
Potential of Antifungal Proteins (AFPs) to Control Penicillium Postharvest Fruit Decay

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Growth Conditions
2.2. AFP Production and Purification
2.3. Antifungal Assays
2.4. Protection Assays against Fungal Infections Caused by Penicillium spp. in Fruits
3. Results
3.1. Antifungal Activity Assays
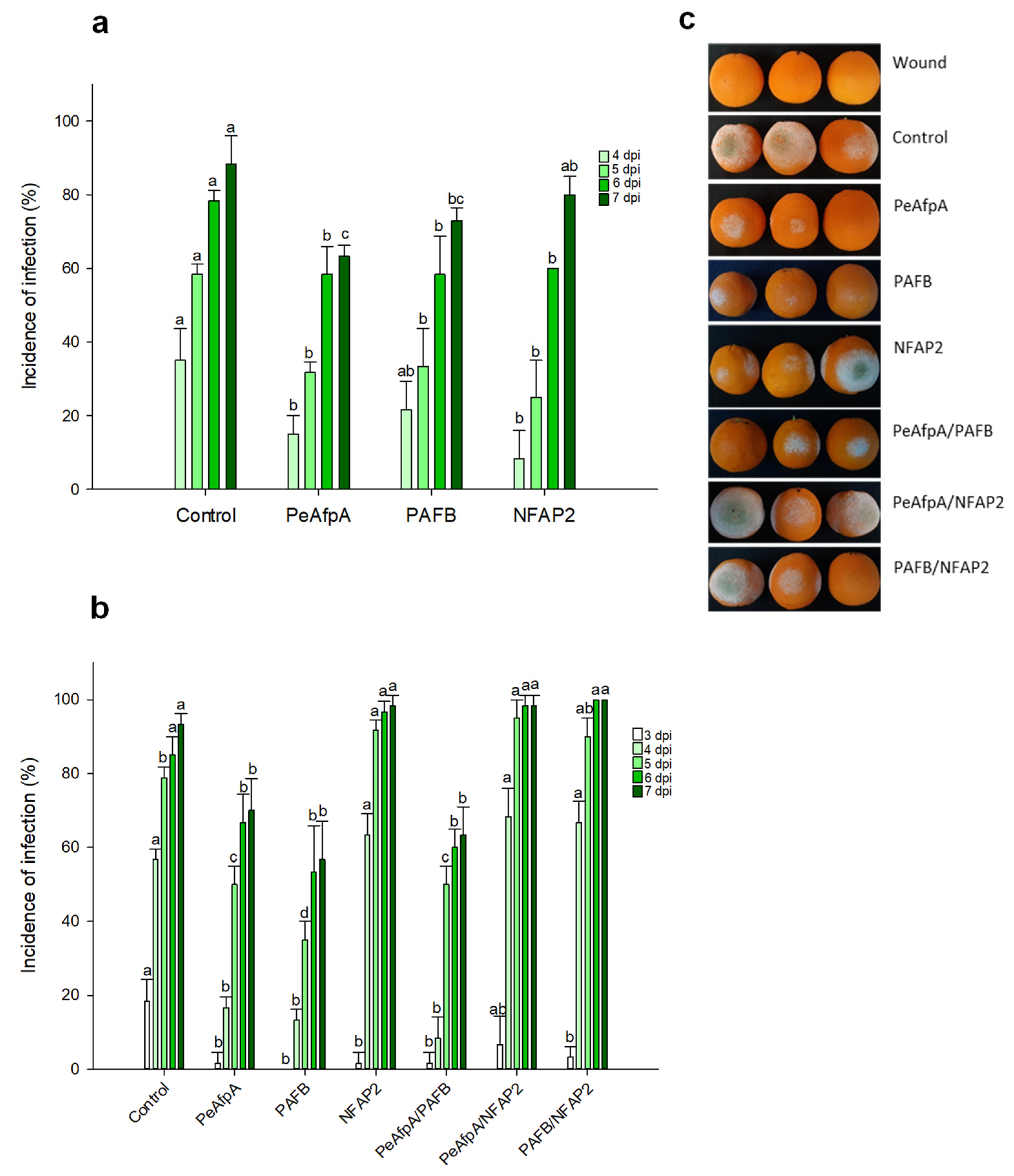
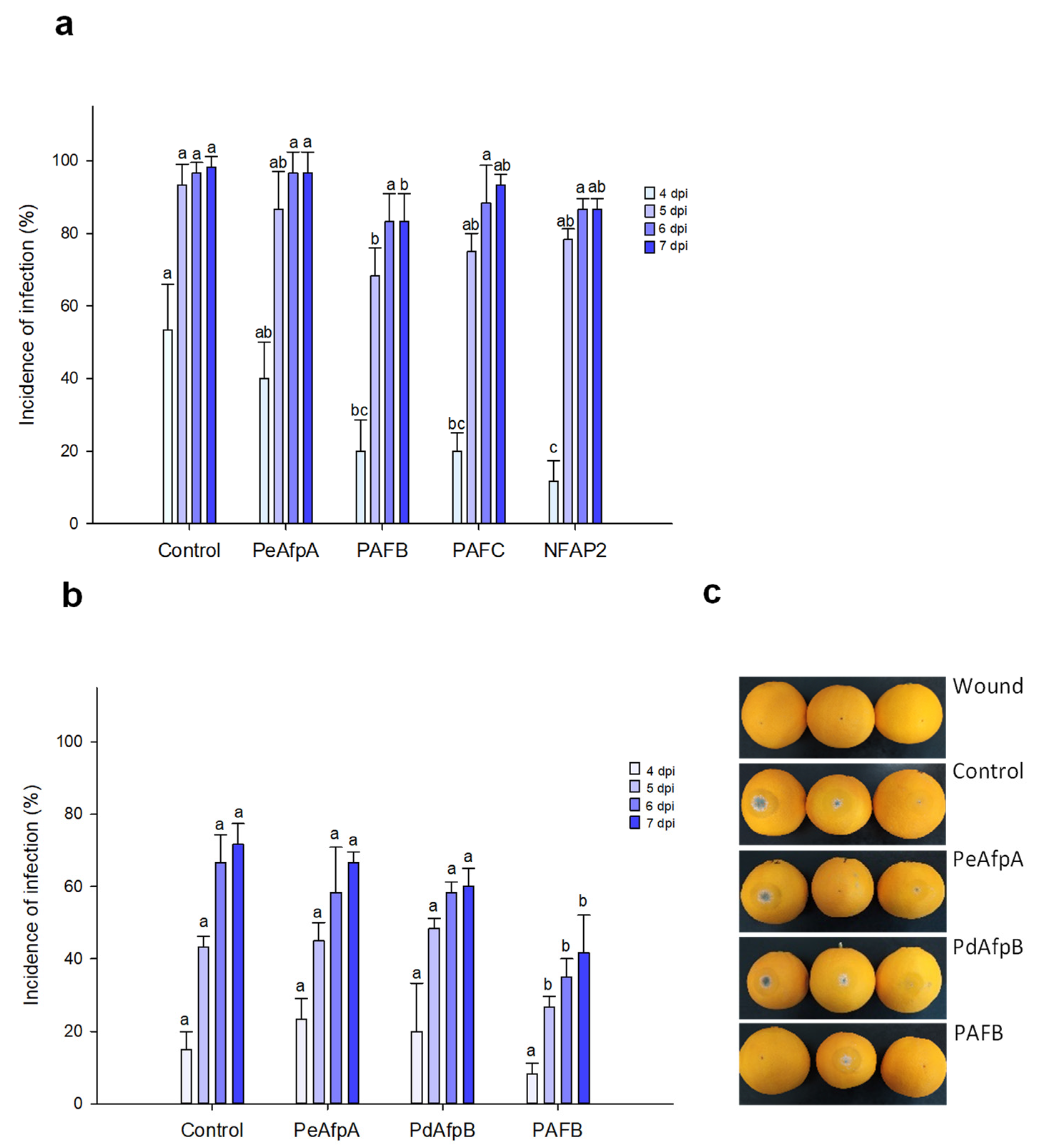
3.2. PAFB Delays P. digitatum Infection in Orange Fruits
3.3. PAFB Delays P. italicum Infection in Orange Fruits
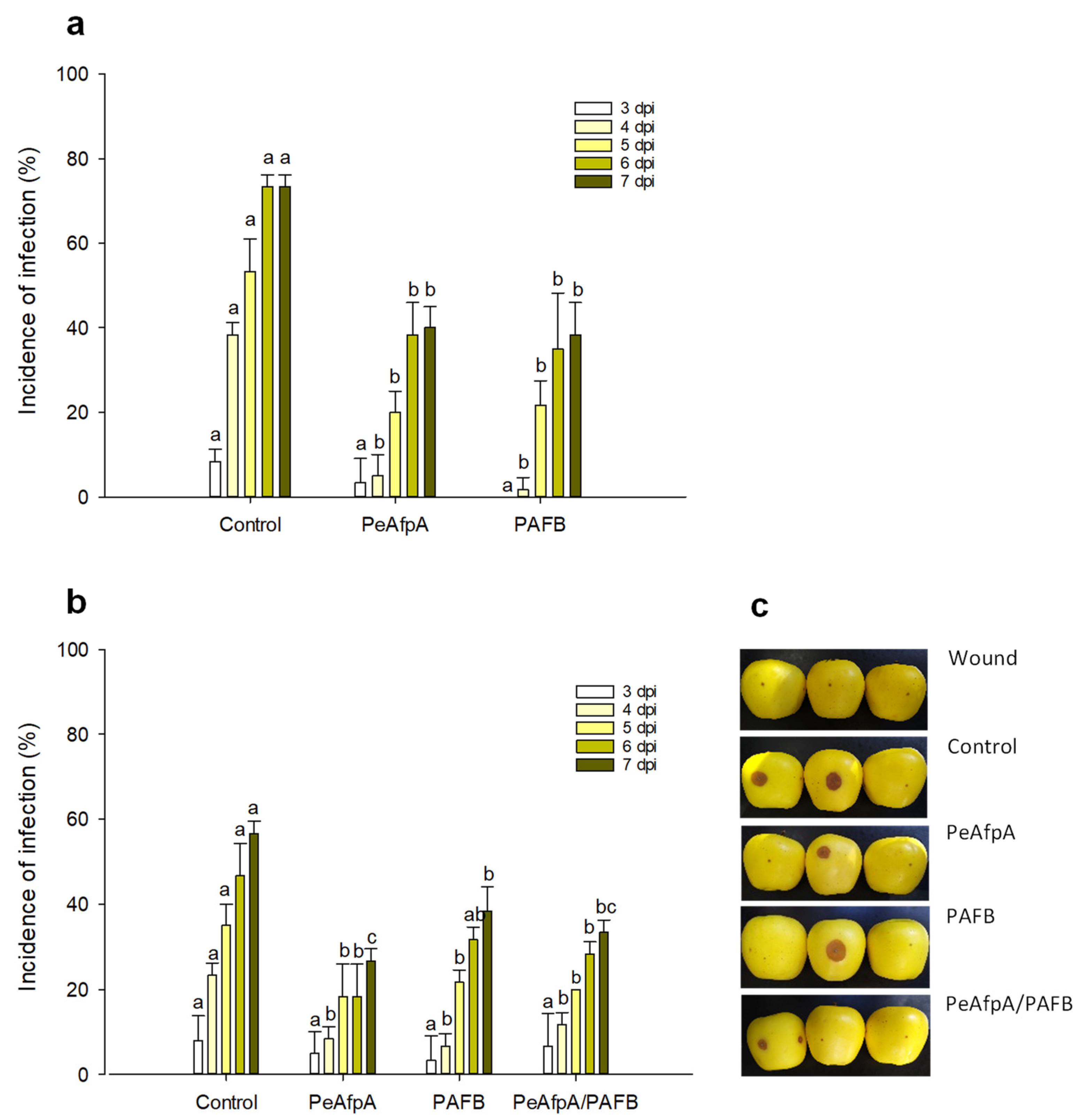
3.4. PAFB Delays P. expansum Infection in Apple Fruits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palou, L.; Ali, A.; Fallik, E.; Romanazzi, G. GRAS, plant- and animal-derived compounds as alternatives to conventional fungicides for the control of postharvest diseases of fresh horticultural produce. Postharvest Biol. Technol. 2016, 122, 41–52. [Google Scholar] [CrossRef]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Palou, L. Penicillium digitatum, Penicillium italicum (Green Mold, Blue Mold). In Postharvest Decay: Control Strategies; Bautista-Baños, S., Ed.; Elsevier: London, UK, 2014; pp. 45–102. [Google Scholar]
- Plaza, P.; Usall, J.; Torres, R.; Lamarca, N.; Asensio, A.; Viñas, I. Control of green and blue mould by curing on oranges during ambient and cold storage. Postharvest Biol. Technol. 2003, 28, 195–198. [Google Scholar] [CrossRef]
- Errampalli, D. Penicillium expansum (blue mold). In Postharvest Decay. Control Strategies; Bautista-Banos, S., Ed.; Academic Press: Cambridge, MA, USA; Elsevier Inc.: London, UK, 2014; pp. 189–231. [Google Scholar]
- Turechek, W.W. Apple diseases and their management. In Diseases of Fruits and Vegetables Volume I: Diagnosis and Management; Naqvi, S.A.M.H., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2004; Volume 1, pp. 1–108. [Google Scholar]
- Tannous, J.; Keller, N.P.; Atoui, A.; El Khoury, A.; Lteif, R.; Oswald, I.P.; Puel, O. Secondary metabolism in Penicillium expansum: Emphasis on recent advances in patulin research. Crit. Rev. Food Sci. Nutr. 2018, 58, 2082–2098. [Google Scholar] [CrossRef]
- Galgóczy, L.; Marx, F. Do Antimicrobial Proteins contribute to overcoming the hidden antifungal crisis at the dawn of a post-antibiotic era? Microorganisms 2019, 7, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos, J.F.; Muñoz, A.; Pérez-Payá, E.; Misra, S.; López-García, B. Identification and rational design of novel antimicrobial peptides for plant protection. Annu. Rev. Phytopathol. 2008, 46, 273–301. [Google Scholar] [CrossRef] [PubMed]
- Delgado, J.; Owens, R.A.; Doyle, S.; Asensio, M.A.; Núñez, F. Antifungal proteins from moulds: Analytical tools and potential application to dry-ripened foods. Appl. Microbiol. Biotechnol. 2016, 100, 6991–7000. [Google Scholar] [CrossRef] [Green Version]
- Hegedüs, N.; Marx, F. Antifungal proteins: More than antimicrobials? Fungal Biol. Rev. 2013, 26, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Marx, F.; Binder, U.; Leiter, É.; Pócsi, I. The Penicillium chrysogenum antifungal protein PAF, a promising tool for the development of new antifungal therapies and fungal cell biology studies. Cell. Mol. Life Sci. 2008, 65, 445–454. [Google Scholar] [CrossRef]
- Garrigues, S.; Gandía, M.; Castillo, L.; Coca, M.; Marx, F.; Marcos, J.F.; Manzanares, P. Three Antifungal Proteins From Penicillium expansum: Different Patterns of Production and Antifungal Activity. Front. Microbiol. 2018, 9, 2370. [Google Scholar] [CrossRef] [Green Version]
- Garrigues, S.; Gandía, M.; Popa, C.; Borics, A.; Marx, F.; Coca, M.; Marcos, J.F.; Manzanares, P. Efficient production and characterization of the novel and highly active antifungal protein AfpB from Penicillium digitatum. Sci. Rep. 2017, 7, 14663. [Google Scholar] [CrossRef] [Green Version]
- Huber, A.; Hajdu, D.; Bratschun-Khan, D.; Gáspári, Z.; Varbanov, M.; Philippot, S.; Fizil, Á.; Czajlik, A.; Kele, Z.; Sonderegger, C.; et al. New antimicrobial potential and structural properties of PAFB: A cationic, cysteine-rich protein from Penicillium chrysogenum Q176. Sci. Rep. 2018, 8, 1751. [Google Scholar] [CrossRef]
- Kovács, R.; Holzknecht, J.; Hargitai, Z.; Papp, C.; Farkas, A.; Borics, A.; Tóth, L.; Váradi, G.; Tóth, G.K.; Kovács, I.; et al. In vivo applicability of Neosartorya fischeri antifungal protein 2 (NFAP2) in treatment of vulvovaginal candidiasis. Antimicrob. Agents Chemother. 2019, 63, e01777-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palicz, Z.; Jenes, Á.; Gáll, T.; Miszti-Blasius, K.; Kollár, S.; Kovács, I.; Emri, M.; Márián, T.; Leiter, É.; Pócsi, I.; et al. In vivo application of a small molecular weight antifungal protein of Penicillium chrysogenum (PAF). Toxicol. Appl. Pharmacol. 2013, 269, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Szappanos, H.; Szigeti, G.P.; Pal, B.; Rusznak, Z.; Szucs, G.; Rajnavolgyi, E.; Balla, J.; Balla, G.; Nagy, E.; Leiter, E.; et al. The antifungal protein AFP secreted by Aspergillus giganteus does not cause detrimental effects on certain mammalian cells. Peptides 2006, 27, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Szappanos, H.; Szigeti, G.P.; Pál, B.; Rusznák, Z.; Szűcs, G.; Rajnavölgyi, É.; Balla, J.; Balla, G.; Nagy, E.; Leiter, É.; et al. The Penicillium chrysogenum-derived antifungal peptide shows no toxic effects on mammalian cells in the intended therapeutic concentration. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2005, 371, 122–132. [Google Scholar] [CrossRef]
- Tóth, L.; Boros, E.; Poor, P.; Ordog, A.; Kele, Z.; Varadi, G.; Holzknecht, J.; Bratschun-Khan, D.; Nagy, I.; Toth, G.K.; et al. The potential use of the Penicillium chrysogenum antifungal protein PAF, the designed variant PAFopt and its γ-core peptide Pγopt in plant protection. Microb. Biotechnol. 2020, 13, 1403–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, L.; Váradi, G.; Boros, É.; Borics, A.; Ficze, H.; Nagy, I.; Tóth, G.K.; Rákhely, G.; Marx, F.; Galgóczy, L. Biofungicidal Potential of Neosartorya (Aspergillus) fischeri Antifungal Protein NFAP and Novel Synthetic γ-Core Peptides. Front. Microbiol. 2020, 11, 820. [Google Scholar] [CrossRef]
- Shi, X.; Cordero, T.; Garrigues, S.; Marcos, J.F.; Daròs, J.A.; Coca, M. Efficient production of antifungal proteins in plants using a new transient expression vector derived from tobacco mosaic virus. Plant Biotechnol. J. 2019, 17, 1069–1080. [Google Scholar] [CrossRef]
- Sonderegger, C.; Galgóczy, L.; Garrigues, S.; Fizil, Á.; Borics, A.; Manzanares, P.; Hegedüs, N.; Huber, A.; Marcos, J.F.; Batta, G.; et al. A Penicillium chrysogenum-based expression system for the production of small, cysteine-rich antifungal proteins for structural and functional analyses. Microb. Cell Fact. 2016, 15, 192. [Google Scholar] [CrossRef] [Green Version]
- Garrigues, S.; Gandía, M.; Marcos, J. Occurrence and function of fungal antifungal proteins: A case study of the citrus postharvest pathogen Penicillium digitatum. Appl. Microbiol. Biotechnol. 2016, 100, 2243–2256. [Google Scholar] [CrossRef]
- Huber, A.; Galgóczy, L.; Váradi, G.; Holzknecht, J.; Kakar, A.; Malanovic, N.; Leber, R.; Koch, J.; Keller, M.A.; Batta, G.; et al. Two small, cysteine-rich and cationic antifungal proteins from Penicillium chrysogenum: A comparative study of PAF and PAFB. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183246. [Google Scholar] [CrossRef]
- Sonderegger, C.; Váradi, G.; Galgóczy, L.; Kocsubé, S.; Posch, W.; Borics, A.; Dubrac, S.; Tóth, G.K.; Wilflingseder, D.; Marx, F. The Evolutionary Conserved γ-Core Motif Influences the Anti-Candida Activity of the Penicillium chrysogenum Antifungal Protein PAF. Front. Microbiol. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed]
- Tóth, L.; Kele, Z.; Borics, A.; Nagy, L.G.; Váradi, G.; Virágh, M.; Takó, M.; Vágvölgyi, C.; Galgóczy, L. NFAP2, a novel cysteine-rich anti-yeast protein from Neosartorya fischeri NRRL 181: Isolation and characterization. AMB Express 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, F.; Haas, H.; Reindl, M.; Stoffler, G.; Lottspeich, F.; Redl, B. Cloning, structural organization and regulation of expression of the Penicillium chrysogenum paf gene encoding an abundantly secreted protein with antifungal activity. Gene 1995, 167, 167–171. [Google Scholar] [CrossRef]
- Huber, A.; Lerchster, H.; Marx, F. Nutrient Excess Triggers the Expression of the Penicillium chrysogenum Antifungal Protein PAFB. Microorganisms 2019, 7, 654. [Google Scholar] [CrossRef] [Green Version]
- Holzknecht, J.; Kühbacher, A.; Papp, C.; Farkas, A.; Váradi, G.; Marcos, J.F.; Manzanares, P.; Tóth, G.K.; Galgóczy, L.; Marx, F. The Penicillium chrysogenum Q176 Antimicrobial Protein PAFC Effectively Inhibits the Growth of the Opportunistic Human Pathogen Candida albicans. J. Fungi 2020, 6, 141. [Google Scholar] [CrossRef] [PubMed]
- Gandía, M.; Monge, A.; Garrigues, S.; Orozco, H.; Giner-Llorca, M.; Marcos, J.F.; Manzanares, P. Novel insights in the production, activity and protective effect of Penicillium expansum antifungal proteins. Int. J. Biol. Macromol. 2020, 164, 3922–3931. [Google Scholar] [CrossRef]
- Marcet-Houben, M.; Ballester, A.R.; de la Fuente, B.; Harries, E.; Marcos, J.F.; González-Candelas, L.; Gabaldón, T. Genome sequence of the necrotrophic fungus Penicillium digitatum, the main postharvest pathogen of citrus. BMC Genom. 2012, 13, 646. [Google Scholar] [CrossRef] [Green Version]
- Ballester, A.-R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; González-Candelas, L.; Gabaldón, T. Genome, transcriptome, and functional analyses of Penicillium expansum provide new insights into secondary metabolism and pathogenicity. Mol. Plant-Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, L.; Váradi, G.; Borics, A.; Batta, G.; Kele, Z.; Vendrinszky, Á.; Tóth, R.; Ficze, H.; Tóth, G.K.; Vágvölgyi, C.; et al. Anti-Candidal Activity and Functional Mapping of Recombinant and Synthetic Neosartorya fischeri Antifungal Protein 2 (NFAP2). Front. Microbiol. 2018, 9, 393. [Google Scholar] [CrossRef] [Green Version]
- Hernanz-Koers, M.; Gandía, M.; Garrigues, S.; Manzanares, P.; Yenush, L.; Orzaez, D.; Marcos, J.F. FungalBraid: A GoldenBraid-based modular cloning platform for the assembly and exchange of DNA elements tailored to fungal synthetic biology. Fungal Genet. Biol. 2018, 116, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiserer, L.; Oberparleiter, C.; Weiler-Gorz, R.; Burgstaller, W.; Leiter, E.; Marx, F. Characterization of the Penicillium chrysogenum antifungal protein PAF. Arch. Microbiol. 2003, 180, 204–210. [Google Scholar] [CrossRef]
- Martínez-Culebras, P.V.; Gandía, M.; Boronat, A.; Marcos, J.F.; Manzanares, P. Differential susceptibility of mycotoxin-producing fungi to distinct antifungal proteins (AFPs). Food Microbiol. 2021, 97, 103760. [Google Scholar] [CrossRef]
- Vila, L.; Lacadena, V.; Fontanet, P.; del Pozo, A.M.; Segundo, B.S. A protein from the mold Aspergillus giganteus is a potent inhibitor of fungal plant pathogens. Mol. Plant-Microbe Interact. 2001, 14, 1327–1331. [Google Scholar] [CrossRef] [Green Version]
- Theis, T.; Marx, F.; Salvenmoser, W.; Stahl, U.; Meyer, V. New insights into the target site and mode of action of the antifungal protein of Aspergillus giganteus. Res. Microbiol. 2005, 156, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Barakat, H. Bio-Control of Alternaria alternata during Banana Storage by Purified AFP Using Isoelectric Focusing Technique. Food Nutr. Sci. 2014, 5, 1482–1495. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Martín, A.; Acosta, R.; Liddell, S.; Nunez, F.; Benito, M.J.; Asensio, M.A. Characterization of the novel antifungal protein PgAFP and the encoding gene of Penicillium chrysogenum. Peptides 2010, 31, 541–547. [Google Scholar] [CrossRef]
- Delgado, J.; Acosta, R.; Rodriguez-Martin, A.; Bermudez, E.; Nunez, F.; Asensio, M.A. Growth inhibition and stability of PgAFP from Penicillium chrysogenum against fungi common on dry-ripened meat products. Int. J. Food Microbiol. 2015, 205, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Delgado, J.; Ballester, A.-R.; Núñez, F.; González-Candelas, L. Evaluation of the activity of the antifungal PgAFP protein and its producer mould against Penicillium spp postharvest pathogens of citrus and pome fruits. Food Microbiol. 2019, 84, 103266. [Google Scholar] [CrossRef] [PubMed]
- Badosa, E.; Ferre, R.; Frances, J.; Bardaji, E.; Feliu, L.; Planas, M.; Montesinos, E. Sporicidal Activity of Synthetic Antifungal Undecapeptides and Control of Penicillium Rot of Apples. Appl. Environ. Microbiol. 2009, 75, 5563–5569. [Google Scholar] [CrossRef] [Green Version]
- López-García, B.; Veyrat, A.; Pérez-Payá, E.; González-Candelas, L.; Marcos, J.F. Comparison of the activity of antifungal hexapeptides and the fungicides thiabendazole and imazalil against postharvest fungal pathogens. Int. J. Food Microbiol. 2003, 89, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Bugeda, A.; Garrigues, S.; Gandía, M.; Manzanares, P.; Marcos, J.F.; Coca, M. The antifungal protein AfpB induces regulated cell death in its parental fungus Penicillium digitatum. mSphere 2020, 5, e00595-20. [Google Scholar] [CrossRef] [PubMed]
- Leiter, E.; Szappanos, H.; Oberparleiter, C.; Kaiserer, L.; Csernoch, L.; Pusztahelyi, T.; Emri, T.; Posci, I.; Salvenmoser, W.; Marx, F. Antifungal protein PAF severely affects the integrity of the plasma membrane of Aspergillus nidulans and induces an apoptosis-like phenotype. Antimicrob. Agents Chemother. 2005, 49, 2445–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Amino Acids (aa) | Molecular Weight (Da) a | Number of Cysteines | pI b | GRAVY c | Charge at pH 7 d |
|---|---|---|---|---|---|---|
| PAF | 55 | 6242 | 6 | 8.93 | −1.374 | +4.7 |
| PeAfpA | 57 | 6618 | 6 | 9.48 | −1.081 | +8.7 |
| PdAfpB | 58 | 6570 | 6 | 9.06 | −1.000 | +5.9 |
| PAFB | 58 | 6500 | 6 | 8.83 | −1.031 | +5.2 |
| PAFC | 64 | 6630 | 8 | 7.71 | −0.767 | +0.9 |
| NFAP2 | 52 | 5600 | 6 | 9.01 | −0.731 | +5.2 |
| P. digitatum | P. italicum | P. expansum | Reference | |
|---|---|---|---|---|
| PAF | 16 (100) | 32 (200) | 16 (100) | this work |
| PAFB | 0.16 (1.0) | 0.25 (1.63) | 0.12 (0.78) | this work |
| PAFC | 4 (26.5) | 8 (53.0) | 32 (212) | this work |
| NFAP2 | 2 (11.1) | 1 (5.5) | >64 (>356) | this work |
| PeAfpA | 0.15 (1.0) | 0.3 (2.0) | 0.3 (2.0) | [13] |
| PdAfpB | 0.6 (4) | 0.3 (2.0) | 0.6 (4) | [14] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandía, M.; Kakar, A.; Giner-Llorca, M.; Holzknecht, J.; Martínez-Culebras, P.; Galgóczy, L.; Marx, F.; Marcos, J.F.; Manzanares, P. Potential of Antifungal Proteins (AFPs) to Control Penicillium Postharvest Fruit Decay. J. Fungi 2021, 7, 449. https://doi.org/10.3390/jof7060449
Gandía M, Kakar A, Giner-Llorca M, Holzknecht J, Martínez-Culebras P, Galgóczy L, Marx F, Marcos JF, Manzanares P. Potential of Antifungal Proteins (AFPs) to Control Penicillium Postharvest Fruit Decay. Journal of Fungi. 2021; 7(6):449. https://doi.org/10.3390/jof7060449
Chicago/Turabian StyleGandía, Mónica, Anant Kakar, Moisés Giner-Llorca, Jeanett Holzknecht, Pedro Martínez-Culebras, László Galgóczy, Florentine Marx, Jose F. Marcos, and Paloma Manzanares. 2021. "Potential of Antifungal Proteins (AFPs) to Control Penicillium Postharvest Fruit Decay" Journal of Fungi 7, no. 6: 449. https://doi.org/10.3390/jof7060449