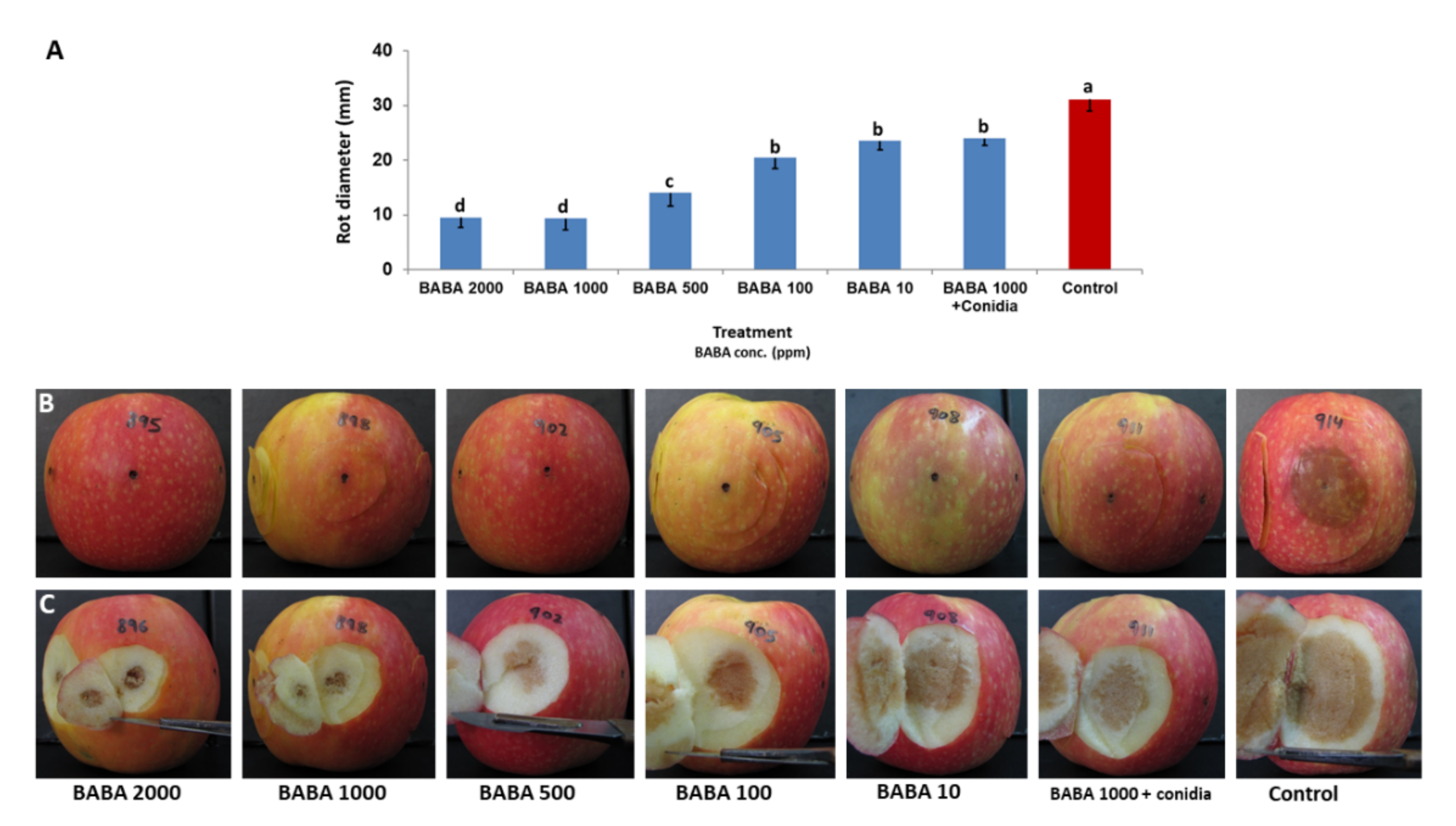
3.2.2. Effect of BABA Concentration on Rot Development
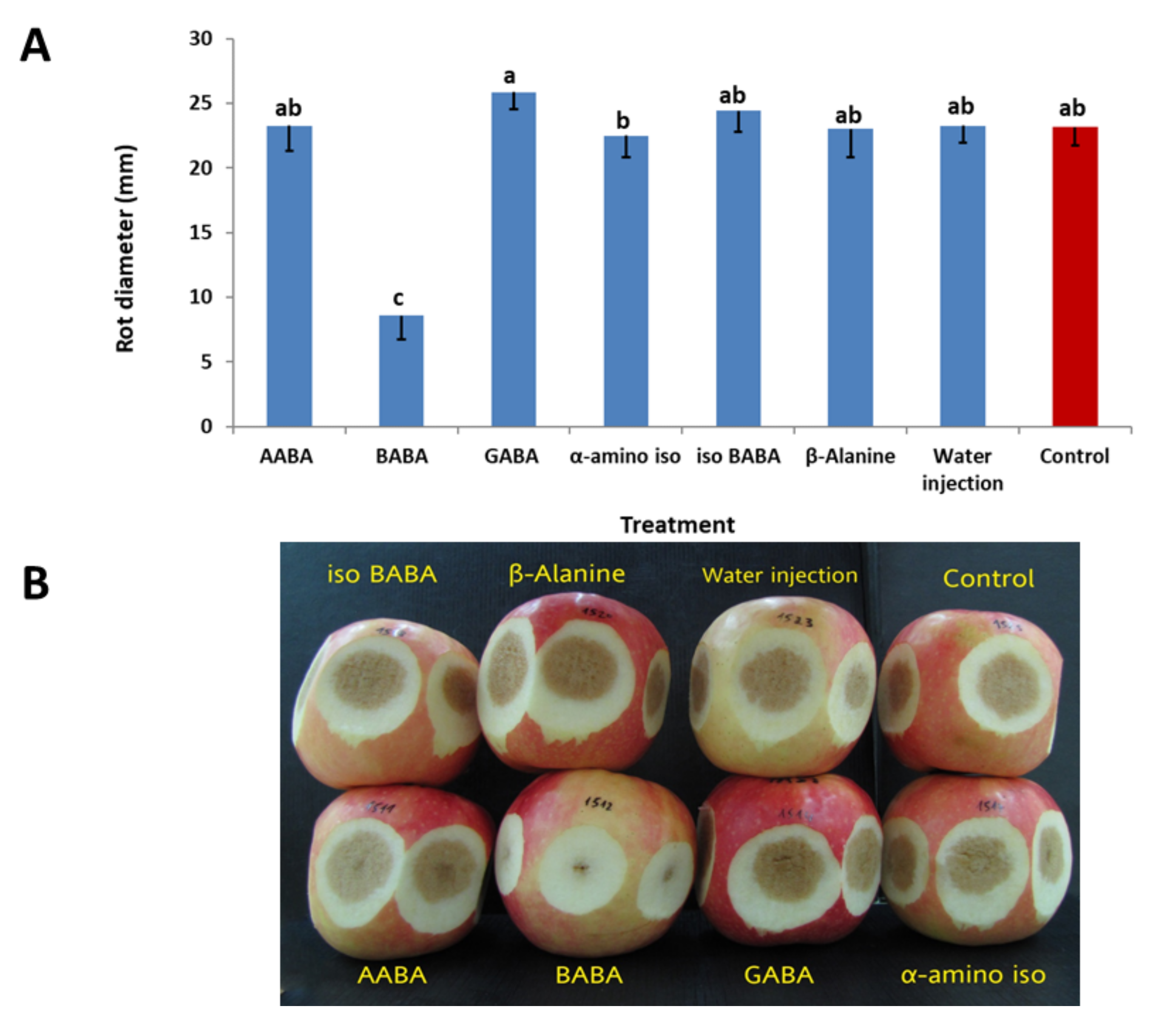
3.2.3. Efficacy of Various Isomers of BABA
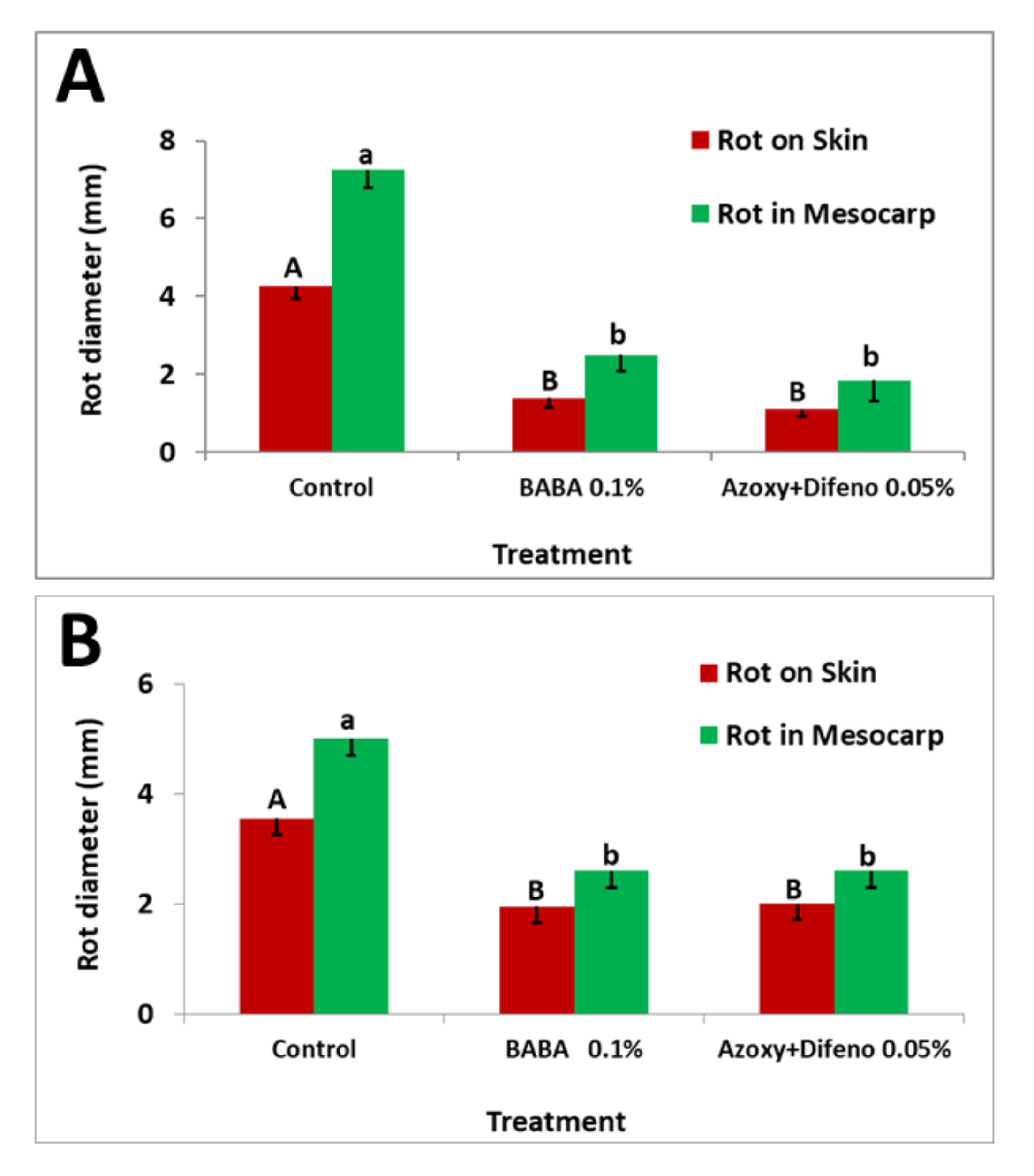
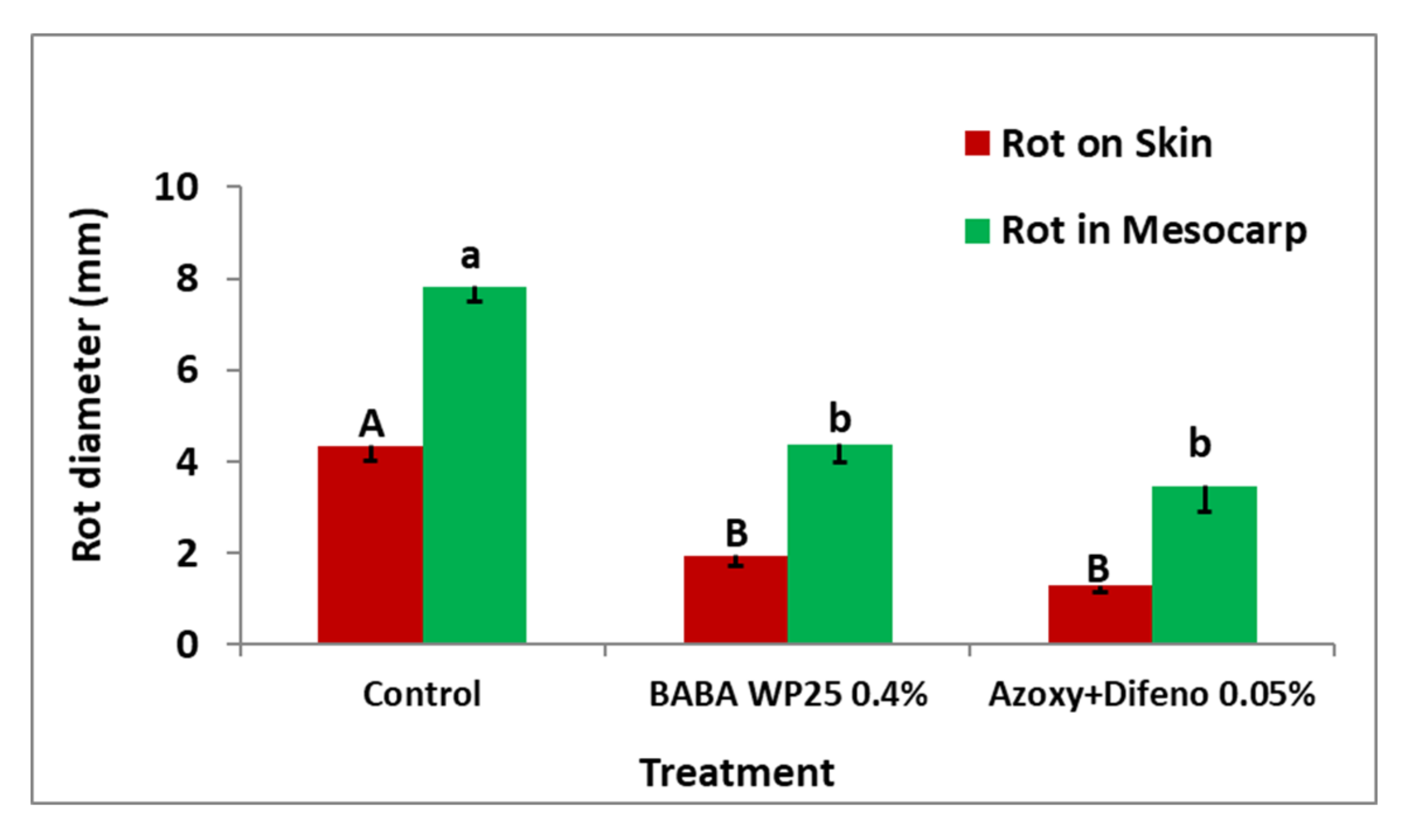
3.3. Field Experiments
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grove, G.G.; Eastwell, K.C.; Jones, A.L.; Sutton, T.B. Diseases of apple. In Apples: Botany, Production and Uses; Ferree, D.C., Warrington, I.J., Eds.; CABI Publishing: Wallingford, UK, 2003; pp. 459–488. [Google Scholar]
- Harteveld, D.O.C.; Akinsanmi, O.A.; Drenth, A. Multiple Alternaria species groups are associated with leaf blotch and fruit spot diseases of apple in Australia. Plant Pathol. 2013, 62, 289–297. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; De Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Bulajic, A.; Filajdic, N.; Babovic, M.; Sutton, T.B. First report of Alternaria mali on apples in Yugoslavia. Plant Dis. 1996, 80, 709. [Google Scholar] [CrossRef]
- Filajdic, N.; Sutton, T.B. Identification and distribution of Alternaria mali on apples in North Carolina and susceptibility of different varieties of apples to Alternaria blotch. Plant Dis. 1991, 75, 1045–1048. [Google Scholar] [CrossRef]
- Hanif, S.; Anjum, T.; Hafeez, R.; Akram, W.; Ali, A.; Hassan, F. First report of Alternaria mali causing core rot of apple in Pakistan. Plant Dis. 2016, 100, 1784. [Google Scholar] [CrossRef]
- Ozgonen, H.; Karaca, G. First report of Alternaria mali causing necrotic leaf spot of apples in Turkey. Plant Pathol. 2006, 55, 578. [Google Scholar] [CrossRef]
- Rotondo, F.; Collina, M.; Brunelli, A.; Pryor, B.M. Comparison of Alternaria spp. collected in Italy from apple with A. mali and other AM-toxin producing strains. Phytopathology 2012, 102, 1130–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawamura, K.; Yukita, K. Alternaria blotch. In Compendium of Apple and Pear Diseases and Pests; Sutton, T.B., Aldwinckle, H.S., Agnello, A.M., Walgenbach, J.F., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2014; pp. 32–33. [Google Scholar]
- Soleimani, M.J.; Esmailzadeh, M. First report of Alternaria mali causing apple leaf blotch disease in Iran. Aust. Plant Dis. Notes 2007, 2, 57–58. [Google Scholar] [CrossRef] [Green Version]
- Gur, L.; Reuveni, M.; Cohen, Y. Occurrence and etiology of Alternaria leaf blotch and fruit spot of apple caused by Alternaria alternata f. sp. mali on cv. Pink lady in Israel. Eur. J. Plant Pathol. 2017, 147, 695–708. [Google Scholar] [CrossRef]
- Ferree, D.C.; Warrington, I.J. Apples: Botany, Production and Uses; CABI Publishing: Wallingford, UK, 2003. [Google Scholar]
- Zhu, L.; Ni, W.; Liu, S.; Cai, B.; Xing, H.; Wang, S. Transcriptomics analysis of apple leaves in response to Alternaria alternata apple pathotype infection. Front. Plant Sci. 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filajdic, N.; Sutton, T.B.; Walgenbach, J.F.; Unrath, C.R. The influence of European red mites on intensity of Alternaria blotch of apple and fruit quality and yield. Plant Dis. 1995, 79, 683–690. [Google Scholar] [CrossRef]
- Filajdic, N.; Sutton, T.B.; Walgenbach, J.F.; Unrath, C.R. The influence of the Apple Aphid/Spirea Aphid complex on intensity of Alternaria blotch of apple and fruit quality characteristics and yield. Plant Dis. 1995, 79, 691–694. [Google Scholar] [CrossRef]
- Jung, K.H. Growth inhibition effect of pyroligneous acid on pathogenic fungus, Alternaria mali, the agent of Alternaria blotch of apple. Biotechnol. Bioprocess. Eng. 2007, 12, 318–322. [Google Scholar] [CrossRef]
- Stern, R.; Ben-Arie, R.; Ginzberg, I. Reducing the incidence of calyx cracking in ‘Pink Lady’apple using a combination of cytokinin (6-benzyladenine) and gibberellins (GA4+7). J. Hortic. Sci. Biotechnol. 2013, 88, 147–153. [Google Scholar]
- Gur, L.; Reuveni, M.; Cohen, Y. Phenology based management of Alternaria fruit rot in Pink Lady apples. Plant Dis. 2018, 102, 1072–1080. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Aldwinckle, H.S.; Sutton, T.; Tsuge, T.; Kang, G.; Cong, P.H.; Cheng, Z.M. Interactions of apple and the Alternaria alternata apple pathotype. Crit. Rev. Plant Sci. 2013, 32, 141–150. [Google Scholar] [CrossRef]
- Gur, L.; Reuveni, M.; Cohen, Y. Control of Alternaria fruit rot in ‘Pink Lady’ apples by fungicidal mixtures. Crop. Prot. 2020, 127, 104947. [Google Scholar] [CrossRef]
- Cohen, Y. β-Aminobutyric acid induces systemic resistance against Peronospore tabacina. Physiol. Mol. Plant Pathol. 1994, 44, 273–288. [Google Scholar] [CrossRef]
- Cohen, Y. Local and systemic control of Phytophthora infestans in tomato plants by DL- β-amino-n-butanoic acids. Phytopathology 1994, 84, 55–59. [Google Scholar] [CrossRef]
- Cohen, Y. Induced resistance against fungal diseases by aminobutyric acids. In Modern Fungicides and Antifungal Compounds; Lyr, H., Russel, P.E., Sisler, H.D., Eds.; Intercept: Andover, UK, 1996; pp. 461–466. [Google Scholar]
- Cohen, Y. β-aminobutyric acid-induced resistance against plant pathogens. Plant Dis. 2002, 86, 448–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, Y.; Vaknin, M.; Mauch-Mani, B. BABA-induced resistance: Milestones along a 55-year journey. Phytoparasitica 2016, 44, 513–538. [Google Scholar] [CrossRef]
- Justyna, P.G.; Ewa, K. Induction of resistance against pathogens by β-aminobutyric acid. Acta Physiol. Plant 2013, 35, 1735–1748. [Google Scholar] [CrossRef]
- Oka, Y.; Cohen, Y.; Spiegel, Y. Local and systemic induced resistance to the root-knot nematode in tomato by DL-β-amino-n-butyric acid. Phytopathology 1999, 89, 1138–1143. [Google Scholar] [CrossRef] [Green Version]
- Reuveni, M.; Sheglov, D.; Cohen, Y. Control of moldy-core decay in apple fruits by ß-aminobutyric acids and potassium phosphites. Plant Dis. 2003, 87, 933–936. [Google Scholar] [CrossRef] [Green Version]
- FRAC (Fungicide Resistance Action Committee). FRAC Code List: Fungicides Sorted by Mode of Action (Including FRAC Code Numbering). Available online: https://www.frac.info/home/news/2020/02/18/the-updated-frac-code-list-2020-is-now-available-for-download (accessed on 1 March 2020).
- Lyon, G.D.; Newton, A.C. Do resistance elicitors offer new opportunities in integrated disease control strategies? Plant Pathol. 1997, 46, 636–641. [Google Scholar] [CrossRef]
- Elsherbiny, E.A.; Dawood, D.H.; Safwat, N.A. Antifungal action and induction of resistance by β-aminobutyric acid against Penicillium digitatum to control green mold in orange fruit. Pestic. Biochem. Physiol. 2021, 171, 104721. [Google Scholar] [CrossRef] [PubMed]
- Ozgonen, H.; Karatas, A. Effect of salicylic acid, DL-beta-amino-n butyric acid and Acibenzolar-s-methyl+ metalaxyl on mycelial growth and spore germination of Alternaria mali in vitro and on young apple seedlings. Int. J. Agric. Biol. 2013, 15, 165–169. [Google Scholar]
- Cohen, Y.; Reuveni, M.; Baider, A. Local and systemic activity of BABA (DL- β- aminobutyric acid) against Plasmopara viticola in grapevines. Eur. J. Plant Pathol. 1999, 105, 351–361. [Google Scholar] [CrossRef]
- Cohen, Y.; Rubin, A.E.; Vaknin, M. Post infection application of DL- β- amino-butyric acid (BABA) induces multiple forms of resistance against Bremia lactucae in lettuce. Eur. J. Plant Pathol. 2011, 130, 13–27. [Google Scholar] [CrossRef]
- Baccelli, I.; Mauch-Mani, B. Beta-aminobutyric acid priming of plant defense: The role of ABA and other hormones. Plant Mol. Biol. 2016, 91, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Chavan, V.; Kamble, A. Induction of total phenolics and defence-related enzymes during β-aminobutyric acid-induced resistance in Brassica carinata against Alternaria blight. Arch. Phytopathol. Plant Protect. 2014, 47, 2200–2212. [Google Scholar] [CrossRef]
- Luna, E.; van Hulten, M.; Zhang, Y.H.; Berkowitz, O.; Lopez, A.; Petriacq, P.; Ton, J. Plant perception of β-aminobutyric acid is mediated by an aspartyl-tRNA synthetase. Nat. Chem. Biol. 2014, 10, 450–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzenbacher, R.E.; Wardell, G.; Stassen, J.; Guest, E.; Zhang, P.; Luna, E.; Ton, J. The IBI1 receptor of β-aminobutyric acid interacts with VOZ transcription factors to regulate abscisic acid signaling and callose-associated defense. Mol. Plant 2020, 13, 1455–1469. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Trivedi, P.; Wang, N. Field evaluation of plant defense inducers for the control of citrus huanglongbing. Phytopathology 2016, 106, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Hwang, B.K.; Yun, J.H. Variability in sensitivity to polyoxin B of isolates of Alternaria mali and decreased fitness of polyoxin resistant isolates. J. Phytopathol. 1986, 115, 305–312. [Google Scholar] [CrossRef]
- Lee, C.U.; Kim, K.H. Cross-tolerance of Alternaria mali to various fungicides. Korean J. Mycol. 1986, 14, 71–78. [Google Scholar]
- Lu, Y.L.; Sutton, T.B.; Ypema, H. Sensitivity of Alternaria mali from North Carolina apple orchards to pyraclostrobin and boscalid. Phytopathology 2003, 93, S54. [Google Scholar]
- Van den Bosch, F.; Paveley, N.; van den Berg, F.; Hobbelen, P.; Oliver, R. Mixtures as a fungicide resistance management tactic. Phytopathology 2014, 104, 1264–1273. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (Conc. µg/mL) | % Germinated Conidia | SE 2 |
|---|---|---|
| Control | 94.7 | 2.80 |
| BABA 10 | 93.7 | 3.33 |
| BABA 100 | 92.5 | 2.75 |
| BABA 1000 | 95.4 | 2.46 |
| BABA 2000 | 88.6 | 3.16 |
| Azoxy + Difeno 162.5 1 | 0 * | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gur, L.; Reuveni, M.; Cohen, Y. β-Aminobutyric Acid Induced Resistance against Alternaria Fruit Rot in Apple Fruits. J. Fungi 2021, 7, 564. https://doi.org/10.3390/jof7070564
Gur L, Reuveni M, Cohen Y. β-Aminobutyric Acid Induced Resistance against Alternaria Fruit Rot in Apple Fruits. Journal of Fungi. 2021; 7(7):564. https://doi.org/10.3390/jof7070564
Chicago/Turabian StyleGur, Lior, Moshe Reuveni, and Yigal Cohen. 2021. "β-Aminobutyric Acid Induced Resistance against Alternaria Fruit Rot in Apple Fruits" Journal of Fungi 7, no. 7: 564. https://doi.org/10.3390/jof7070564