3.2. Taxonomy
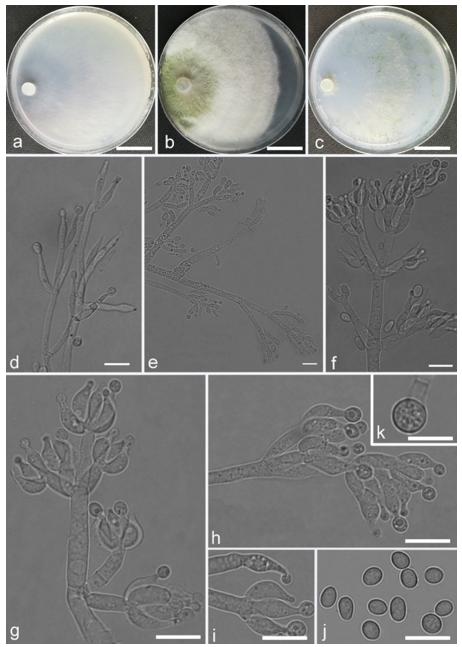
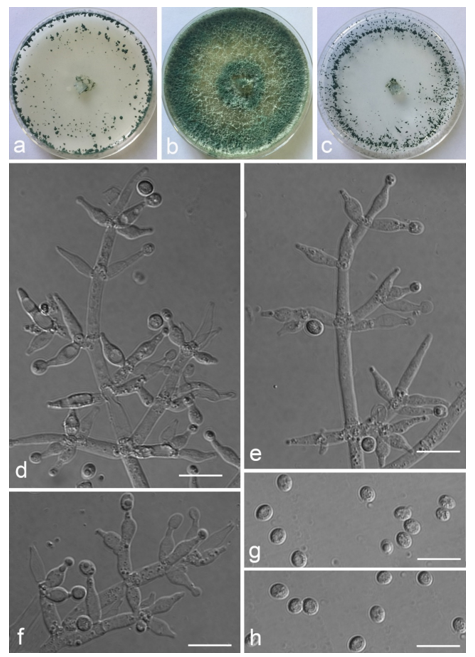
Trichoderma achlamydosporum Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 2)
MycoBank MB 834558
Etymology. Latin, a-, meaning without, not, -chlamydosporum, referred to the chlamydospore.
Type: CHINA, Yunnan province, Jianshui county, N23°15′16″, E102°57′11″, 1170 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06226, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021065.
Conidiophores straight or slightly curved, comprising an indistinguishable main axis and relatively few distantly placed side branches of varying length, branches sparsely disposed, mostly asymmetrically arranged, also paired, or sometimes at irregular intervals along the central axis, not only once again branched, mostly 1–2 steps, often orientated towards the conidiophore terminus, short. Phialides solitary, narrowly lageniform to lanceolate, often asymmetric, straight, less commonly curved or sinuous, generally with long necks, paired or in whorls of 2–4, (6.1–)7.1–13.9(–14.9) × (2.0–)2.4–3.2(–3.5) μm, l/w ratio (2.0–)2.5–6.7 (–7.3), 1.6–2.4(–2.7) μm-wide at the base. Conidia oval to ellipsoid, less commonly subglobose or oblong, green, smooth, 3.2–4.2 × 2.3–3.0 μm, l/w ratio 1.2–1.5.
Culture characteristics: Optimum temperature for growth 25 °C. No growth at 35 °C in PDA, CMD, and SNA. Colony radius on CMD after 72 h: 38 mm at 25 °C, 27 mm at 30 °C, covering the plate after 6 days at 25 °C, white zonation conspicuous. Colonies hyaline, circular, mycelium dense, formed reticular texture. Aerial hyphae scant, only found along the colony margin. Chlamydospores were unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 43 mm at 25 °C, 21 mm at 30 °C, covering the plate after 6 days at 25 °C, zonation conspicuous. Colonies translucent, mycelium dense, forming a membraniform covering layer on the medium. Aerial hyphae formed, white, sterile belt that 8 mm from colony margin. Chlamydospores unobserved. Light yellow pigments noted after 4 days. No distinct odor noted. Colony radius on SNA after 72 h: 33 mm at 25 °C, 27 mm at 30 °C, covering the plate after 6 days at 25 °C. Colonies characteristics resemble to those on CMD, but with denser mycelium.
Notes: Phylogenetically,
Trichoderma achlamydosporum is close to
T. longiphialidicum Q.V. Montoya et al., and
T. polypori K. Chen & W.Y. Zhuang. However, the growth rate of
T. achlamydosporum is slower than
T. longiphialidicum at 25 °C on PDA, CMD, and SNA. Moreover,
T. achlamydosporum has shorter phialides [(6.1–)7.1–13.9(–14.9) μm vs. 7.0–21 μm] and larger conidia (3.2–4.2 × 2.3–3.0 μm vs. 2.5–3.5 × 1.6–2.4 μm) than
T. longiphialidicum [
54].
T. polypori is different from
T. achlamydosporum by having more complex branches, longer phialides [(8.9–)11.7–16.6(−20.0) μm], and wider conidia (2.5–3.3 μm) [
44]. In addition,
T. longiphialidicum and
T. polypori can grow at 35 °C and were observed the production of chlamydospores.
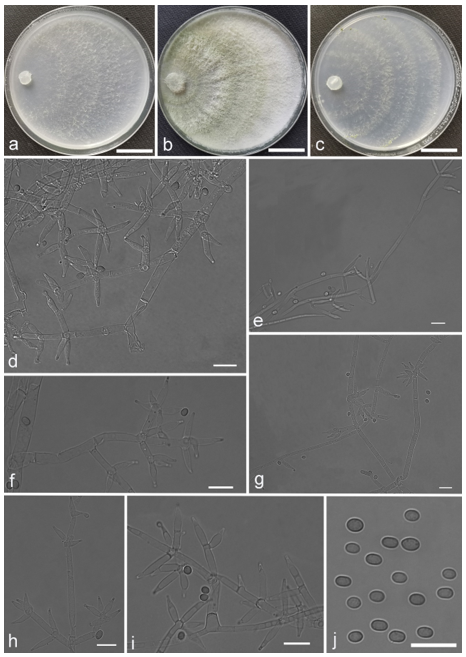
Trichoderma amoenum Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 3)
MycoBank MB 834569
Etymology. Latin, amoenum, meaning beautiful, pleasing.
Type: CHINA, Yunnan province, Jianshui county, N23°15′17″, E102°57′15″, 1170 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06209, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021068.
Conidiophores verticillium-like, straight or slightly curved, paired or unpaired, rebranching on 1–2 levels, sometimes substituted by phialides singly or in whorls of 3–5(–7). Phialides ampuliform to tube, few circular with homogeneous necks, widest around the middle, (3.6–)4.3–10.0(–14.9) × (2.1–)2.5–3.8(–4.5) μm, l/w ratio (1.0–)1.4–4.6(–5.8), 1.4–2.9(–3.7) μm wide at the base. Conidia smooth, green, oval, subspheroidal, (3.0–)3.7–4.5(–5.0) × 3.2–3.9 μm, l/w ratio 1.0–1.4. Chlamydospores globose, smooth, terminal, 6.7–8.9 × 5.5–8.7 μm.
Culture characteristics: Optimum temperature for growth 25 °C. Colony radius on CMD after 72 h: 55 mm at 25 °C, 47 mm at 30 °C, covering the plate after 5 days at 30 °C, attaining 2 mm after 7 days at 35 °C. Colonies hyaline, mycelia forming subsurface around the inoculum but aerial hyphae present at the margin. Aerial hyphae scanty, and coiling slightly. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: cover the plate at 25 °C, 55 mm at 30 °C, attaining 13 mm after 7 days at 35 °C. Colonies white, radial, loose. Aerial hyphae dense, clustered on the outer layer of the colony and extending to the Petri dish lid that gives the colony a downy texture. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 42 mm at 25 °C, 22 mm at 30 °C, covering the plate after 5 days at 25 °C. Colonies green, circular, radial, zonate. Aerial hyphae scant, inconspicuous. Conidiation formed in pustules after 2 days, pustules abundant in kelly and bottle-green bands, spreading in 4–5 concentric rings. No diffusing pigment noted, odor indistinct.
Additional specimen examined: CHINA, Yunnan province, Jianshui county, N23°15′17″, E102°57′15″, 1170 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv., living culture YMF 1.06210.
Notes: Phylogenetically,
Trichoderma amoenum is closely related to
T. koningii Oudemans & Koning and
T. koningiopsis. However,
T. amoenum can be distinguished from them in morphology.
T. amoenum forms intricately branched conidiophores with branches that bear numerous ampuliform phialides in whorls of 3–5(–7) and has smooth, oval to subspheroidal conidia, while
T. koningii has lageniform phialides in whorls of 3–4 and oblong conidia [
55].
T. koningiopsis resembles
T. amoenum by having similar conidiophores, but has narrower conidia (2.2–3.0 μm vs. 3.2–3.9 μm) [
55].
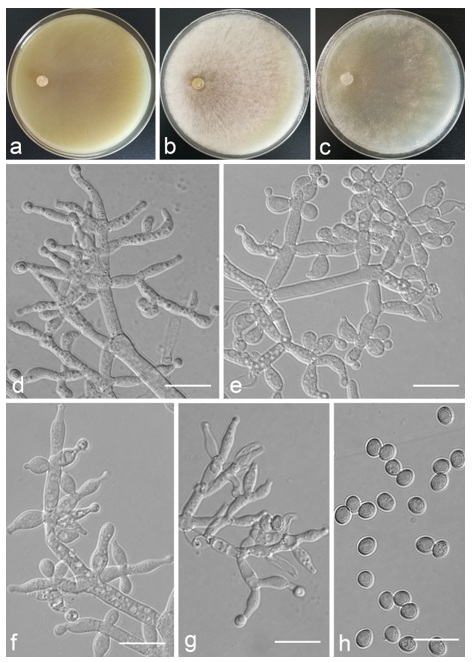
Trichoderma anaharzianum Z.F. Yu & X. Du, sp. nov. (
Figure 4)
MycoBank MB825471
Etymology. Greek, ana-, meaning up to, toward, exceedingly, back, against, -harzianum, referring to the species Trichoderma harzianum.
Type: CHINA, Yunnan province, Luliang county, N24°57′22″, E103°46′30″, 1800 m alt., from soil of tobacco rhizosphere, July 2007, Z.F. Yu, Y.F. Lv. Holotype YMF 1.00383, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19086.
Conidiophores straight or slightly curved, numerous, branched, commonly contain paired branches, re-branching 1–2 times, 3.0–5.0 μm wide at base, the distance between two neighboring branches is (15.0–)19.0–23.0(–27.0) μm. The branches generally perpendicular to main axis, terminating in 3–7 phialides. Phialides ampulliform to lageniform in a verticillate fashion, sometimes arising singly directly from the main axis, (4.0–)5.0–6.0(–7.0) × 2.0–4.0 μm, l/w ratio (1.0–)1.3–2.0(–2.3). Conidia globose to subglobose, thin-walled, green, smooth, (2.5–)2.6–3.1(–3.2) × (2.1–)2.2–2.8(–3.0) μm, l/w ratio (1.0–)1.1–1.3(–1.4).
Culture characteristics: Optimum temperature for growth 25 °C. Colony radius on CMD after 72 h: cover the plate at 25 °C, 38 mm at 30 °C, attaining 33 mm after 7 days at 35 °C. Colonies homogenous, translucent, radial. Aerial hyphae distinct, loosely aggregated. Conidiation formed on minute pustules after 3 days, pustule surface with roundish or irregular outline, successively turning green after 5 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 64–76 mm at 25 °C, 50 mm at 30 °C, 11–18 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies circular and dense. Aerial hyphae abundant, whitish arachnoid, Conidia noted after 3 days, and successively turning green after 4 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 46–53 mm at 25 °C, 30–35 mm at 30 °C, 6 mm at 35 °C, covering the plate after 6 days at 25 °C. Colonies hyaline, loose, radial, indistinctly zonate. Aerial hyphae white and sparse, mostly around the center of colony, conidia noted after 4 days, gradually turning green after 5 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Additional specimen examined: CHINA, Yunnan province, Luliang county, N24°57′22″, E103°46′30″, 1800 m alt., from soil of tobacco rhizosphere, July 2007, Z.F. Yu, living culture YMF 1.00241.
Notes:
Trichoderma anaharzianum is phylogenetically closely related to
T. harzianum Rifai. Morphologically,
T. anaharzianum has a wider branch base. Moreover, the conidia of
T. harzianum forms from thick and dense concentric ring on PDA and SNA, or uniformly throughout the colony [
56], while this was not observed on
T. anaharzianum.
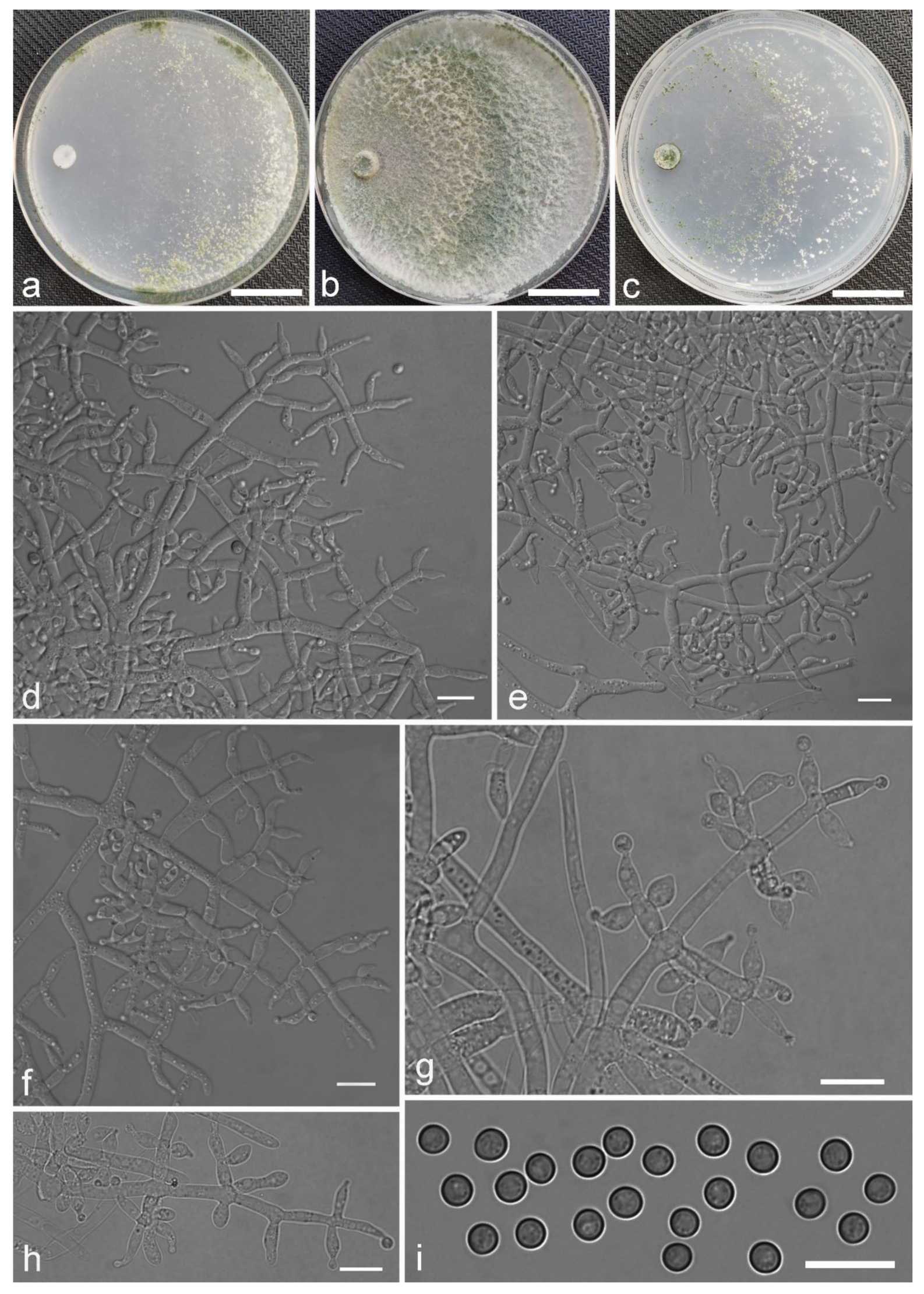
Trichoderma anisohamatum Z. F. Yu & X. Du, sp. nov. (
Figure 5)
MycoBank MB825467
Etymology. Greek, aniso-, meaning unequal, uneven, -hamatum, referring to the species Trichoderma hamatum.
Type: CHINA, Yunnan province, Luliang county, N24°57′22″, E103°46′30″, 1800 m alt., from soil of tobacco rhizosphere, July 2007, Z.F. Yu. Holotype YMF 1.00333, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19083.
Conidiophores regularly dendriform, straight or slightly curved, comprising a distinct main axis with side branches paired or unilateral and often terminating in whorl of 3–5 divergent phialides, rarely in whorl of 7, 3.0–4.0(–5.0) μm wide at the base, the distance between two neighbor branches is (19.0–)20.0–24.0(–26.0) μm. Phialides mostly lageniforms, spindly, sometimes with cylindrical bent neck, fasciculate near the tip of main axis, occasionally solitary, 5.0–8.0 × 3.0–4.0(–5.0) μm, l/w ratio (1.0–)1.3–2.3(–2.7). Conidia subglobose to globose, sometimes ellipsoidal, green, smooth, (3.6–)3.8–4.6(–4.8) × (2.7–)2.8–3.3(–3.4) μm, l/w ratio (1.1–)1.2–1.6(–1.7).
Culture characteristics: Optimum temperature for growth 25 °C. No growth at 35 °C in PDA, CMD and SNA. Colony radius on CMD after 72 h: 51–59 mm at 25 °C, 16 mm at 30 °C, covering the plate after 4 days at 25 °C. Colonies hyaline, circular, margin distinct. Aerial hyphae inconspicuous. Conidiation formed in pustules after 3 days, pustules relatively sparse, spreading on the side of inoculum. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 64–68 mm at 25 °C, 16 mm at 30 °C, covering the plate after 4 days at 25 °C. Colonies layered distinctly, green in the center, white at the margin, radial. Aerial hyphae hairy to floccose, dense on inner layer, but relative sparse on the margin, a large green disk around the inoculums. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 45–49 mm at 25 °C, 10 mm at 30 °C, covering the plate after 5 days at 25 °C. Colonies green in center, white at the margin, margin well-fined. Aerial hyphae sparse on outer layer, radiated indistinctly. Conidiation formed in pustules after 3 days, pustules abundant, originally white and gradually turning green. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Additional specimens examined: China, Yunnan province, Yunnan province, Luliang county, N24°57′22″, E103°46′30″, 1800 m alt., from soil of tobacco rhizosphere, July 2007, Z.F. Yu, living culture YMF 1.00253, YMF 1.00333.
Notes:
Trichoderma anisohamatum differs morphologically from the phylogenetically nearest species
T. hamatum by its sterile secondary branches. Furthermore, the phialides of
T. hamatum are somewhat swollen in the middle [
57].
Trichoderma aquatica Z.F. Yu & X. Du, sp. nov. (
Figure 6)
MycoBank MB830633
Etymology. Latin, aquatica, referring to the isolated source from water environment.
Type: CHINA, Sichuan province, Luhuo county, N31°39′7.5″, E100°15′31.9″, 3507 m alt., endophytic in living root of Batrachium bungei, July 2014, Z.F. Yu, Y. Huang. Holotype YMF 1.04625, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19077.
Conidiophores macronematous, mononematous, pyramidal aspect, alternately or irregular branched, often terminating in a verticil of the 3–5 phialides, the distance between two neighboring branches is 5.5–19.1 μm. Phialides lageniform, rare ampulliform, slightly curved, 5.2–11.5 × (2.5–)3.1–4.2 μm, l/w ratio 1.1–2.8(–3.0), discrete, perpendicular to the main axis or integrated terminal arranged, sometimes arising in crowded and compact fascicles or singly directly from the main axis, 2.4–4.4 μm wide at base. Conidia mostly ovoid, ellipsoidal, rarely subglobose, thin-walled, green, smooth, 3.3–4.8 × 2.4–3.3 μm, l/w ratio 1.1–1.6. Chlamydospores globose, smooth, terminal, 4.3–7.6 × 3.91–7.2 μm.
Culture characteristics: Colony grows very fast on CMD and PDA. Colony radius on CMD after 72 h: cover the plate at 25 °C, 30 °C, and 35 °C. Colonies hyaline, thin, mycelium sparse. Aerial hyphae sparse, scant, inconspicuous. Conidiation minute, formed in pustules after 3 days, pustules relatively sparse, spreading uniformly throughout the colony, turning dark green after 4 days. Light yellow pigments noted after 3 days. No distinct odor noted. Colony radius on PDA after 72 h: cover the plate at 25 °C, 30 °C, and 35 °C. Colonies white in the center, dark green in margin, circular, zonate distinctly. Arial hyphae radial distinctly, abundant, dense, gradually forms a continuous lawn, granulated to pulvinate pustules on the margin and central around the inoculums. Conidiation formed in pustules after 3 days, pustules abundant, gradually turning dark green after 4 days. Yellow pigments noted distinctly. No distinct odor noted. Colony radius on SNA after 72 h: 49–55 mm at 25 °C, 47–60 mm at 30 °C, 34–40 mm at 35 °C, covering the plate after 5 days at 25 °C. Colonies not zonate, hyaline. Aerial hyphae sparse, scant, inconspicuous and radial indistinctly. Conidiation formed in pustules after 2 days, pustules minute, compact, scattered, turning dark green rapidly. No diffusing pigment noted, odor indistinct.
Additional specimen examined: CHINA, Sichuan province, Luhuo county, N31°39′7.5″, E100°15′31.9″, 3507 m alt., endophytic in living stem of Batrachium bungei, July 2014, Z.F. Yu, Y. Huang, living culture YMF 1.04624.
Notes: The phylogenetic analyses based on
rpb2 and
tef1 reveal that
Trichoderma aquatica is related to
T. parareesei Jaklitsch et al. and
T. reesei E.G. Simmons.
T. aquatica is morphologically most similar to
T. parareesei in the loosely arranged and unpaired branches, cylindrical-neck phialides and smooth, green conidia, whereas
T. parareesei is obviously distinguished by lageniform, shorter and narrower phialide [(4.5–)5.0–8.0(–11.0) × (2.5–)2.7–3.5(–3.8) μm], and uniformly ellipsoidal, longer and wider conidia [(3.3–)3.8–4.5(–6.2) × (2.5–)2.8–3.2(–3.5) μm] [
58]. Furthermore,
T. reesei has narrower phialides (2.2–)2.5–3.5(–4.0) μm and longer conidia 3.5–6.0 (–9.0) μm [
58], which distinctly differs from mostly ovoid conidia of the new species.
Trichoderma asiaticum Z.F. Yu & X. Du, sp. nov. (
Figure 7)
MycoBank MB825470
Etymology. Latin, asiaticum, referring to the continent of Asia.
Type: CHINA, Yunnan province, Luliang county, N24°57′22″, E103°46′30″, 1800 m alt., from soil of tobacco rhizosphere, July 2007, Z.F. Yu, Y.F. Lv. Holotype YMF 1.00352, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19085.
Conidiophores typically comprise a distinct main axis with one terminal whorl of 4–5 phialides and mostly paired side branches. Branches mostly are perpendicular to the main axis, with septa inconspicuous. The distance between two neighboring branches is (5.0–)6.0–22.0(–24.0) μm. Base not well-defined, about 2.0–4.0 μm wide. Each branch terminating in a whorl of up to 3–5 phialides. Phialides ampulliform to lageniform, (3.0–)4.0–6.0(–7.0) × (1.0–)2.0–3.0(–4.0) μm, l/w ratio (1.0–)1.3–3.0(–4.0), usually verticillated around branch. Conidia commonly subglobose to globose, oblong rarely noted, green, smooth, (2.3–)2.4–3.0(–3.1) × (2.0–)2.1–2.7(–2.8) μm, l/w ratio (1.0–)1.1–1.3(–1.4).
Culture characteristics: Optimum temperature for growth 25 °C. Colony radius on CMD after 72 h: 55 mm at 25 °C, 30–35 mm at 30 °C, 9–13 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies hyaline, outline distinct, fan-shaped. Aerial hyphae loose, sparse, radial and arachnoid, branched distinctly. Pustules white, minute, distributed in a scattered fashion around the point of inoculation, turning green after 5 days. Chlamydospores unobserved. Light yellow pigments noted. No distinct odor noted. Colony radius on PDA after 72 h: 54 mm at 30 °C, 12 mm at 35 °C, covering the plate after 3 days at 25 °C. Colonies thick and dense, the zone around the central part of colony forms a distinct circular and green part. Aerial hyphae whitish, partly green, abundant, floccose to cottony, zonate distinctly. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 58–65 mm at 25 °C, 37 mm at 30 °C, 7 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies similar to that on CMD, regular and distinct outline, but translucent and round-like. Aerial hyphae branched, loose and sparse, radial and arachnoid. Pustules white, minute, scattered around the point of inoculation, gradually turning green after 3 days. Chlamydospores unobserved. Light yellow pigments noted. No distinct odor noted.
Additional specimen examined: CHINA, Yunnan province, Luliang county, N24°57′22″, E103°46′30″, 1800 m alt., from soil of tobacco rhizosphere, July 2007, Z.F. Yu, living culture YMF 1.00168.
Notes:
Trichoderma asiaticum is a member of the
Harzianum clade. Phylogenetically,
T. asiaticum is closely related to
T. afroharzianum Chaverri & Jaklitsch and
T. atrobrunneum Chaverri & Jaklitsch, but the branch base of the two species are narrower [(1.0–)1.5–2.2(–3.5) and 1.0–2.7 mm, respectively][
56]. In addition, some phialides of
T. asiaticum are somewhat shorter than those of
T. afroharzianum [(3.0–)4.0–6.0(–7.0) vs. (3.5–)5.2–10.2(–17.5) mm]. For the
T. atrobrunneum, the base of the branch is narrower than that of
T. asiaticum and the conidia of the
T. atrobrunneum are subglobose to ovoid [
56], which significantly differs from the globose and smaller conidia of
T. asiaticum [(2.3–)2.4–3.0(–3.1) × (2.0–)2.1–2.7(–2.8) μm].
Trichoderma asymmetricum Z.F. Yu & X. Du, sp. nov. (
Figure 8)
MycoBank MB830637
Etymology. Latin, asymmetricus, referring to the asymmetrically arranged branches.
Type: CHINA, Sichuan province, Daocheng county, N29°29′48.5″, E100°14′46.5″, 4362 m alt., endophytic in living leaf of Hippuris vulgaris in pond, July 2014, Z.F. Yu, Y. Huang. Holotype YMF 1.04618, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19164.
Conidiophores typically tree-like, rarely comprising a main axis with a terminal solitary phialide, often crowded, typically asymmetrically branched. The bases of branches are not well-defined, about 2.3–4.2 μm wide. Branches commonly sinuous and swollen below the phialides, diverging to form 1–3 phialides at the terminal. Phialides usually sterile, occasionally lageniform, with the apex generally obtuse, sometimes arising alone from the main axis often with a long base, (4.4–)5.9–11.8(–12.6) × 2.3–4.0(–4.4) μm, l/w ratio (1.3–)1.7–4.3(–5.0). Conidia green, smooth, subglobose to globose, sometimes broadly ellipsoidal, rarely ovoid, 3.5–4.5(–5.0) × 3.2–4.1 μm, l/w ratio 1.0–1.4.
Culture characteristics: Optimum temperature for growth 25 °C. No growth at 30 °C in PDA and CMD, and at 35 °C in SNA. Colony radius on CMD after 72 h: 41 mm at 25 °C. Colonies translucent, thin, indistinctly radial around the point of inoculation, conspicuously arachnoid and relatively dense near the margin. Aerial hyphae common, denser in distant areas. Conidiation formed in pustules after 15 days; pustules white, turning green after 23 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 57 mm at 25 °C, covering the plate after 5 days at 25 °C. Colonies white, circular in outline, compact, indistinctly zonate, with well-defined margin. Aerial hyphae radial conspicuously, dense and extending toward the distal margin. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 37–40 mm at 25 °C, less than 10 mm at 30 °C, covering the plate after 8 days at 25 °C. Colonies hyaline, thin, circular, margin obscure. Aerial hyphae sparse, arachnoid, somewhat branched, becoming fertile latter. Pustules arranged regularly, generally in a broad marginal zone, asymmetrical to hemispherical, formed after 13 days and turned green after 14 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Notes: Phylogenetic analyses showed that
Trichoderma asymmetricum is closely related to
T. viridescens Jaklitsch & Samuels, they both share undiscernible conidiophores and some branches that only have a few phialides. The middle part of phialides of the two species are usually swollen and the necks are long cylindrical. Moreover, the conidia of two species in SNA are mostly subglobose, some ovoid. The biggest difference between two species are that the top of phialides of
T. asymmetricum is often curved rather than constricted, and mostly sterile, which differs from the narrow phialides top of
T. viridescens. The phialide of
T. viridescens generally looks submoniliform. Furthermore, the conidia of
T. asymmetricum are commonly slightly wider than those of
T. viridescens [(2.2–)3.0–3.7(–4.7) μm] [
59].
T. asymmetricum is also phylogenetically close to
T. viridialbum Jaklitsch et al., and the two species have optimal growth at 25 °C on all media, with restricted growth at 30 °C and no growth at 35 °C. However,
T. viridialbum has longer phialides (6.8–)8.8–12.7(–15.0) × (2.2–)2.5–3.0(–3.5) μm than
T. asymmetricum [
60].
Trichoderma inaequilaterale Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 9)
MycoBank MB 834562
Etymology. Latin, inaequilaterale, referring to the asymmetric phialides.
Type: CHINA, Yunnan province, Jianshui county, N23°16′22″, E102°57′37″, 1170 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06203, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021066.
Conidiophores penicillium-like, mostly asymmetrically arranged, below terminal branches also paired or in whorls of 2–3, not only primary branches, mostly 1–3 times at irregular intervals along the central axis, straight or slightly bent, orientating towards the conidiophore terminus. Phialides ampuliform or narrowly lageniform, inequilateral, curved, less commonly straight, with long or bent necks in whorls of (1–)2–5, occasionally solitary or paired along conidiophores, asymmetrical, only symmetrical at the conidiophore terminus, (4.2–)6.0–13.6(–19.7) × (2.2–)2.7–4.1(–4.7) μm, l/w ratio (1.4–)1.6–4.3(–5.9), (1.3–)1.6–2.7 μm wide at base, widest around the middle. Conidia ellipsoidal to oval, sometimes oblong, light green, smooth, 4.2–5.8 × (3.2–)3.5–4.2(–4.4) μm, l/w ratio 1.1–1.6. Chlamydospores generally globose to subglobose, smooth, terminal, 5.6–9.3 × 5.5–9.1 μm.
Culture characteristics: Colony radius on CMD after 72 h: 62 mm at 25 °C, 58 mm at 30 °C, 10 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies hyaline, mycelium extend from the inoculation plug, forming a reticulum. Aerial hyphae scant, inconspicuous. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 44 mm at 25 °C, 46 mm at 30 °C, 15 mm at 35 °C, covering the plate after 6 days at 25 °C. Colonies white, but greenish yellow around the inoculation plug, zonate, dense. Aerial hyphae abundant, hairy to floccose, a large green disk around the inoculums. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 63 mm at 25 °C, 16 mm at 35 °C, covering the plate after 3 days at 30 °C. Colonies hyaline, loose, radial, indistinctly zonate. Aerial hyphae white, abundant on the colony middle. Pustules minute, dispersedly distributed, firstly white, gradually turning green after 3 days. No diffusing pigment noted, odor indistinct.
Notes: Phylogenetic analyses showed that
Trichoderma inaequilaterale is the closely related species to
T. crassum Bissett and
T. virens J.H. Miller et al. Morphologically,
T. inaequilaterale can be distinguished from
T. crassum by the size of the phialides (4.2–)6.0–13.6(–19.7) × (2.2–)2.7–4.1(–4.7) μm, whereas the phialides of
T. crassum are (8.5–)13.5–15.7(–28.0) × (3.3–)4.3–4.6(–5.7) μm [
61]. Moreover,
T crassum is unable to grow at 35 °C.
T. inaequilaterale grows faster on SNA than on PDA, the same as
T. virens, but
T. virens produces yellow-diffusing pigments on CMD and PDA while
T. inaequilaterale does not [
62].
Trichoderma inconspicuum Z.F. Yu & X. Du, sp. nov. (
Figure 10)
MycoBank MB830639
Etymology. Latin, inconspicuum, meaning not remarkable, not striking, referring to the conidiophores and pialides growing in pure cultures.
Type: CHINA, Yunnan province, Jianchuan county, Jian Lake, N26°28′5.16″, E99°55′4.88″, 2182 m alt., endophytic in living stem of Nymphoides peltatum, July 2014, Z.F. Yu, Y. Huang. Holotype YMF 1.04623, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19159.
Conidiophores difficult to distinguish between mycelium, tree-like or irregular branches, the branches usually terminating in a whorl of 3–4 phialides, 2.7–4.3 μm wide at the base of branches. Phialides commonly narrow lageniform, some swollen in the middle, ampulliform, the neck usually curved, 6.4–14.4(–16.2) × 1.9–4.0 μm, l/w ratio 1.9–6.5. Conidia smooth, thin-walled, mostly subglobose to globose, a few ovoid, pale yellow-green, smooth, 2.8–4.5 × 2.6–4.0 μm, l/w ratio 1.0–1.4. Chlamydospore common, globose, ellipsoidal, smooth, terminal and intercalary, 6.6–10.1 × 5.8–8.7 μm.
Culture characteristics: Optimum temperature for growth 25 °C. No growth at 30 °C in CMD, PDA and SNA. Colony radius on CMD after 72 h: 27–32 mm at 25 °C, covering the plate after 6 days at 25 °C. Colonies hyaline, margin not well-defined. Aerial hyphae nearly lacking. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 48–53 mm at 25 °C, covering the plate after 6 days at 25 °C. Colonies white, flat, zonate indistinctly. Aerial hyphae powdery, hard to picking, sparse on outer layer, relatively compact on inner layer. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 29 mm at 25 °C, covering the plate after 7 days at 25 °C. Colonies similar to that on CMD. No diffusing pigment noted, odor indistinct.
Notes:
Trichoderma inconspicuum is phylogenetically close to
T. paraviridescens Jaklitsch et al., but their differences in morphology are distinct. The conidiophores of
T. paraviridescens are typically dendriform, with unpaired and right angle arranged branches, and the phialides is straight lageniform or sometimes curved. However, the phialides of
T. inconspicuum are hard to distinguish when grow, they are sparsely distributed, usually longer than
T. paraviridescens (7.3–10.3 μm), and the conidia of
T. paraviridescens [(3.3–)3.7–4.3(–4.7) μm] are longer than those of
T. inconspicuum [
60]. Morphologically,
T. inconspicuum is somewhat similar to
T. viridialbum in having globose to subglobose conidia, but no pustules were observed in
T. inconspicuum [
60].
Trichoderma insigne Z.F. Yu & X. Du, sp. nov. (
Figure 11)
MycoBank MB825464
Etymology. Latin, insigne, meaning distinguished, remarkable.
Type: CHINA, Yunnan province, Luliang county, N24°57′22″, E103°46′30″, 1800 m alt., from soil of tobacco rhizosphere, July 2007, Z.F. Yu. Holotype YMF 1. 00207, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19080.
Conidiophores pyramidal, with a whorl of phialides at the top, followed by short paired branches in right angles, each with a terminal whorl of phialides, the distance between two neighboring branches (15.0–)16.0–25.0(–27.0) μm, base not well-defined, about 3.0–4.0 μm wide. Phialides lateral or terminal, ampulliform or subglobose, less lageniform, widest mostly in or below the middle, in whorls, with a curved neck, (3.0–)4.0–5.0 × 3.0–4.0 μm, l/w ratio (1.1–)1.2–1.7(–1.8). Conidia commonly ellipsoidal to ovoid, rarely globose, thin-walled, green, smooth, (3.5–)3.7–4.6(–4.7) × (2.5–)2.6–3.5(–3.6) μm, l/w ratio (1.1–)1.2–1.6(–1.7).
Culture characteristics: Optimum temperature for growth 25 °C. No growth at 35 °C in PDA, CMD, and SNA. Colony radius on CMD after 72 h: 45–54 mm at 25 °C, 17 mm at 30 °C, covering the plate after 5 days at 25 °C. Colonies translucent, thin, margin visible, radial, indistinctly zonate. Aerial hyphae loosely disposed, inconspicuous. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 13 mm at 30 °C, covering the plate after 3 days at 25 °C. Colonies circular, zonate distinctly, white in outermost layer, green in innermost layer. Aerial hyphae abundant, cottony, disposed in one or two concentric rings around the point of inoculation in different color. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 55–65 mm at 25 °C, 7 mm at 30 °C, covering the plate after 4 days at 25 °C. Colonies hyaline, loose, circular, radial, margin not well-defined, indistinctly zonate. Aerial hyphae inconspicuous. Conidiation formed in pustules after 2 days, pustules spreading in 1–2 irregular concentric rings, commonly hemispherical around the point of inoculation, first white, turning green after 3 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Additional specimens examined: CHINA, Yunnan province, Luliang county, N24°57′22″, E103°46′30″, 1800 m alt., from soil of tobacco rhizosphere, July 2007, Z.F. Yu, living culture YMF 1.00272, YMF 1.00351 = CGMCC 3.19084.
Notes:
Trichoderma insigne is phylogenetically close to
T. hamatum. Morphologically,
T. hamatum is similar to
T. insigne for their densely clustered and ovoid phialides [
61]. However, the apical and secondary branches of conidiophores of
T. hamatum are sterile in elongation, for
T. insigne, these structures are abundant. In addition, the phialides of
T. insigne are significantly shorter than those of
T. hamatum [(3.0–)4.0–5.0 vs. (4.7–)5.2–8.5 μm].
Trichoderma obovatum Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 12)
MycoBank MB 834570
Etymology. Latin, ob-, meaning against, contrary, + ovatum, meaning ovoid shape, like an egg, referring to the conidia.
Type: CHINA, Yunnan province, Jianshui county, N23°20′22″, E103°06′57″, 1170 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06211, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021069.
Conidiophores commonly straight, sometimes curved, emerging in right angles or oriented towards to the conidiophore axis, typically with 1–2 branched levels, side branches simple. Phialides ampuliform to lageniform, sometimes nearly round, mostly inhomogeneous, commonly curved or sinuous, with narrow cylindrical or hooked necks, formed mostly around the stipes at regular levels in whorls of 1–4, some growing directly on the main axis in whorls of 1–3, (4.0–)4.8–8.9(–10.7) × (2.3–)2.5–3.9(–4.2) μm, l/w ratio (1.0–)1.4–3.7(–4.3), 1.4–2.5(–3.2) μm wide at base. Conidia globose, oval to obovate, less commonly oblong, green, smooth, 3.2–3.8 × 3.0–3.6 μm, l/w ratio 1.0–1.2.
Culture characteristics: Optimum temperature for growth 25 °C. Colony radius on CMD after 72 h: 38–41 mm at 25 °C, 28 mm at 30 °C, covering the plate after 5 days at 25 °C, attaining 1–2 mm after 7 days at 35 °C. Colonies translucent, margin dense, center loose, inconspicuously zonate. Aerial hyphae short. Pustules firstly white, turning green after 3 days, concentrated in a marginal zone, sparse in the middle. Chlamydospores unobserved. Especially pleasant odor noted. No diffusing pigment noted. Colony radius on PDA after 72 h: cover the plate at 25 °C, 22–24 mm at 30 °C, 3–4 mm at 35 °C. Colonies grayish yellow-green to pistachio green alternately, well-defined, dense, floccose, homogeneous, indistinctly zonate. Aerial hyphae abundant, forming a dense mat. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 25–27 mm at 25 °C, 16–19 mm at 30 °C, covering the plate after 6 days at 25 °C, attaining 1–2 mm 35 °C after 7 days. Colonies hyaline, irregularly circular, indistinctly zonate. Aerial hyphae sparse, inconspicuous, mycelium creeping on the surface of media. Pustules firstly white, turning green after 3 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Additional specimen examined: CHINA, Yunnan province, Jianshui county, N23°20′22″, E103°06′57″, 2018 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv, living culture YMF 1.06212.
Notes:
Trichoderma obovatum is characterized by its distinct odor and the rather dense pustules on CMD. Phylogenetic analyses reveal that
T. obovatum is related to
T. paratroviride Jaklitsch & Voglmayr, and they share the same optimal growth temperature. However, the colonies of
T. paratroviride are slightly yellowish in reverse and chlamydospores were observed, whereas they were not observed in
T. obovatum. Furthermore, the phialides of
T. paratroviride are longer than those of
T. obovatum [(5.2–)6.2–11(–14) vs. (4.0–)4.8–8.9(–10.7) um] [
46].
Trichoderma paraviride Z. F. Yu & X. Du, sp. nov. (
Figure 13)
MycoBank MB830640
Etymology. Latin, para-, meaning like, resemble, -viride, referring to the species Trichoderma viride.
Type: CHINA, Sichuan province, Litang county, N30°4′12.4″, E100°7′5.1″, 4029 m alt., endophytic in living root of Elodea nuttallii in pond, July 2014, Z.F. Yu, Y. Huang, Holotype YMF 1.04628, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19160.
Conidiophores ill-defined, variably curved to sinuous extension, rarely main axis conspicuously discernible, the primary branches arising at narrow angle from the main axis, toward the tip, crowded and arranged irregularly, the secondary branches generally paired in a tree fashion, all branches terminating in a whorl of 3–5 phialides, the distance between two neighboring branches 8.6–30.1 μm, bases noted distinctly, about 2.3–3.9 um wide. Phialides commonly spindly to ellipsoid, rarely cylindrical, sometimes curved, the necks of phialides typically sinuous, sometimes not visible, (9.2–)9.4–15.5(–16.4) × (2.1–)2.2–3.8(–4.1) μm, l/w ratio (2.6–)2.7–5.3(–6.6). Conidia thin-walled, globose to subglobose, smooth, green, smooth, (3.4–)3.6–4.9(–5.0) × 3.2–4.6(–4.8) μm, l/w ratio 1.0–1.3.
Culture characteristics: Optimum temperature for growth 25 °C. No growth at 35 °C in PDA and SNA. Colony radius on CMD after 72 h: 47–51 mm at 25 °C, 43 mm at 30 °C, 4 mm at 35 °C, covering the plate after 5 days at 25 °C. Colonies hyaline, thin, margin slightly lobed, not well-defined. Aerial hyphae inconspicuous. Pustules commonly pulvinate, less hemispherical, forms relatively less, firstly white, turning green after 5 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 70 mm at 25 °C, 39 mm at 30 °C, covering the plate after 4 days at 25 °C. Colonies circular, white, margin wavy and surface flat. Aerial hyphae dense, continuous, arachnoid. Chlamydospores unobserved. No diffusing pigment noted. A distinct coconut-like odor noted. Colony radius on SNA after 72 h: 56 mm at 25 °C, 39 mm at 30 °C, covering the plate after 5 days at 25 °C. Colonies thin, hyaline. Aerial hyphae inconspicuous, sparse. Pustules abundant, compact, asymmetrical to hemispherical, loosely arranged in outer layer, appear within 6 days, turning green from after 7 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Notes: Phylogenetic analyses show that
Trichoderma paraviride belongs to the
Viride clade and is close to
T. viride Fries. Morphologically,
T. viride is similar to
T. paraviride in the branching pattern of conidiophore and conidia shape; whereas the branches of
T. viride arise from the main axis at a wide angle, the phialides of
T. viride are lageniform to long cylindrical [
46].
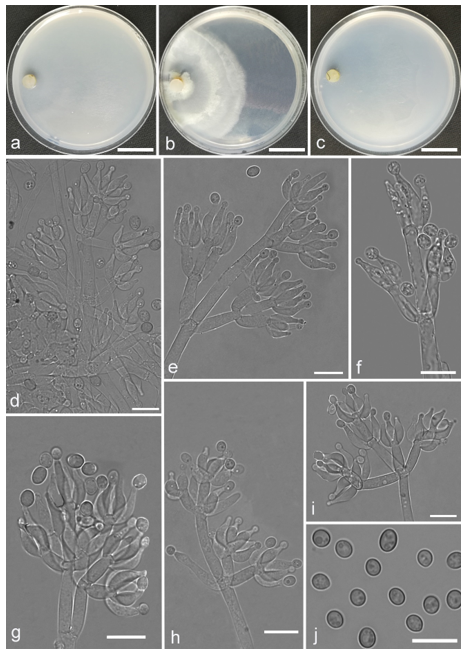
Trichoderma pluripenicillatum Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 14)
MycoBank MB 834554
Etymology. Latin, pluri-, meaning several, many, -penicillatum, referring to the penicillate arrangement of the phialides.
Type: CHINA, Yunnan province, Jianshui county, N23°20′07″, E103°06′32″, 2017 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv, Holotype YMF 1.06198, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021062.
Conidiophores straight or slightly curved, gliocladium-like, with sparingly branching structures from the central axis, unpaired, irregular, typically asymmetrical branching of 4–6 branches in verticils in steep angles. Branches clung to the conidiophore terminus and bore ampuliform phialides that do not develop directly on the axis but aggregate at the branch terminus. Several stipes mounted on one branch terminus with a solitary phialide. Phialides ampuliform with symmetrical necks, (6.2–)6.5–9.0(–9.8) × 2.7–3.7(–4.0) μm, l/w ratio (1.9–)2.2–2.7(–3.1), 1.5–2.5 μm wide at the base, widest at the middle. Conidia oval to round, pale yellow-green, smooth, 2.7–3.4(–3.7) × (2.2–)2.5–3.2 μm, l/w ratio 1.0–1.2(–1.4). Chlamydospores roundness, smooth, terminal, 5.5–8.3 × 5.3–7.9 μm.
Culture characteristics: Optimum temperature for growth 30 °C. Colony radius on CMD after 72 h: 33 mm at 25 °C, 38 mm at 30 °C, 27 mm at 35 °C, covering the plate after 9 days at 30 °C. Colonies thin, translucent, indistinctly zonate, radial. Aerial hyphae scant, inconspicuous. Pustules minute, firstly white, finally turning downy light-green, surrounded the inoculation plug. A distinct odor noted. No diffusing pigment noted. Colony radius on PDA after 72 h: 36 mm at 25 °C, 42 mm at 30 °C, 35 mm at 35 °C, covering the plate after 7 days at 30 °C. Colonies zonation conspicuous, white to yellowish-white alternately, mycelium dense and radial, forming a continuous white lawn around the center and thinner at the margin. Aerial hyphae abundant, short. Yellow soluble pigments observed. A distinct odor noted. Colony radius on SNA after 72 h: 20 mm at 25 °C, 46 mm at 30 °C, 15 mm at 35 °C, covering the plate after 8 days at 30 °C. Colonies hyaline, mycelium sparse. Aerial hyphae scant, inconspicuous. No diffusing pigment noted, odor indistinct.
Notes:
Trichoderma pluripenicillatum is located in the
Longibrachiatum clade and distinguishable by the
gliocladium-like conidiophores, which are uncommon in
Trichoderma. Morphologically,
T. pluripenicillatum is similar to
T. gliocladium Jaklitsch & Voglmayr in the branch patterns of conidiophore and shapes of phialide, but differs in its conidia and colony characteristics:
T. pluripenicillatum formed bright yellow pigments on PDA, while
T. gliocladium produced orange-brown pigments [
46]. Phylogenetic analyses have shown that
T. pluripenicillatum is closely related to
T. citrinoviride Bissett with relatively high supports, but the latter has optimal growth at 35 °C on all media, while
T. pluripenicillatum grow better at 30 °C. Moreover,
T. citrinoviride has shorter phialides than
T. pluripenicillatum [(3.7–)4.7–7.8(–10.5) vs. (6.2–)6.5–9.0(–9.8) μm]. For the conidia shape,
T. citrinoviride produces oblong or ellipsoidal conidia, but
T. pluripenicillatum forms oval to round conidia [
63].
Trichoderma propepolypori Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 15)
MycoBank MB 834556
Etymology. Latin, prope-, meaning near, -polypori, referring to the phylogeny affinities with Trichoderma polypori.
Type: CHINA, Yunnan province, Jianshui county, N23°20′13″, E103°06′24″, 2017 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06224, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021067.
Conidiophores straight or slightly curved, comprised developed main axis and paired branches at relatively regular intervals along the central axis, branches tended towards the conidiophore terminus in steep angles. Phialides ampulliform with sinuous, often constricted below the tip to form a narrow neck, less frequently solitary, often in whorls of 2–5, mostly inequilateral, equilateral only in central whorls, (6.5–)7.3–11.5(–14.1) × (2.8–)3.5–5.0 μm, l/w (1.3–)2.0–3.8(–5.0), (1.4–)1.9–3.0(–3.8) μm wide at the base, widest around the middle. Conidia pale yellow-green, smooth, ellipsoidal or oval, spheroidal present, (3.4–)3.7–4.2(–4.6) × 3.2–4.0 μm, l/w ratio 1.0–1.3.
Culture characteristics: Optimum temperature for growth 30 °C on CMD and SNA, 25 °C on PDA. Colony radius on CMD after 72 h: 36 mm at 25 °C, 38 mm at 30 °C, 9 mm at 35 °C, covering the plate after 4 days at 30 °C. Colonies translucent, thin, flat. Aerial hyphae nearly lacking. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 55 mm at 25 °C, 32 mm at 30 °C, 9–13 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies pale white, thick, fluffy, zonate indistinctly. Aerial hyphae dense, cotton-like. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 31 mm at 25 °C, 42 mm at 30 °C, 8 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies similar to that on CMD.
Additional specimen examined: CHINA, Yunnan province, Jianshui county, N23°20′13″, E103°06′24″, 2017 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv, living culture YMF 1.06199.
Notes:
Trichoderma propepolypori is distinctive by the cotton-like aerial hyphae on PDA. Phylogenetically,
T. propepolypori is closely related to
T. polypori and
T. longifialidicum. However,
T. propepolypori can be easily distinguished by slower growth rate, shape of phialides, and size of conidia. The phialides of
T. propepolypori are sinuously ampulliform phialides, while
T. polypori is lageniform and subulate [
44]. The conidia of
T. propepolypori are much bigger than those of
T. polypori, (3.4–)3.7–4.2(–4.6) × 3.2–4.0 vs. 2.8–3.6(−4.2) × 2.5–3.3 μm. In addition,
T. longifialidicum has elongated and cylindrical phialides (7–21 × 1.4–2.8 μm) and simpler conidiophore patterns [
54].
Trichoderma pseudoasiaticum Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 16)
MycoBank MB 834559
Etymology. Latin, pseudo-, meaning like, similar, -asiaticum, referring to the species Trichoderma asiaticum.
Type: CHINA, Yunnan province, Jianshui county, N23°19′22″, E103°06′13″, 2010 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06200, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021061.
Conidiophores verticillium-like, straight or curved, mostly emerging in aerial hyphae, typically with 1–3 branching levels, branches orientating slightly towards the conidiophore terminus, the main branches often unpaired or irregular, the side branches simple, rebranching 1–2 times, replaced by a solitary phialide or paired phialides. Phialides ampulliform, sometimes with narrow cylindrical or slightly bent neck, solitary or in whorls of 2–4, symmetric or inequilateral, (5.2–)6.1–9.0(–9.7) × (2.1–)2.6–3.6(–4.0) μm, l/w ratio (1.1–)1.5–3.6(–5.2), (1.0–)1.4–2.3(–2.6) μm wide at base. Conidia green, smooth, globose or subglobose, 2.4–3.2 × 2.4–3.0 μm, l/w ratio 1.0–1.1. Chlamydospores globose, relatively common, smooth, terminal, 4.7–7.7 × 4.0–7.6 μm.
Culture characteristics: Optimum temperature for growth 25 °C. Colony radius on CMD after 72 h: 60 mm at 25 °C, 49 mm at 30 °C, 21 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies pale green, thin, circular, zonate indistinctly, radial, mycelium loose. Aerial hyphae loose, mostly concentrated on the margin. Conidiation formed in pustules after 3 days, pustules dense, firstly white, finally turning dark green. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: cover the plate at 25 °C and 30 °C, 13 mm at 35 °C. Colonies white to dark green alternately, regularly circular, indistinctly zonate, radial, mycelium dense. Aerial hyphae conspicuous. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 62 mm at 25 °C, 35 mm at 30 °C, 2 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies hyaline to light green, thin, mycelium sparse. Aerial hyphae loose. Conidiation formed in pustules after 3 days, pustules green, abundant in margin. No diffusing pigment noted, odor indistinct.
Notes:
Trichoderma pseudoasiaticum belongs to the
Harzianum clade, and is phylogenetically close to
T. guizhouense Q.R. Li, McKenzie & Yong Wang. However,
T. pseudoasiaticum is obviously different from
T. guizhouense in culture characteristics on three media and phialides. The colonies of
T. pseudoasiaticum are white to light green and no diffusing pigment was observed, whereas those of
T. guizhouense are white and produce brown diffusing pigmentation on PDA. Furthermore, the phialide of
T. pseudoasiaticum is wider than that of
T. guizhouense (2.6–3.6 vs. 2.0–3.0 μm) [
64].
Trichoderma pseudoasperelloides Z.F. Yu & X. Du, sp. nov. (
Figure 17)
MycoBank MB825463
Etymology. Latin, pseudo-, meaning like, similar, -asperelloides, referring to the species Trichoderma asperelloides.
Type: CHINA, Sichuan province, Xichang county, Qionghai Lake, N27°83′77.36″, E102°28′2.98″, 1520 m alt., endophytic in living leaf of Myriophyllum spicatum, July 2014, Z.F. Yu, Y. Huang. Holotype YMF 1.04629, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19078.
Conidiophores dendriform, little rebranching, often terminating in whorls of 3–5 phialides, rarely solitary, base well-defined, inconspicuous near the tip of main axis, about 3.0–5.0 um wide, branches commonly opposite, each branches terminating in a whorl of 3 or 4, the distance between two neighbor branches (14.0–)18.0–36.0(–37.0) μm. Phialides spindle-shaped and lageniform, (5.0–)6.0–8.0(–9.0) × (2.0–)3.0–4.0(–5.0) μm, l/w ratio (1.3–)1.4–2.7(–3.0). Conidia thin-walled, subglobose to ellipsoidal, rarely globose, green, smooth, (3.7–)3.8–4.0(–4.1) × (2.7–)2.8–3.3(–3.8) μm, l/w ratio (1.0–)1.1–1.4(–1.5).
Culture characteristics: Colony grows very fast on CMD and PDA. Colony radius on CMD after 72 h: cover the plate at 25 °C, 30 °C, and 35 °C. Colonies translucent, zonate indistinctly. Aerial hyphae scant, radial, inconspicuous. Pustules firstly white, gradually turning green after 3 days, relatively rare in middle, abundant at the margin. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: cover the plate at 25 °C, 30 °C, and 35 °C. Colonies pale green to dark green alternately, layered distinctly, margin conspicuous, zonate indistinctly. Aerial hyphae hairy to floccose, dense, abundant in a large green disk around the inoculum. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 55–61 mm at 25 °C, 53 mm at 30 °C, 42–47 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies hyaline, circular, indistinctly zonate, mycelium sparse. Aerial hyphae sparse. Pustules dark green, relatively rare in the middle, abundant in the margin. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Additional specimens examined: CHINA, Sichuan province, Xichang county, Qionghai Lake, N27°83′77.36″, E102°28′2.98″, 1520 m alt., endophytic in living leaf of Myriophyllum spicatum. July 2014, Z.F. Yu, Y. Huang, living culture YMF 1.04633; CHINA, Yunnan province, Luliang county, N24°57′22", E103°46′30", 1800 m alt., from soil of tobacco rhizosphere, July 2007, Z.F. Yu, living culture YMF 1.00258 = CGMCC 3.19081, YMF 1.00378, YMF 1.00152.
Notes:
Trichoderma pseudoasperelloides is phylogenetically close to
T. asperelloides Samuels. They are similar in shape of the phialides and conidia. But for
T. asperelloides, secondary branches tend to be paired, also commonly unilateral and consist of a single cell near the tip of the conidiophore, which cannot be represented in
T. pseudoasperelloides for its fascicular phialides near the tip [
65].
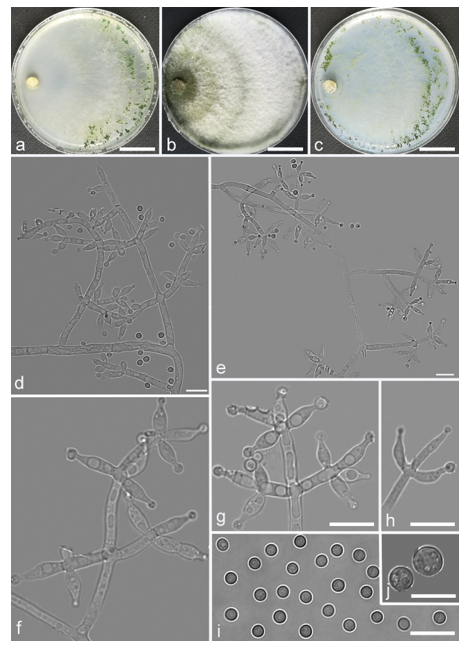
Trichoderma scorpioideum Z.F. Yu & X. Du, sp. nov. (
Figure 18)
MycoBank MB830638
Etymology. Latin, scorpioideus, referring to the scorpioid arrangement of the branches and phialides.
Type: CHINA, Sichuan province, Daocheng county, N29°29′48.5″, E100°14′46.5″, 4362 m alt., endophytic on living stem of Hippuris vulgaris in pond, July 2014, Z.F. Yu, Y. Huang. Holotype YMF 1.04616, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19623.
Conidiophores straight or curved, comprised a slightly curved main axis and generally verticillate branches, the main axis often terminating in a whorl of 2–3 divergent phialides, the base of branches about 2.1–4.3 μm wide, branches generally toward the top of main axis and sometimes sterile, terminating 1–3 divergent phialides, the distance between two neighbor branches 12.8–18.8 μm. Phialides commonly narrow lageniform, some ellipsoidal, slightly swollen in the middle, the necks of phialides sometimes curved, sometimes arose singly from the main axis or branches, (6.5–)6.8–12.7(–13.3) × 2.0–3.9 μm, l/w ratio (1.8–)2.1–4.5(–4.7). Conidia commonly globose to subglobose, a few ovoidal to ellipsoidal, hyaline, thin-wall, green, smooth, 3.3–4.4 × 2.4–3.8 μm, l/w ratio 1.0–1.7.
Culture characteristics: Optimum temperature for growth 25 °C. No growth at 35 °C in PDA and SNA. Colony radius on CMD after 72 h: 46–54 mm at 25 °C, 32–36 mm at 30 °C, 18–20 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies translucent, thin, flat, radial, mycelium loose. Aerial hyphae sparse, inconspicuous radiate. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 55–61 mm at 25 °C, 31–37 mm at 30 °C, covering the plate after 4 days at 25 °C. Colonies white, inconspicuous radial, mycelium dense. Aerial hyphae numerous, long and wooly. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 48–53 mm at 25 °C, 27–32 mm at 30 °C, covering the plate after 4 days at 25 °C. Colonies translucent, thin, radial. Aerial hyphae sparse, arachnoid, arranged in a scattered fashion. Pustules white, minute, abundant in the margin. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Notes:
Trichoderma scorpioideum is phylogenetically closely related to two species:
T. viridescens and
T.
sempervirentis Jaklitsch & Voglmayr, the branches of
T. scorpioideum and
T. viridescens are slightly curved, whereas the branches of
T. viridescens is paired and often terminated in 1 or 2 phialides [
59], which distinctly differs from verticillate branches of
T. scorpioideum. As for the phialides of
T. viridescens, it often forms a submoniliform chain of 2–6 cells when cultured on CMD [
59], in contrast, the structure is inconspicuous on the phialides of
T. scorpioideum. Furthermore, the conidia of
T. viridescens sometimes are wider (3.0–3.7 μm) than those of
T. scorpioideum.
Trichoderma simile Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 19)
MycoBank MB 834560
Etymology. Latin, simile, meaning like, resemblance, which this species has resemblance to Trichoderma guizhouense.
Type: CHINA, Yunnan province, Jianshui county, N23°17′24″, E103°06′32″, 2015 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06201, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021064.
Conidiophores tree-like, formed densely intricate reticulum, main axis unrecognizable, mostly curved, integrated into the reticulum, side branches arising from main axis asymmetrically, perpendicular to the axis, some slightly orientated towards the conidiophore terminally, rebranching 1–3 times. Phialides varied, borne in regular levels around the axis, mostly paired arrangements or in whorls of 2–5, sometimes crowded at the stipe terminus, less commonly singly, straight or curved, ampulliform, less lageniform with long, symmetrical or slightly bent necks, (3.8–)4.3–11.9(–14.3) × (2.3–)2.7–3.9 μm, l/w ratio 1.3–4.4(–5.2), 1.5–2.8(–3.6) μm wide at base, widest around the middle. Conidia oval, elliptic to subspheroidal, less oblong, green, smooth, 2.6–3.2 × 2.2–2.8 μm, l/w ratio 1.0–1.2. Chlamydospores elliptic or round, smooth, terminal and intercalary, 4.2–7.8 × 4.0–7.2 μm.
Culture characteristics: Optimum temperature for growth 25 °C. Colony radius on CMD after 72 h: 47 mm at 30 °C, 6 mm at 35 °C, cover the plate after 3 days at 25 °C. Colonies translucent, thin, radial. Aerial hyphae sparse. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 63 mm at 25 °C, 60 mm at 30 °C, 10 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies pale green to greyish green alternately, a few flesh-colored areas occurred on the medium, circular, conspicuously zonate. Aerial hyphae abundant, firstly white, alternately forming pale green to grayish green lawn. Pustules formed along the margin. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 47 mm at 25 °C, 45 mm at 30 °C, 3 mm at 35 °C, covering the plate after 4 days at 25 °C. Colonies translucent, thin, radial, conspicuously zonate. Aerial hyphae loose, sparse in the middle, relatively abundant in the margin. Conidiation formed on pustules, pustules minute, relatively abundant in the zonation regions. No diffusing pigment noted, odor indistinct.
Additional specimen examined: CHINA, Yunnan province, Jianshui county, N23°17′24″, E103°06′32″, 2015 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv, living culture YMF 1.06202.
Notes: Phylogenetically,
Trichoderma simile is closely related to
T. guizhouense. However,
T. simile is distinguished from
T. guizhouense by producing chlamydospores. Moreover, there are significant differences in size and shape of phialides and conidia, for instance, the phialides of
T. guizhouense, (4.5–10 × 2.0–3.0 μm), are narrower than
T. simile, (3.8–)4.3–11.9(–14.3) × (2.3–)2.7–3.9 μm, and
T. guizhouense has globose conidia while the conidia of
T. simile are oval [
64].
Trichoderma subazureum Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 20)
MycoBank MB 834565
Etymology. Latin, subazureum, meaning pale bluish, referred to the color of the colonies on CMD, and SNA media.
Type: CHINA, Yunnan province, Jianshui county, N23°17′34″, E103°06′28″, 2015 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06207, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021060.
Conidiophores straight or slightly curved, typically comprising 1–2 levels branched with phialides arising at the top in whorls of 2–4(–5), less commonly solitarily along stipes, with side branches short at the main axis terminus and much longer subjacent, branches paired or in whorls of 1–3, disposed perpendicular to the axis, some orientating slightly towards the conidiophore terminus. Phialides ampulliform to lageniform, with symmetrical or slightly curved necks, (4.7–)5.2–11.9(–20.6) × (2.2–)2.7–3.6(–4.2) μm, l/w ratio (1.3–)1.7–4.9(–8.5), (2.0–)2.2–2.9(–3.5) μm wide at base, widest around the middle. Conidia ellipsoidal to oblong, less globose, green, smooth, (3.2–)3.5–4.4(–4.9) × (2.1–)2.4–2.9 μm, l/w ratio (1.1–)1.4–1.8.
Culture characteristics: Optimum temperature for growth 25 °C. Colony radius on CMD after 72 h: 45 mm at 25 °C, 38 mm at 30 °C, attaining 5 mm after 7 days at 35 °C, covering the plate after 5 days at 25 °C. Colonies translucent, circular with wavy margin, zonate inconspicuously, mycelium radial, denser at the zonation rings. Aerial hyphae scant, reticular. Pustules white, minute, sparse, relatively abundant in margin. Chlamydospores unobserved. Light yellow pigments noted. No indistinct odor. Colony radius on PDA after 72 h: 60 mm at 25 °C, 47 mm at 30 °C, attaining 5 mm at 35 °C after 7 days, covering the plate after 4 days at 25 °C. Colonies white to pale bluish green, downy, thick, dense, finely wavy hyphae. Aerial hyphae abundant, dense. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 45 mm at 25 °C, 39 mm at 30 °C, attaining 3 mm at 35 °C after 7 days, covering the plate after 5 days at 25 °C. Colonies translucent to pale bluish green, fluffy, villous. Aerial hyphae dense, slightly coiling. Conidiation formed on pustules after 4 days, pustules minute, dense, relatively abundant in margin, firstly white, turning green after 7 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Notes:
Trichoderma subazureum is characterized by crowded pustules and relatively larger conidia. Phylogenetically, it is closely related to
T. spirale Bissett. However,
T. subazureum is obviously different from
T. spirale in morphology by having longer phialides, (4.7–)5.2–11.9(–20.6) vs. (3.2–) 4.5–6.5 (–8.2) μm [
61]. Moreover, the growth rate of
T. spirale, e.g. 60–70 mm on SNA after 3 d at 30 °C, is faster than that of
T. subazureum.
Trichoderma subuliforme Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 21)
MycoBank MB 834564
Etymology. Latin, subuliforme, meaning subulate, awl-shaped, referring to the phialides.
Type: CHINA, Yunnan province, Jianshui county, N23°15′16″, E102°57′13″, 1170 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06204, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021059.
Conidiophores verticillium-like, phialides not formed directly around the axis, side branches emerging from the main axis, perpendicular relative to the stipe axis, or orientating slightly towards the conidiophore terminus, often paired or in verticils, also solitary, mostly straight. Secondarybranches appear, no tertiary branches noted. Phialides subuliform, sometimes long lageniform with long neck, symmetrical or slightly curved, in whorls of 3–5(–6), (3.7–)7.0–15.8(–19.4) × (1.9–)2.4–3.0(–3.4) μm, l/w ratio (1.2–)2.4–5.9(–6.6), 1.9–3.0 μm wide at base. Conidia oblong to ellipsoidal, less subspheroidal, green, smooth, (2.9–)3.2–3.9(–4.3) × 2.1–2.7(–3.4) μm, l/w ratio 1.2–1.6.
Culture characteristics: Optimum temperature for growth 25 °C. Colony radius on CMD after 72 h: 45 mm at 25 °C, 43 mm at 30 °C, attaining 23 mm at 35 °C after 7 days, covering the plate after 4 days at 25 °C. Colonies translucent, circular, loose, mycelium dense. Aerial hyphae abundant, downy. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 54 mm at 25 °C, 31 mm at 30 °C, attaining 4 mm at 35 °C after 7 days, covering the plate after 4 days at 25 °C. Colonies white to grayish yellow-green alternately, zonate inconspicuously, flocky, mycelium dense. Aerial hyphae abundant, forming a dense flat. Yellow pigments noted. No indistinct odor noted. Colony radius on SNA after 72 h: 34 mm at 25 °C, 24 mm at 30 °C, attaining 9 mm at 35 °C after 7 days, covering the plate after 6 days at 25 °C. Colonies firstly white, gradually turning grayish yellow-green, zonate, circular, mycelium dense. Aerial hyphae common. Pustule occurred on the zonate margin, firstly white, gradually turning dark green. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Additional specimens examined: CHINA, Yunnan province, Jianshui county, N23°15′16″, E102°57′12″, 1170 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv, living culture YMF 1.06205, YMF 1.06206.
Notes:
Trichoderma subuliforme is phylogenetically closed to
T. spirale. In morphology,
T. subuliforme has slenderer phialides than
T. spirale, (2.9–)3.2–3.9(–4.3) × 2.1–2.7(–3.4) vs. (3.6–)4.2–5.6 (−7.5) × 3.1–3.9 μm [
44].
Trichoderma supraverticillatum Z.F. Yu & Y.F. Lv, sp. nov. (
Figure 22)
MycoBank MB 834566
Etymology. Latin, supra-, meaning above, over, -verticillatum, meaning verticillate, referring to the phialides.
Type: CHINA, Yunnan province, Jianshui county, N23°15′22″, E102°57′28″, 1170 m alt., from soil, June 2018, Z.F. Yu, Y.F. Lv. Holotype YMF 1.06208, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CCTCC AF 2021063.
Conidiophores regularly tree-like, side branches from main axis typically paired, perpendicular to the axis at irregular intervals, some slightly orientating towards the conidiophore terminus. Stage 2 branches present, no stage 3 branches noted. Phialides formed mostly around the stipes at regular levels, in whorls of 3–4, paired or solitary directly on the main axis, ampuliform to spindly, homogeneous, straight, less curved or sinuous, with symmetrical or slightly bent long necks, (5.5–)7.5–11.5(–13.6) × (2.2–)2.5–3.0(–3.5) μm, l/w ratio (1.6–)2.7–4.6(–5.7), (1.9–)2.2–3.0(–3.3) μm wide at base. Conidia variable with two different types, ellipsoidal, less subglobose, or obpyriform to obovoid with an apparent protuberance, green, smooth, (3.2–)3.5–4.0(–4.6) × (2.3–)2.6–3.4(–3.8) μm, l/w ratio (1.1–)1.3–1.6(–1.8). Chlamydospores ellipsoidal, smooth, uncommon, terminal and intercalary, 5.3–8.1 × 5.0–7.9 μm.
Culture characteristics: Optimum temperature for growth 25 °C on CMD and SNA, 30 °C on PDA. Colony radius on CMD after 72 h: 49 mm at 25 °C, 43 mm at 30 °C, attaining 5 mm at 35 °C after 7 days, covering the plate after 5 days at 30 °C. Colonies translucent, circular, radial, zonate inconspicuously, mycelium scant. Aerial hyphae virtually scarce, relatively abundant at the margin, slightly coiling. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 45 mm at 25 °C, 53 mm at 30 °C, attaining 5 mm at 35 °C after 7 days, covering the plate after 5 days at 30 °C. Colonies white, circular, first hairy, later velutinous, forming a 15-mm-wide translucent ring around the inoculation plug, mycelium abundant. Aerial dense, fluffy, downy texture. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 51 mm at 25 °C, 46 mm at 30 °C, attaining 8 mm at 35 °C after 7 days, covering the plate after 5 days at 25 °C. Colonies hyaline. Aerial hyphae scant, relatively abundant at the margin. No diffusing pigment noted, odor indistinct.
Notes: Trichoderma supraverticillatum forms a sole clade with relatively high statistical support. It can be distinguished by two types of conidia, which are ellipsoidal or obpyriform to obovoid with an apparent protuberance. Morphologically, T. supraverticillatum is similar to T. subuliforme with characteristics of conidiophores and phialides. However, T. subuliforme has oblong or ellipsoid conidia, and smaller than those of T. supraverticillatum, (3.2–)3.5–4.0(–4.6) × (2.3–)2.6–3.4(–3.8) vs. (2.9–)3.2–3.9(–4.3) × 2.1–2.7(–3.4) μm.
Trichoderma tibetica Z.F. Yu & X. Du, sp. nov. (
Figure 23)
MycoBank MB825464
Etymology. Latin, tibeticum, referring to the sampling site Tibet.
Type: CHINA, Tibet, Gongjue county, Xiongba village, N32°32′7.74″, E82°31′13.42″, 4475 m alt., endophytic on living stem of Batrachium bungei, August 2018, Z.F. Yu, X. Du. Holotype YMF 1.05583, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19628.
Conidiophores consist of a discernible, slightly curved main axis and generally paired branches, the main axis usually terminated in three cruciform phialides, the branches often slightly upward and sometimes perpendicular to main axis, the distance between two neighbor branches 10.5–34.3 μm, base slightly swollen, about 2.4–3.6 μm wide, terminating in a whorl of 2–4 divergent phialides. Phialides usually lageniform, sometimes globose, ellipsoidal, or pyramidal, the neck of phialides sometimes hooked or degenerated, phialides around the tip sometimes arose singly from the main axis, (5.2–)5.9–11.8(–12.3) × (2.1–)2.3–3.5 μm, l/w ratio1.9–4.5(–4.9). Conidia ellipsoidal to ovoid, subglobose rarely noted, green, smooth, 3.7–4.8 × (2.9–)3.1–4.0 μm, l/w ratio 1.1–1.4.
Culture characteristics: Optimum temperature for growth 25 °C. No growth at 35 °C on CMD, PDA and SNA. Colony radius on CMD after 72 h: 56–58 mm at 25 °C, 56 mm at 30 °C, covering the plate after 4 days at 30 °C. Colonies hyaline, radial, thin. Aerial hyphae sparse. Chlamydospores unobserved. Light pigments noted. No distinct odor noted. Colony radius on PDA after 72 h: cover the plate after 3 days at 25 °C and 30 °C. Colonies white, radial, flat, mycelium abundant. Aerial hyphae dense, floccose. Chlamydospores unobserved. Yellow pigments noted. No distinct odor noted. Colony radius on SNA after 72 h: 29–37 mm at 25 °C, 29 mm at 30 °C, covering the plate after 6 days at 25 °C. Colonies translucent, thin, mycelium relatively sparse around the inoculation plug. Aerial hyphae loose. Chlamydospores unobserved. Light yellow pigments noted. No distinct odor noted.
Notes:
Trichoderma tibetica phylogenetically belongs to the
Koningii clade, and is closely related to
T.
petersenii Samuels, Dodd & Schroers
. T. tibetica and
T. petersenii showed no significant difference in the morphological features of conidiophores, which both species having a well-defined main axis and generally paired branches, and the phialides of two species being lageniform, sometimes cylindrical, and slightly swollen in the middle [
55]. But the phialides of
T. tibetica are sometimes globose or pyramidal, and sometimes hook-like on the bent neck, which is rarely noted in
T.
petersenii. The conidia of
T. tibetica are mostly ellipsoidal, sometimes ovoidal or subglobose, which distinguishes them from the generally ellipsoidal conidia of
T.
petersenii, and the conidia of
T.
petersenii are commonly smaller than those of
T. tibetica (3.5–4.5 × 2.7–3.0 μm). In addition, no conidia formed on CMD for
T. tibetica, only vegetative hyphae and pigment were obviously noted, but the conidia of
T.
petersenii appeared within 7 days on CMD [
55].
Trichoderma uncinatum Z.F. Yu & X. Du, sp. nov. (
Figure 24)
MycoBank MB830641
Etymology. Latin, uncinatus, referring to the hooked, uncinated neck of the phialides.
Type: CHINA, Guizhou province, Huaxi county, N26°26′31.96″, E106°40′27.15″, 1081 m alt., endophytic on living stem of Potamogeton malaianus in wetland, July 2014, Z.F. Yu, Y. Huang. Holotype YMF 1.04622, preserved in a metabolically inactive state (deep freezing) in the Conservation and Utilization of Bio-Resources in Yunnan. Ex-type culture CGMCC 3.19621.
Conidiophores comprised a hard-discernable, slightly curved main axis, irregular alternate branches, the distance between two neighbor branches of 11.2–26.2 μm, base sometimes swollen, about 2.6–4.2 μm wide, the main axis often terminating in two divergent phialides, every branch often terminating in a whorl of 2–5 phialides. Phialides commonly lageniform, sometimes ampulliform to subglobose, sometimes sterile, sometime the neck of phialides uncinate or constricted sharply, rarely single, (4.3–)5.2–9.3(–10.3) × 2.3–3.9 μm, l/w ratio 1.3–3.6. Conidia globose to subglobose, rarely ovoidal or ellipsoidal, thin-walled, green, smooth, 3.1–4.4 × 2.7–4.0 μm, l/w ratio 1.0–1.4.
Culture characteristics: Optimum temperature for growth 30 °C on CMD and SNA, 25 °C on PDA. No growth at 35 °C on CMD, PDA and SNA. Colony radius on CMD after 72 h: 55–62 mm at 25 °C, 61–68 mm at 30 °C, covering the plate after 4 days at 30 °C. Colonies white, thin, flat. Aerial hyphae relatively scant. Pustules minute, dispersedly distributed, abundant in the margin, white at first, turning green after 5 days. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on PDA after 72 h: 61 mm at 30 °C, covering the plate after 3 days at 25 °C. Colonies white, loose, coarse. Aerial hyphae abundant, arachnoid. Pustules minute, abundant around the inoculation plug, firstly white, gradually turning dark green. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct. Colony radius on SNA after 72 h: 43–49 mm at 25 °C, covering the plate after 3 days at 30 °C. Colonies white, circular, zonate inconspicuously. Aerial hyphae abundant. Pustules minute, formed after 3 days, turning green after 5 days, mostly distributed around the inoculation plug. Chlamydospores unobserved. No diffusing pigment noted, odor indistinct.
Notes:
Trichoderma uncinatum is close to
T. paratroviride in the phylogenetic tree. Morphologically, they share some characters, such as spindly branches generally toward the tip, lageniform to ampulliform phialides sometimes with curved necks, and subglobose conidia [
46]. However, the distance between two neighboring branches in
T. paratroviride is generally longer, the phialides of
T. paratroviride are commonly in whorls of 2–4 vs. in whorl of 2–5 in
T. uncinatum, and the phialides are spindlier (6.2–11 × 2.5–3.2 μm) than in
T. uncinatum.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
























