
Freeing Aspergillus fumigatus of Polymycovirus Infection Renders It More Resistant to Competition with Pseudomonas aeruginosa Due to Altered Iron-Acquiring Tactics

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates
2.2. Fungal Strain Cultivation
2.3. Extraction of Siderophores and Calibration
2.4. Liquid Chromatography and Mass Spectrometry
2.5. Statistical Analysis
3. Results
3.1. Pigment Secretion Is Observed by Virus-Infected but Not VF A. fumigatus
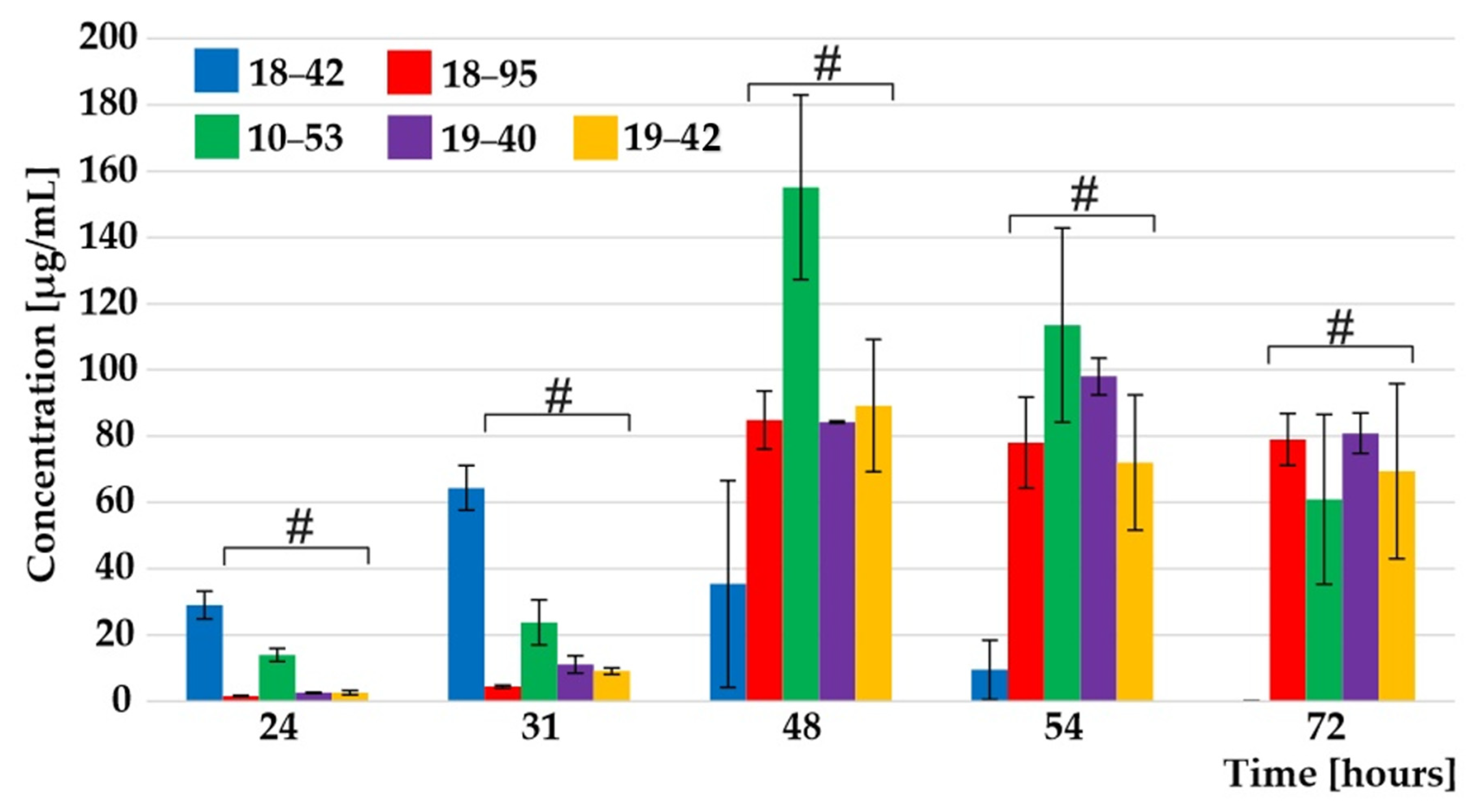
3.2. Monitoring Extracellular Secretion Kinetics Reveals Differential Secretion of Siderophores
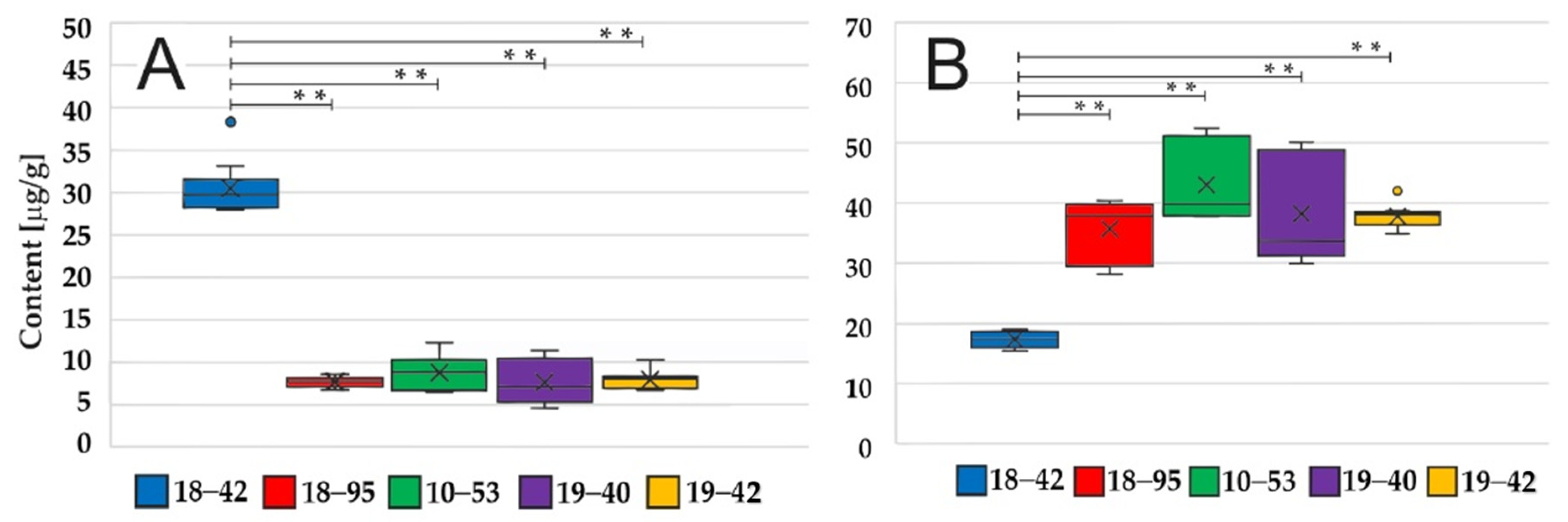
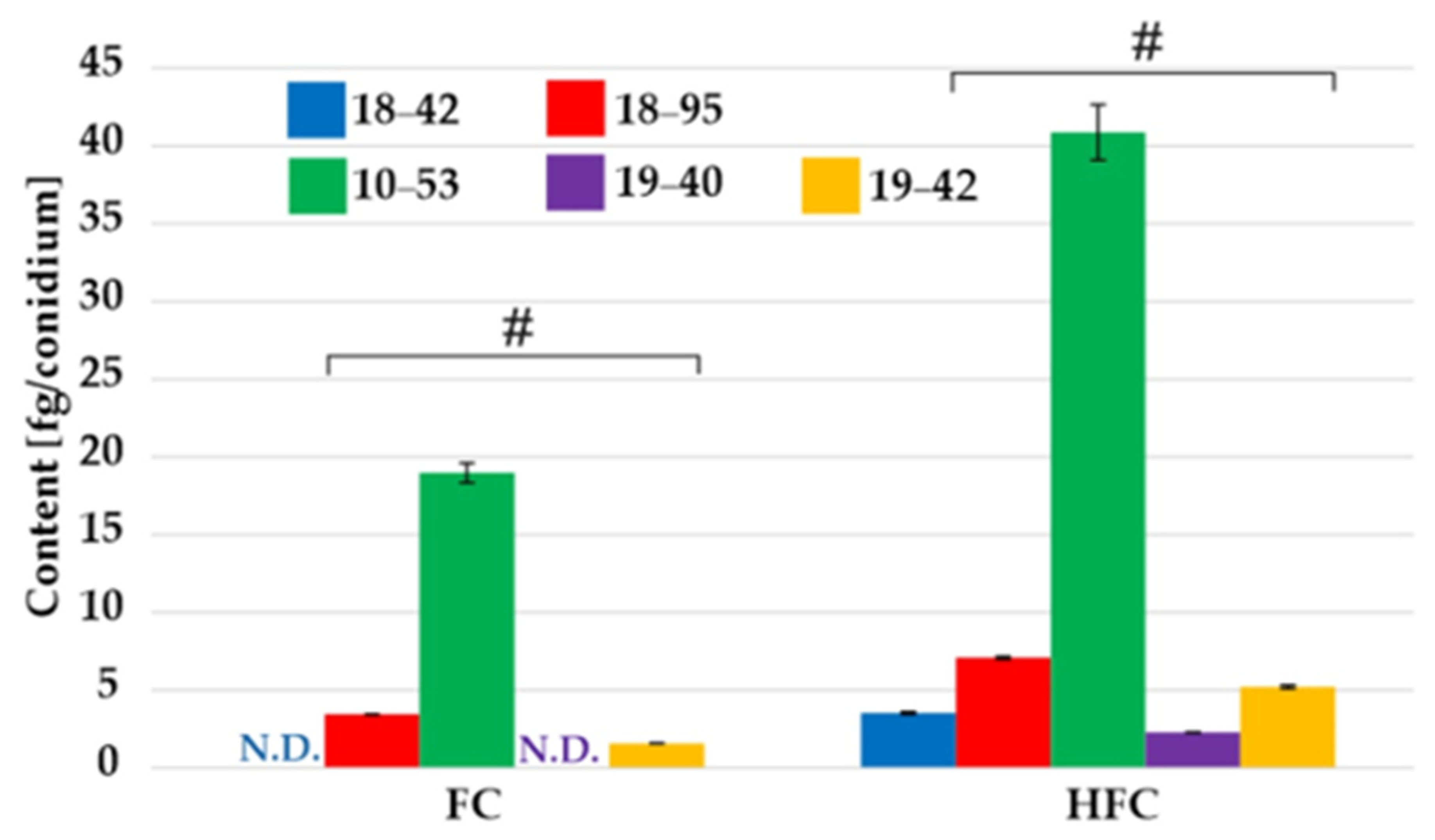
3.3. Fungal Pellets but Not Conidia Have Different HFC/FC Ratios
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Dobiáš, R.; Havlíček, V. Microbial siderophores: Markers of infectious diseases. In Microbial and Natural Macromolecules: Synthesis and Applications; Das, S., Das, H.R., Eds.; Elsevier Academic Press: London, UK, 2021; ISBN 978-0-12-820084-1. [Google Scholar]
- Rutsaert, L.; Steinfort, N.; Van Hunsel, T.; Bomans, P.; Naesens, R.; Mertes, H.; Dits, H.; Van Regenmortel, N. COVID-19-associated invasive pulmonary aspergillosis. Ann. Intensive Care 2020, 10, 71. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Coutts, R.H.A. Mycoviruses in aspergilli: A comprehensive review. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.; Sass, G.; Swietnicki, W.; Stevens, D.A. Review of potential Pseudomonas weaponry, relevant to the Pseudomonas–Aspergillus interplay, for the mycology community. J. Fungi 2020, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Kanhayuwa, L.; Kotta-Loizou, I.; Özkan, S.; Gunning, A.P.; Coutts, R.H.A. A novel mycovirus from Aspergillus fumigatus contains four unique dsRNAs as its genome and is infectious as dsRNA. Proc. Natl. Acad. Sci. USA 2015, 112, 9100–9105. [Google Scholar] [CrossRef]
- Nazik, H.; Kotta-Loizou, I.; Sass, G.; Coutts, R.H.H.; Stevens, D.A. Virus infection of Aspergillus fumigatus compromises the fungus in intermicrobial competition. Viruses 2021, 13, 686. [Google Scholar] [CrossRef]
- Nazik, H.; Sass, G.; Deziel, E.; Stevens, D.A. Aspergillus is inhibited by Pseudomonas aeruginosa volatiles. J. Fungi 2020, 6, 118. [Google Scholar] [CrossRef]
- Wallner, A.; Blatzer, M.; Schrettl, M.; Sarg, B.; Lindner, H.; Haas, H. Ferricrocin, a siderophore involved in intra- and transcellular iron distribution in Aspergillus fumigatus. Appl. Environ. Microb. 2009, 75, 4194–4196. [Google Scholar] [CrossRef] [PubMed]
- Oide, S.; Berthiller, F.; Wiesenberger, G.; Adam, G.; Turgeon, B.G. Individual and combined roles of malonichrome, ferricrocin, and TafC siderophores in Fusarium graminearum pathogenic and sexual development. Front. Microbiol. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Blatzer, M.; Schrettl, M.; Sarg, B.; Lindner, H.H.; Pfaller, K.; Haas, H. SidL, an Aspergillus fumigatus transacetylase involved in biosynthesis of the siderophores ferricrocin and hydroxyferricrocin. Appl. Environ. Microbiol. 2011, 77, 4959–4966. [Google Scholar] [CrossRef]
- Kotta-Loizou, I. Mycoviruses and their role in fungal pathogenesis. Curr. Opin. Microbiol. 2021, in press. [Google Scholar] [CrossRef]
- Filippou, C.; Diss, R.M.; Daudu, J.O.; Coutts, R.H.A.; Kotta-Loizou, I. The polymycovirus-mediated growth enhancement of the entomopathogenic fungus Beauveria bassiana is dependent on carbon and nitrogen metabolism. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Özkan, S.; Coutts, R.H.A. Aspergillus fumigatus mycovirus causes mild hypervirulent effect on pathogenicity when tested on Galleria mellonella. Fungal Genet. Biol. 2015, 76, 20–26. [Google Scholar] [CrossRef]
- Lagashetti, A.C.; Dufossé, L.; Singh, S.K.; Singh, P.N. Fungal pigments and their prospects in different industries. Microorganisms 2019, 7, 604. [Google Scholar] [CrossRef]
- Heinekamp, T.; Thywissen, A.; Macheleidt, J.; Keller, S.; Valiante, V.; Brakhage, A. Aspergillus fumigatus melanins: Interference with the host endocytosis pathway and impact on virulence. Front. Microbiol. 2013, 3. [Google Scholar] [CrossRef]
- Fuller, K.K.; Cramer, R.A.; Zegans, M.E.; Dunlap, J.C.; Loros, J.J. Aspergillus fumigatus photobiology illuminates the marked heterogeneity between isolates. MBio 2016, 7, e01517. [Google Scholar] [CrossRef]
- Takahashi-Nakaguchi, A.; Shishido, E.; Yahara, M.; Urayama, S.-I.; Ninomiya, A.; Chiba, Y.; Sakai, K.; Hagiwara, D.; Chibana, H.; Moriyama, H.; et al. Phenotypic and molecular biological analysis of polymycovirus AfuPmv-1M from Aspergillus fumigatus: Reduced fungal virulence in a mouse infection model. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Liu, G.-L.; Chi, Z.; Wang, G.-Y.; Wang, Z.-P.; Li, Y.; Chi, Z.-M. Yeast killer toxins, molecular mechanisms of their action and their applications. Crit. Rev. Biotechnol. 2015, 35, 222–234. [Google Scholar] [CrossRef]
- Van de Sande, W.W.J.; Vonk, A.G. Mycovirus therapy for invasive pulmonary aspergillosis? Med. Mycol. 2019, 57, S179–S188. [Google Scholar] [CrossRef]
- Schmidt, F.; Lemke, P.; Esser, K. Viral influences on aflatoxin formation by Aspergillus flavus. Appl. Microbiol. Biotechnol. 1986, 24, 248–252. [Google Scholar] [CrossRef]
- Silva, V.N.; Durigon, E.L.; Pires, M.d.F.C.; Lourenço, A.; Faria, M.J.d.; Corrêa, B. Time course of virus-like particles (VLPs) double-stranded RNA accumulation in toxigenic and non-toxigenic strains of Aspergillus flavus. Braz. J. Microbiol. 2001, 32, 56–60. [Google Scholar] [CrossRef]
- Nerva, L.; Chitarra, W.; Siciliano, I.; Gaiotti, F.; Ciuffo, M.; Forgia, M.; Varese, G.C.; Turina, M. Mycoviruses mediate mycotoxin regulation in Aspergillus ochraceus. Environ. Microbiol. 2019, 21, 1957–1968. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Nakaguchi, A.; Shishido, E.; Yahara, M.; Urayama, S.I.; Sakai, K.; Chibana, H.; Kamei, K.; Moriyama, H.; Gonoi, T. Analysis of an intrinsic mycovirus associated with reduced virulence of the human pathogenic fungus Aspergillus fumigatus. Front. Microbiol. 2019, 10, 3045. [Google Scholar] [CrossRef] [PubMed]
- Özkan, S.; Mohorianu, I.; Xu, P.; Dalmay, T.; Coutts, R.H.A. Profile and functional analysis of small RNAs derived from Aspergillus fumigatus infected with double-stranded RNA mycoviruses. BMC Genom. 2017, 18, 416. [Google Scholar] [CrossRef] [PubMed]
- Matthaiou, E.I.; Sass, G.; Stevens, D.A.; Hsu, J.L. Iron: An essential nutrient for Aspergillus fumigatus and a fulcrum for pathogenesis. Curr. Opin. Infect. Dis. 2018, 31, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Houšť, J.; Spížek, J.; Havlíček, V. Antifungal Drugs. Metabolites 2020, 10, 106. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Designation | Conidia $ (×108) | Cdw (mg) # |
|---|---|---|---|
| 18–42 (VF) | UK Af293 cured from AfuPmV-1 | 7.03 | 55.3 ± 3.4 |
| 18–95 | UK Af293 with AfuPmV-1 | 2.75 | 42.6 ± 3.9 |
| 10–53 | USA Af293 with AfuPmV-1 | 2.30 | 42.1 ± 0.5 |
| 19–40 | 18–42 re-infected with AfuPmV-1 | 2.25 | 43.1 ± 3.3 |
| 19–42 | 18–42 re-infected with AfuPmV-1 | 1.84 | 42.5 ± 3.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patil, R.H.; Kotta-Loizou, I.; Palyzová, A.; Pluháček, T.; Coutts, R.H.A.; Stevens, D.A.; Havlíček, V. Freeing Aspergillus fumigatus of Polymycovirus Infection Renders It More Resistant to Competition with Pseudomonas aeruginosa Due to Altered Iron-Acquiring Tactics. J. Fungi 2021, 7, 497. https://doi.org/10.3390/jof7070497
Patil RH, Kotta-Loizou I, Palyzová A, Pluháček T, Coutts RHA, Stevens DA, Havlíček V. Freeing Aspergillus fumigatus of Polymycovirus Infection Renders It More Resistant to Competition with Pseudomonas aeruginosa Due to Altered Iron-Acquiring Tactics. Journal of Fungi. 2021; 7(7):497. https://doi.org/10.3390/jof7070497
Chicago/Turabian StylePatil, Rutuja H., Ioly Kotta-Loizou, Andrea Palyzová, Tomáš Pluháček, Robert H. A. Coutts, David A. Stevens, and Vladimír Havlíček. 2021. "Freeing Aspergillus fumigatus of Polymycovirus Infection Renders It More Resistant to Competition with Pseudomonas aeruginosa Due to Altered Iron-Acquiring Tactics" Journal of Fungi 7, no. 7: 497. https://doi.org/10.3390/jof7070497