
Soil Reservoir Dynamics of Ophidiomyces ophidiicola, the Causative Agent of Snake Fungal Disease


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Initial Survey
2.2. Sample Collection
2.3. Environmental Oo Detection
2.4. Soil Sterilization and Characterization
2.5. Microcosm Growth Assays
2.6. Microbial Community Analyses
3. Results
3.1. Initial Survey
3.2. Oo Is More Prevalent Within Snake Hibernacula
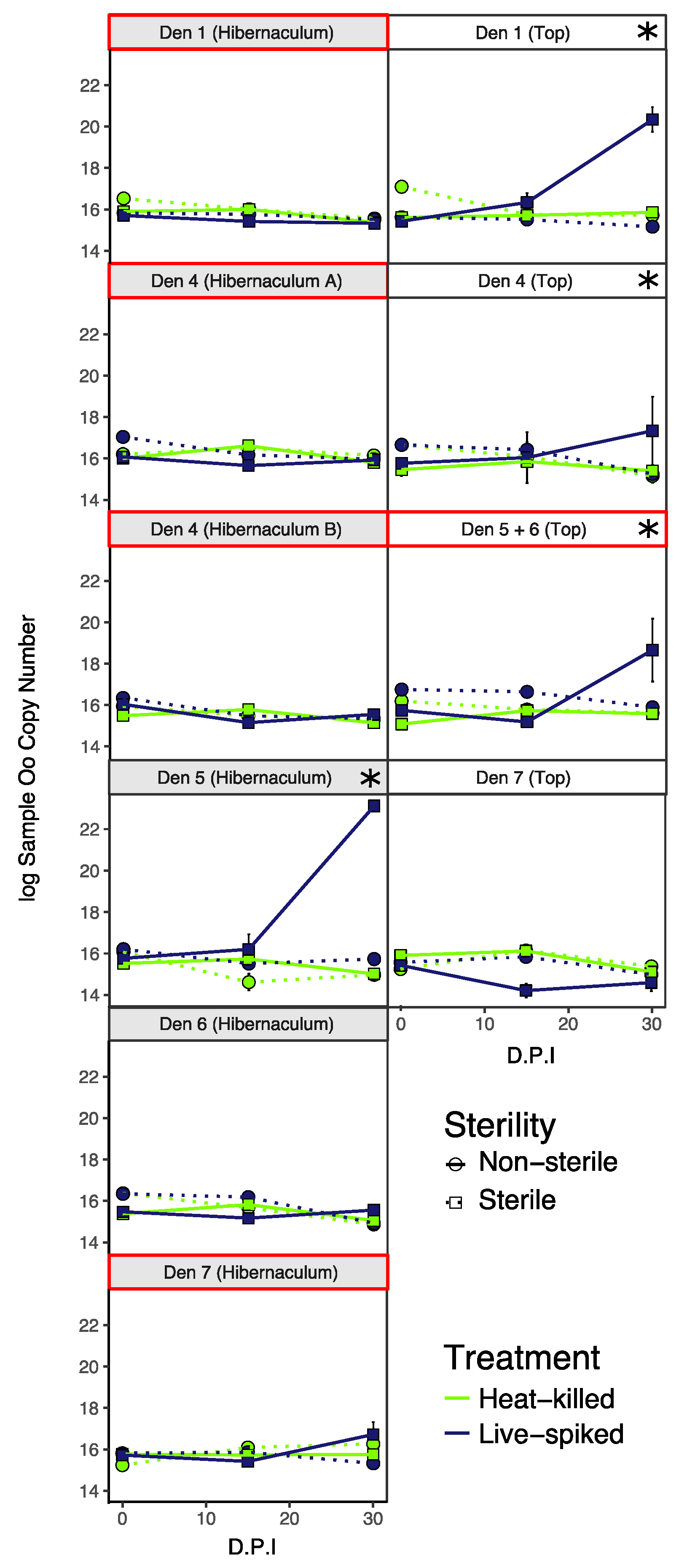
3.3. Oo Is Capable of Growth in Sterile Soils
3.4. No Evidence That Detection or Growth of Oo Is Linked to Abiotic Soil Parameters
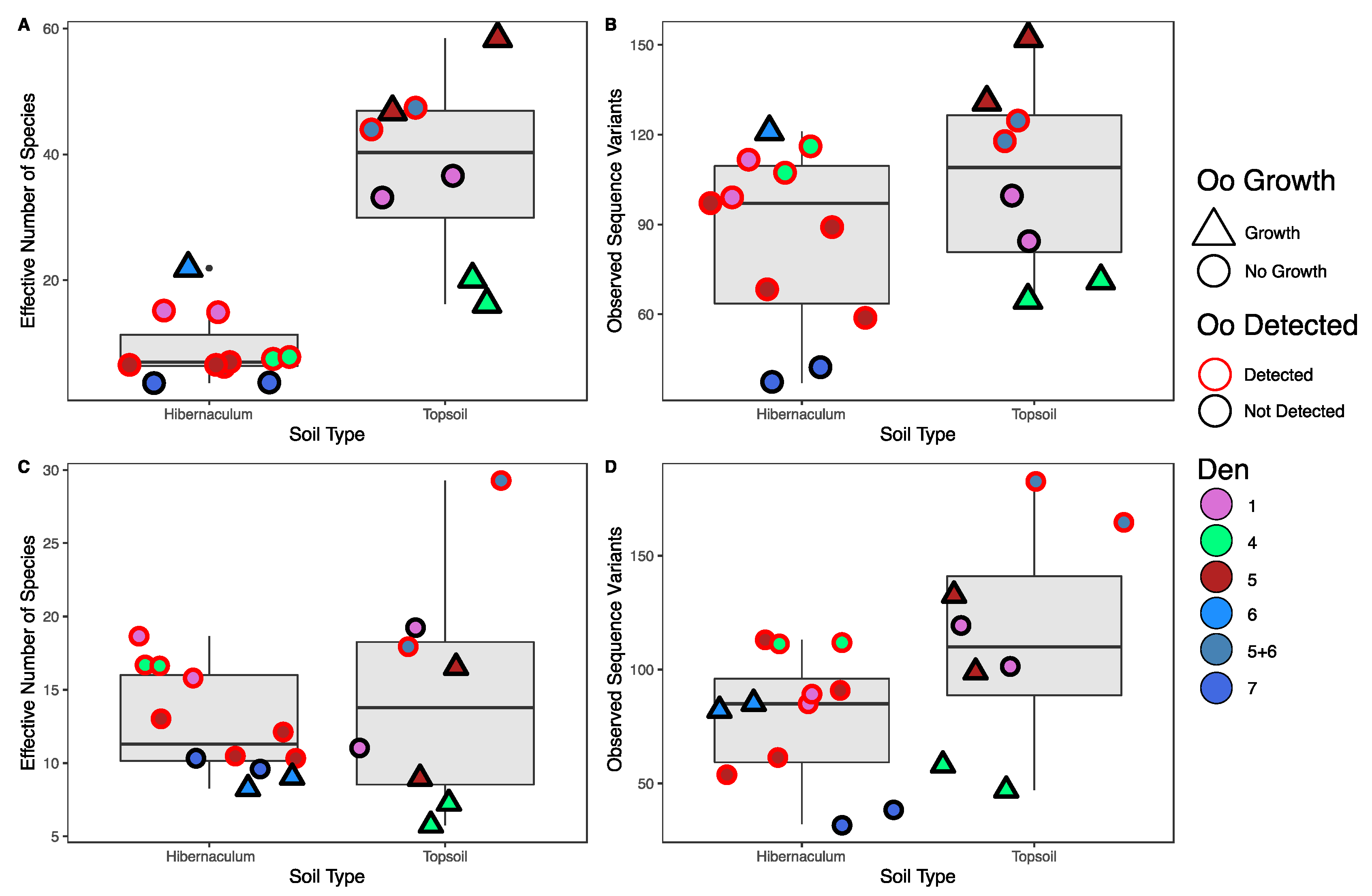
3.5. Detection and Growth of Oo Are Correlated with Microbial Community Diversity but Not Richness
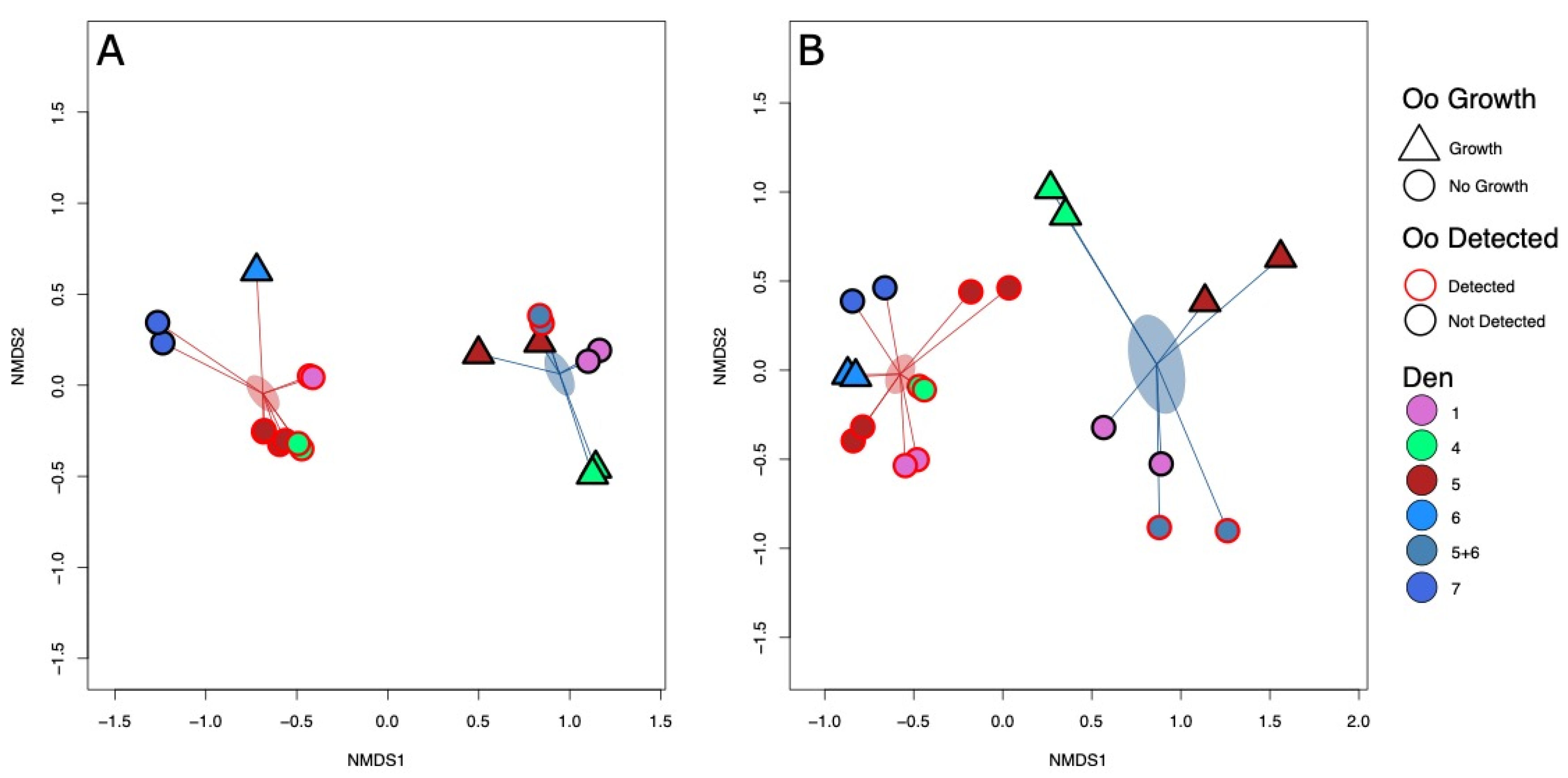
3.6. Detection and Growth of Oo Are Correlated with Microbial Community Composition
4. Discussion
5. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Jones, K.E.; Patel, N.G.; Levy, M.A. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J.R.; Barrett, C.B.; Civitello, D.J.; Craft, M.E.; Delius, B.; DeLeo, G.A.; Hudson, P.J.; Jouanard, N.; Nguyen, K.H.; Ostfeld, R.S.; et al. Emerging human infectious diseases and the links to global food production. Nat. Sustain. 2019, 2, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Mathiason, C.K.; Nalls, A.V.; Seelig, D.M.; Kraft, S.L.; Carnes, K.; Anderson, K.R.; Hayes-Klug, J.; Hoover, E.A. Susceptibility of Domestic Cats to Chronic Wasting Disease. J. Virol. 2013, 87, 1947–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, A.B.; Jones, K.E.; Nunn, C.L.; Altizer, S. Infectious diseases and extinction risk in wild mammals. Conserv. Biol. 2007, 21, 1269–1279. [Google Scholar] [CrossRef]
- van Agtmaal, M.; Straathof, A.; Termorshuizen, A.; Teurlincx, S.; Hundscheid, M.; Ruyters, S.; Busschaert, P.; Lievens, B.; Boer, W. de Exploring the reservoir of potential fungal plant pathogens in agricultural soil. Appl. Soil Ecol. 2017, 121, 152–160. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Coevolution of hosts and parasites. Parasitology 1982, 85, 411–426. [Google Scholar] [CrossRef]
- De Castro, F.; Bolker, B. Mechanisms of disease-induced extinction. Ecol. Lett. 2005, 8, 117–126. [Google Scholar] [CrossRef]
- Leach, C.B.; Webb, C.T.; Cross, P.C. When environmentally persistent pathogens transform good habitat into ecological traps. R. Soc. Open Sci. 2016, 3, 160051. [Google Scholar] [CrossRef] [Green Version]
- Lipsitch, M.; Stiller, S.; Nowak, M.A. The Evolution of Virulence in Pathogens With Vertical and Horizontal Transmission. Evolution 1996, 50, 1729–1741. [Google Scholar] [CrossRef]
- Burger, R.; Lynch, M. Evolution and Extinction in a Changing Environment: A Quantitative-Genetic Analysis. Evolution 1995, 49, 151–163. [Google Scholar] [CrossRef]
- Breban, R. Role of environmental persistence in pathogen transmission: A mathematical modeling approach. J. Math. Biol. 2013, 66, 535–546. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Z.; Dai, S.; Yan, J.; Wise, M.J. The Sit-and-Wait hypothesis in bacterial pathogens: A theoretical study of durability and virulence. Front. Microbiol. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewald, P.W. Transmission Modes and Evolution of the Parasitism-Mutualism Continuum. Ann. N. Y. Acad. Sci. 1987, 503, 295–306. [Google Scholar] [CrossRef]
- Walther, B.A.; Ewald, P.W. Pathogen survival in the external environment and the evolution of virulence. Biol. Rev. Camb. Philos. Soc. 2004, 79, 849–869. [Google Scholar] [CrossRef] [PubMed]
- Lorch, J.M.; Knowles, S.; Lankton, J.S.; Michell, K.; Edwards, J.L.; Kapfer, J.M.; Staffen, R.A.; Wild, E.R.; Schmidt, K.Z.; Ballmann, A.E.; et al. Snake fungal disease: An emerging threat to wild snakes. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allender, M.C.; Dreslik, M.; Wylie, S.; Phillips, C.; Wylie, D.B.; Maddox, C.; Delaney, M.A.; Kinsel, M.J. Chrysosporium sp. Infection in Eastern Massasauga Rattlesnakes. Emerg. Infect. Dis. 2011, 17, 2383–2384. [Google Scholar] [CrossRef] [PubMed]
- Burbrink, F.T.; Lorch, J.M.; Lips, K.R. Host susceptibility to snake fungal disease is highly dispersed across phylogenetic and functional trait space. Sci. Adv. 2017, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, W.J.; Aveling, R.; Brooks, T.M.; Clout, M.; Dicks, L.V.; Fellman, L.; Fleishman, E.; Gibbons, D.W.; Keim, B.; Lickorish, F.; et al. A horizon scan of global conservation issues for 2014. Trends Ecol. Evol. 2014, 29, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Allender, M.C.; Raudabaugh, D.B.; Gleason, F.H.; Miller, A.N. The natural history, ecology, and epidemiology of Ophidiomyces ophiodiicola and its potential impact on free-ranging snake populations. Fungal Ecol. 2015, 17, 187–196. [Google Scholar] [CrossRef]
- Lorch, J.M.; Lankton, J.; Werner, K.; Falendysz, E.A.; McCurley, K.; Blehert, D.S. Experimental infection of snakes with ophidiomyces ophiodiicola causes pathological changes that typify snake fungal disease. MBio 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, D.M.; Leys, J.E.; Grisnik, M.; Grajal-Puche, A.; Murray, C.M.; Allender, M.C. Variability in snake skin microbial assemblages across spatial scales and disease states. ISME J. 2019, 13, 2209–2222. [Google Scholar] [CrossRef] [Green Version]
- Nash Smith, S. Comparison of Germination of Pathogenic Fusarium oxysporum Chlamydospores in Host Rhizosphere Soils Conducive and Suppressive to Wilts. Phytopathology 1977, 77, 502. [Google Scholar] [CrossRef]
- Ajello, L. Soil as natural reservoir for human pathogenic fungi. Science 1956, 123, 876–879. [Google Scholar] [CrossRef] [PubMed]
- Lennon, J.T.; Jones, S.E. Microbial seed banks: The ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 2011, 9, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Kühn, J.; Rippel, R.; Schmidhalter, U. Abiotic soil properties and the occurrence of rhizoctonia crown and root rot in sugar beet. J. Plant Nutr. Soil Sci. 2009, 172, 661–668. [Google Scholar] [CrossRef]
- Peng, H.X.; Sivasithamparam, K.; Turner, D.W. Chlamydospore germination and Fusarium wilt of banana plantlets in suppressive and conducive soils are affected by physical and chemical factors. Soil Biol. Biochem. 1999, 31, 1363–1374. [Google Scholar] [CrossRef]
- Schlatter, D.; Kinkel, L.; Thomashow, L.; Weller, D.; Paulitz, T. Disease suppressive soils: New insights from the soil microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar] [CrossRef] [Green Version]
- Burger, J.; Zappalorti, R.T.; Gochfeld, M.; Boarman, W.I.; Caffrey, M.; Doig, V.; Garber, S.D.; Lauro, B.; Mikovsky, M.; Caffrey, M.; et al. Hibernacula and Summer Den Sites of Pine Snakes ( Pituophis melanoleucus ) in the New Jersey Pine Barrens. J. Herpetol. 1988, 22, 425–433. [Google Scholar] [CrossRef]
- Burger, J.; Zappalorti, R.T.; Gochfeld, M.; Devito, E.; Schneider, D.; McCort, M.; Jeitner, C. Long-term use of hibernacula by Northern Pinesnakes (Pituophis melanoleucus). J. Herpetol. 2012, 46, 596–601. [Google Scholar] [CrossRef]
- Burger, J.; Zappalorti, R. Hibernation Site Philopatry in Northern Pine Snakes (Pituophis melanoleucus) in New Jersey. J. Herpetol. 2015, 49, 245–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burger, J.; Zappalorti, R. Conservation and Protection of Threatened Pine Snakes in the New Jersey Pine Barrens. Herpetol. Conserv. Biol. 2016, 11, 304–314. [Google Scholar]
- Bohuski, E.; Lorch, J.M.; Griffin, K.M.; Blehert, D.S. TaqMan real-time polymerase chain reaction for detection of Ophidiomyces ophiodiicola, the fungus associated with snake fungal disease. BMC Vet. Res. 2015, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorch, J.M.; Muller, L.K.; Russell, R.E.; O’Connor, M.; Lindner, D.L.; Blehert, D.S. Distribution and environmental persistence of the causative agent of white-nose syndrome, geomyces destructans, in bat hibernacula of the eastern United States. Appl. Environ. Microbiol. 2013, 79, 1293–1301. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.0.1; R Foundation for Statistical Computing: Wien, Austria, 2020; Available online: https://cran.r-project.org/ (accessed on 31 October 2020).
- Lorch, J.M.; Meteyer, C.U.; Behr, M.J.; Boyles, J.G.; Cryan, P.M.; Hicks, A.C.; Ballmann, A.E.; Coleman, J.T.H.; Redell, D.N.; Reeder, D.M.; et al. Experimental infection of bats with Geomyces destructans causes white-nose syndrome. Nature 2011, 480, 376–378. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burger, J.; Zappalorti, R.T. The Northern Pine Snake (Pituophis melanoleucus): Its Life History, Behaviour and Conservation; Nova Science Publishers: Hauppauge, NY, USA, 2011; Volume 16, ISBN 9781612094526. [Google Scholar]
- Reynolds, H.T.; Raudabaugh, D.B.; Lilje, O.; Allender, M.C.; Miller, A.N.; Gleason, F.H. Emerging Mycoses and Fungus-Like Diseases of Vertebrate Wildlife. In The Fungal Community; Dighton, J., White, J.F., Eds.; CRC: Boca Raton, FL, USA, 2017; pp. 385–404. [Google Scholar]
- Weller, D.M.; Raaijmakers, J.M.; McSpadden Gardener, B.B.; Thomashow, L.S. Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.J.; Rovira, A.D. The role of bacteria in the biological control of Gaeumannomyces graminis by suppressive soils. Soil Biol. Biochem. 1976, 8, 269–273. [Google Scholar] [CrossRef]
- Fleck, C.B.; Schöbel, F.; Brock, M. Nutrient acquisition by pathogenic fungi: Nutrient availability, pathway regulation, and differences in substrate utilization. Int. J. Med. Microbiol. 2011, 301, 400–407. [Google Scholar] [CrossRef]
- Powlson, D.S.; Jenkinson, D.S. The effects of biocidal treatments on metabolism in soil-II. Gamma irradiation, autoclaving, air-drying and fumigation. Soil Biol. Biochem. 1976, 8, 179–188. [Google Scholar] [CrossRef]
- Hornby, D. Suppressive Soils. Annu. Rev. Phytopathol. 1983, 21, 65–85. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Burpee, L.L. The influence of abiotic factors on biological control of soilborne plant pathogenic fungi. Can. J. Plant Pathol. 1990, 12, 308–317. [Google Scholar] [CrossRef]
- Al-Shorbaji, F.N.; Gozlan, R.E.; Roche, B.; Robert Britton, J.; Andreou, D. The alternate role of direct and environmental transmission in fungal infectious disease in wildlife: ThreatsS for biodiversity conservation. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, H.T.; Ingersoll, T.; Barton, H.A. Modeling the environmental growth of Pseudogymnoascus destructans and its impact on the White-nose syndrom epidemic. J. Wildl. Dis. 2015, 51, 318–331. [Google Scholar] [CrossRef]
- Frick, W.; Cheng, T.; Langwig, K.; Hoyt, J.; Janicki, A.; Parise, K.; Foster, J.; Kilpatrick, A. Pathogen dynamics during invasion and establishment of white-nose syndrome explain mechanisms of host persistence. Ecology 2017, 98, 624–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigler, L.; Hambleton, S.; Paré, J.A. Molecular characterization of reptile pathogens currently known as members of the chrysosporium anamorph of nannizziopsis vriesii complex and relationship with some human-associated isolates. J. Clin. Microbiol. 2013, 51, 3338–3357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraut-Cohen, J.; Zolti, A.; Shaltiel-Harpaz, L.; Argaman, E.; Rabinovich, R.; Green, S.J.; Minz, D. Effects of tillage practices on soil microbiome and agricultural parameters. Sci. Total Environ. 2020, 705, 135791. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| A—Initial Survey | |||
| Soil name | Type | Oo qPCR CT Value | qPCR Interpretation |
| Den 1 A | Hibernaculum | 35.43 | Detected |
| Den 1 B | Hibernaculum | 37.42 | Detected |
| Den 1 C | Hibernaculum | 31.74 | Detected |
| Den 1 D | Hibernaculum | 31.96 | Detected |
| Den 1 E | Hibernaculum | 32.01 | Detected |
| Den 1 F | Hibernaculum | 35.78 | Detected |
| Den 1 G | Hibernaculum | 31.6 | Detected |
| Den 1 H | Hibernaculum | NA | Not Detected |
| Den 2 A | Hibernaculum | 35.68 | Detected |
| Den 2 B | Hibernaculum | 40 | Detected |
| Den 3 A | Hibernaculum | 34.87 | Detected |
| Den 3 B | Hibernaculum | 37.25 | Detected |
| Den 3 C | Hibernaculum | NA | Not Detected |
| Den 3 D | Hibernaculum | 35.55 | Detected |
| Den 3 E | Hibernaculum | 34.23 | Detected |
| B—Follow-Up Study | Mean CT | ||
| Den 1 | Hibernaculum | 34.86 | Detected |
| Den 4 A | Hibernaculum | 27.14 | Detected |
| Den 4 B | Hibernaculum | 32.66 | Detected |
| Den 5 | Hibernaculum | NA | Not Detected |
| Den 6 | Hibernaculum | NA | Not Detected |
| Den 7 | Hibernaculum | 35.549 | Detected |
| Den 1 | Topsoil | NA | Not Detected |
| Den 4 | Topsoil | NA | Not Detected |
| Den 5+6 | Topsoil | 39.19 | Detected |
| Den 7 | Topsoil | NA | Not Detected |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, L.J.; Burger, J.; Zappalorti, R.T.; Bunnell, J.F.; Winzeler, M.E.; Taylor, D.R.; Lorch, J.M. Soil Reservoir Dynamics of Ophidiomyces ophidiicola, the Causative Agent of Snake Fungal Disease. J. Fungi 2021, 7, 461. https://doi.org/10.3390/jof7060461
Campbell LJ, Burger J, Zappalorti RT, Bunnell JF, Winzeler ME, Taylor DR, Lorch JM. Soil Reservoir Dynamics of Ophidiomyces ophidiicola, the Causative Agent of Snake Fungal Disease. Journal of Fungi. 2021; 7(6):461. https://doi.org/10.3390/jof7060461
Chicago/Turabian StyleCampbell, Lewis J., Joanna Burger, Robert T. Zappalorti, John F. Bunnell, Megan E. Winzeler, Daniel R. Taylor, and Jeffrey M. Lorch. 2021. "Soil Reservoir Dynamics of Ophidiomyces ophidiicola, the Causative Agent of Snake Fungal Disease" Journal of Fungi 7, no. 6: 461. https://doi.org/10.3390/jof7060461