
Elevated Sporulation Efficiency in Fission Yeast Schizosaccharomyces japonicus Strains Isolated from Drosophila


Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains, Media, and Culture Conditions
2.2. Isolation of S. japonicus from Drosophila
2.3. Sequence Analysis of S. japonicus Wild Strains
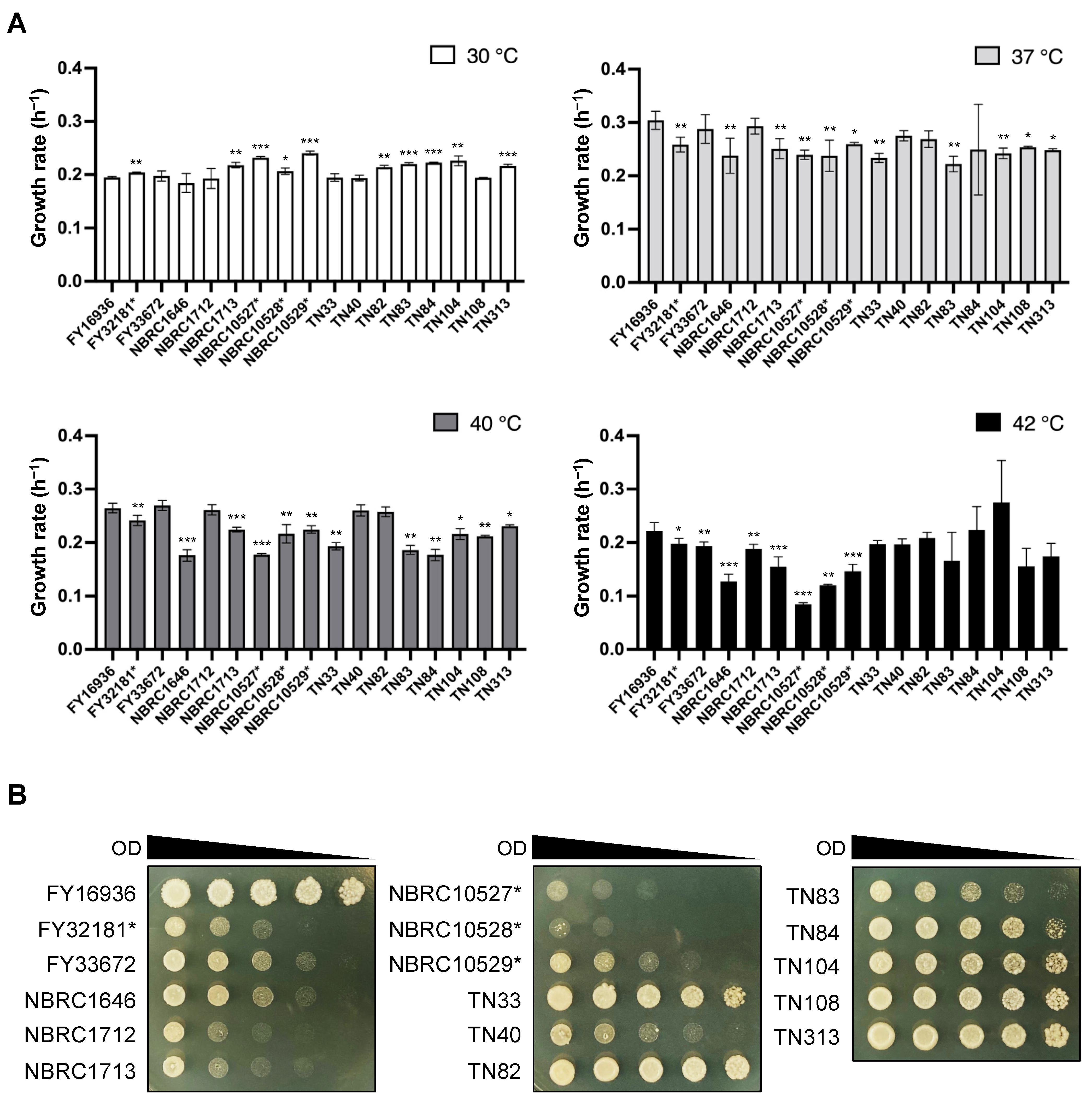
2.4. Growth Assay at Various Temperatures
2.5. Spotting Assay
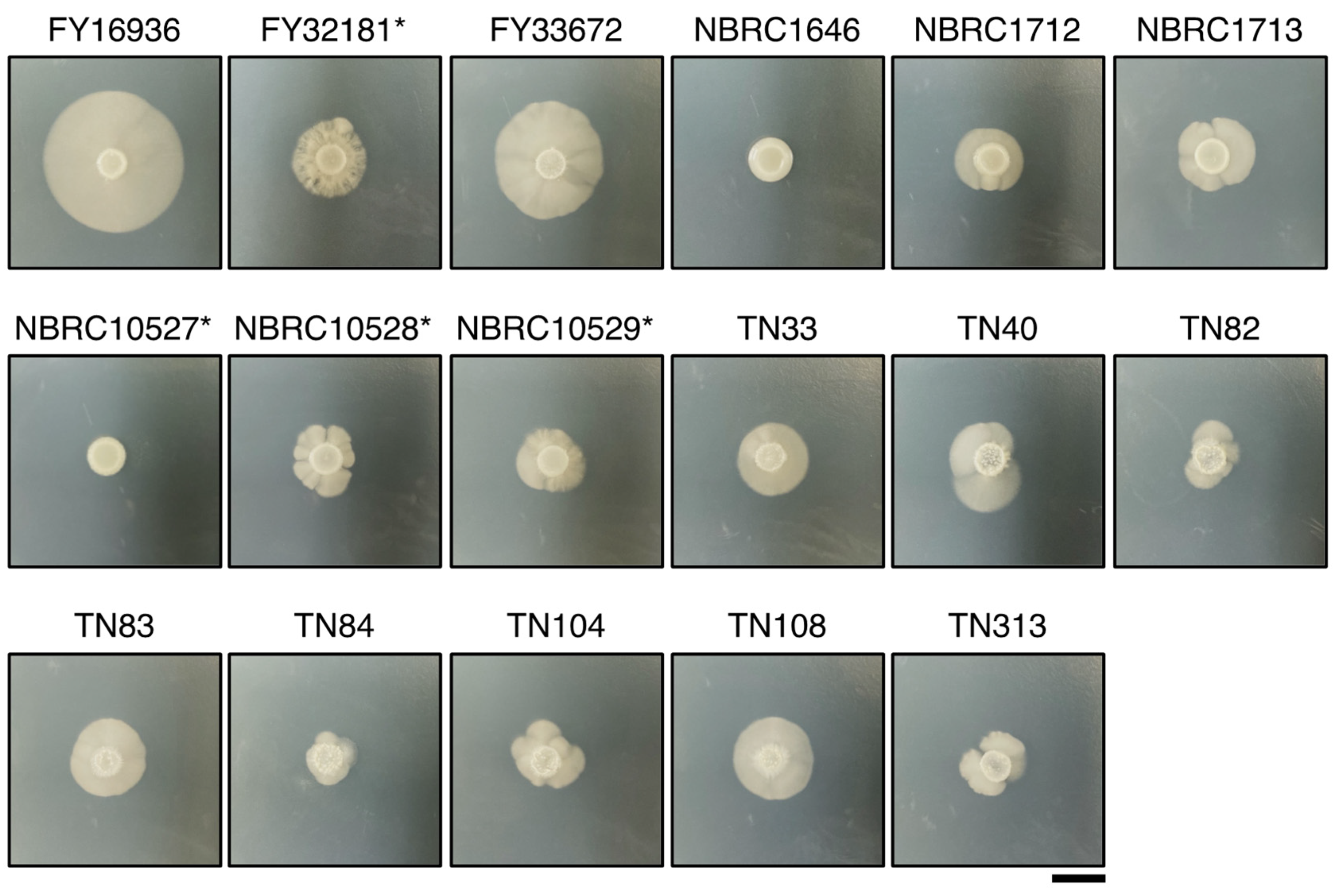
2.6. Hyphal Formation
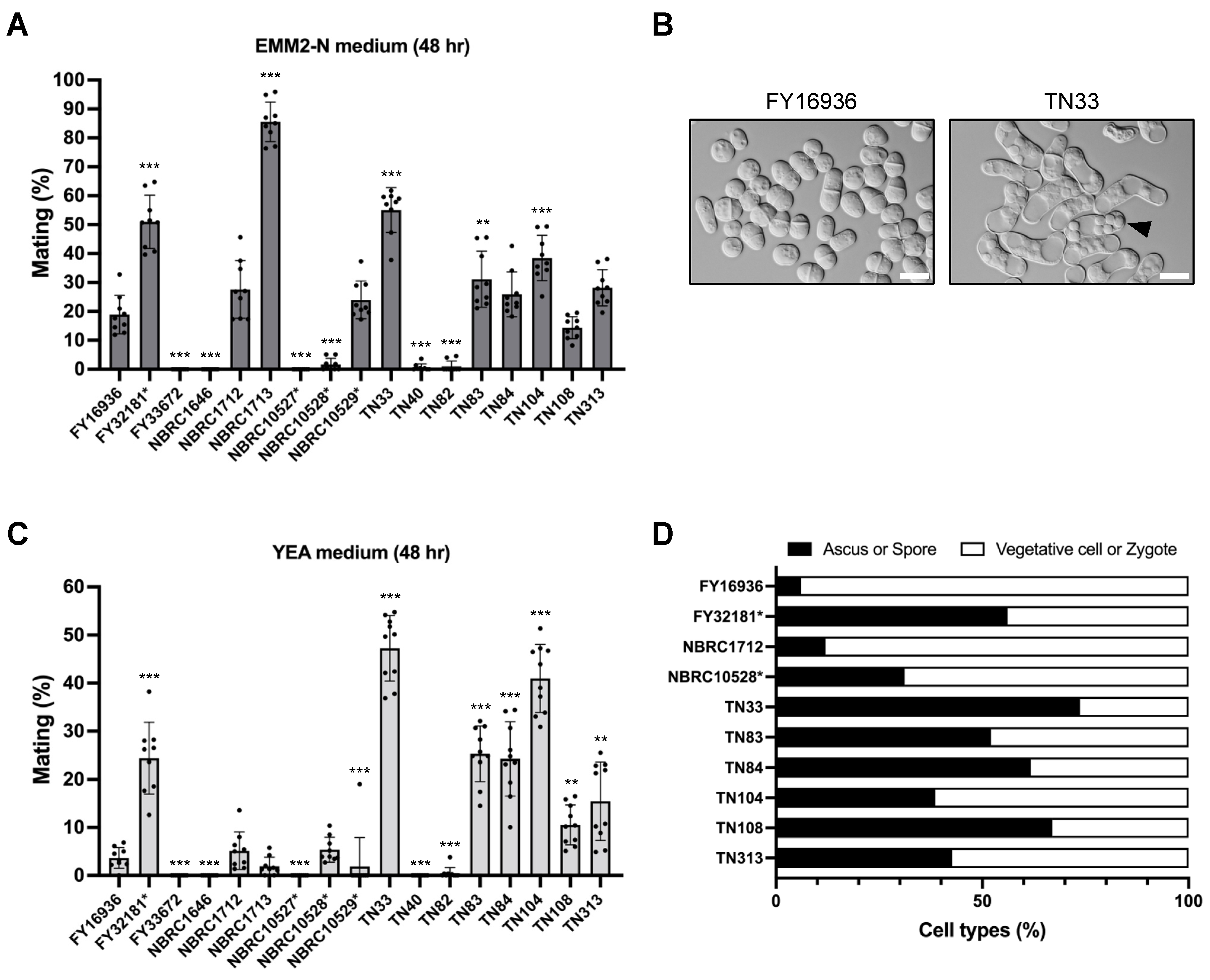
2.7. Mating Frequency
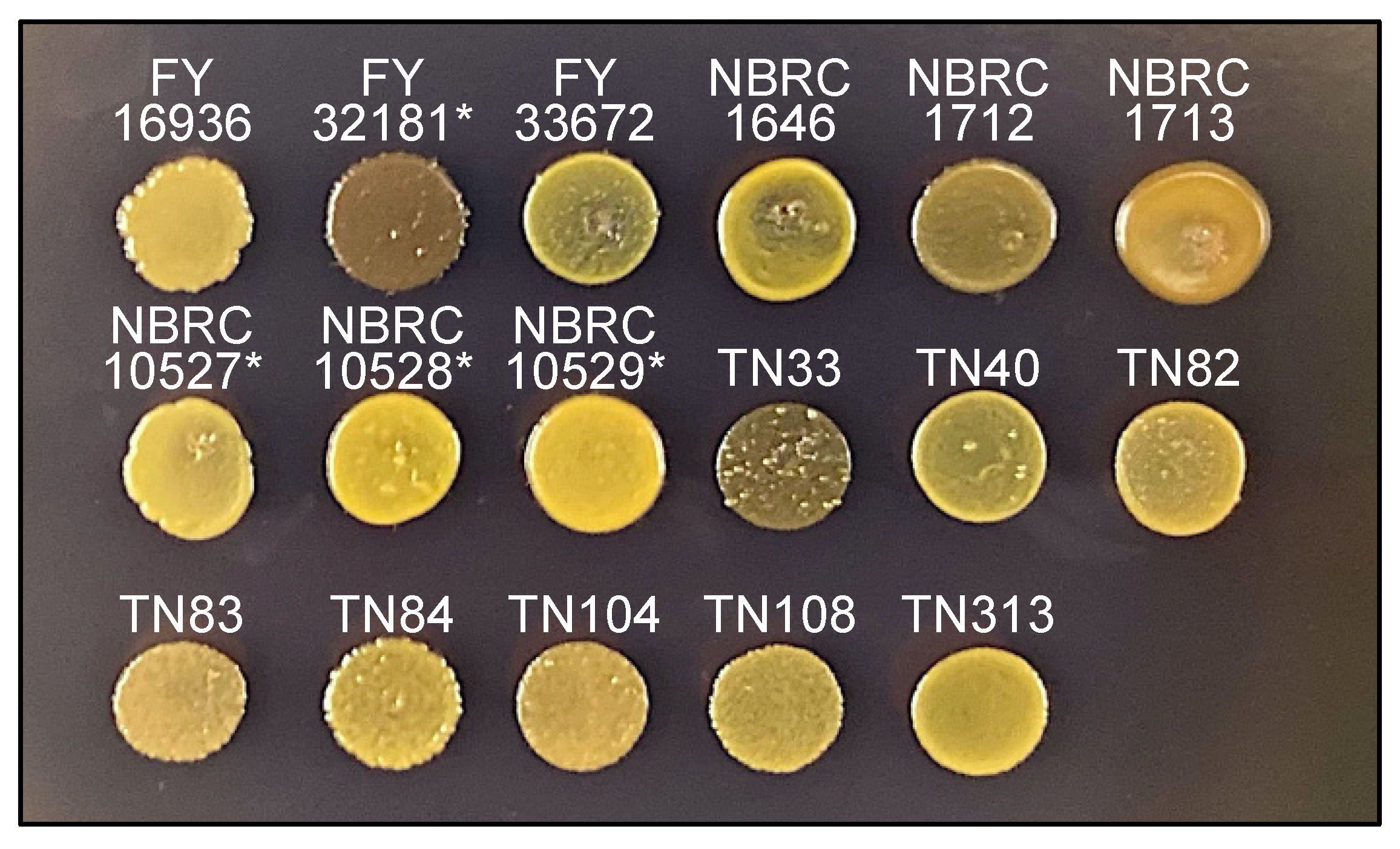
2.8. Iodine Staining
2.9. Plasmid Construction and Transformation
2.10. Shmoo Formation by Pheromone Treatment
2.11. Data Availability
3. Results
3.1. Isolation of the Fission Yeast S. japonicus var. japonicus from Drosophila
3.2. Heat Tolerance in Most of the Strains Isolated from Drosophila
3.3. Differences in Hyphal Formation among S. japonicus Strains
3.4. Elevated Sporulation Efficiency in S. japonicus Strains Isolated from Drosophila
3.5. The TN33 Strain of S. japonicus var. japonicus Shows Iodine-Positivity
3.6. Differences in Pheromone-Related Genes among S. japonicus Strains
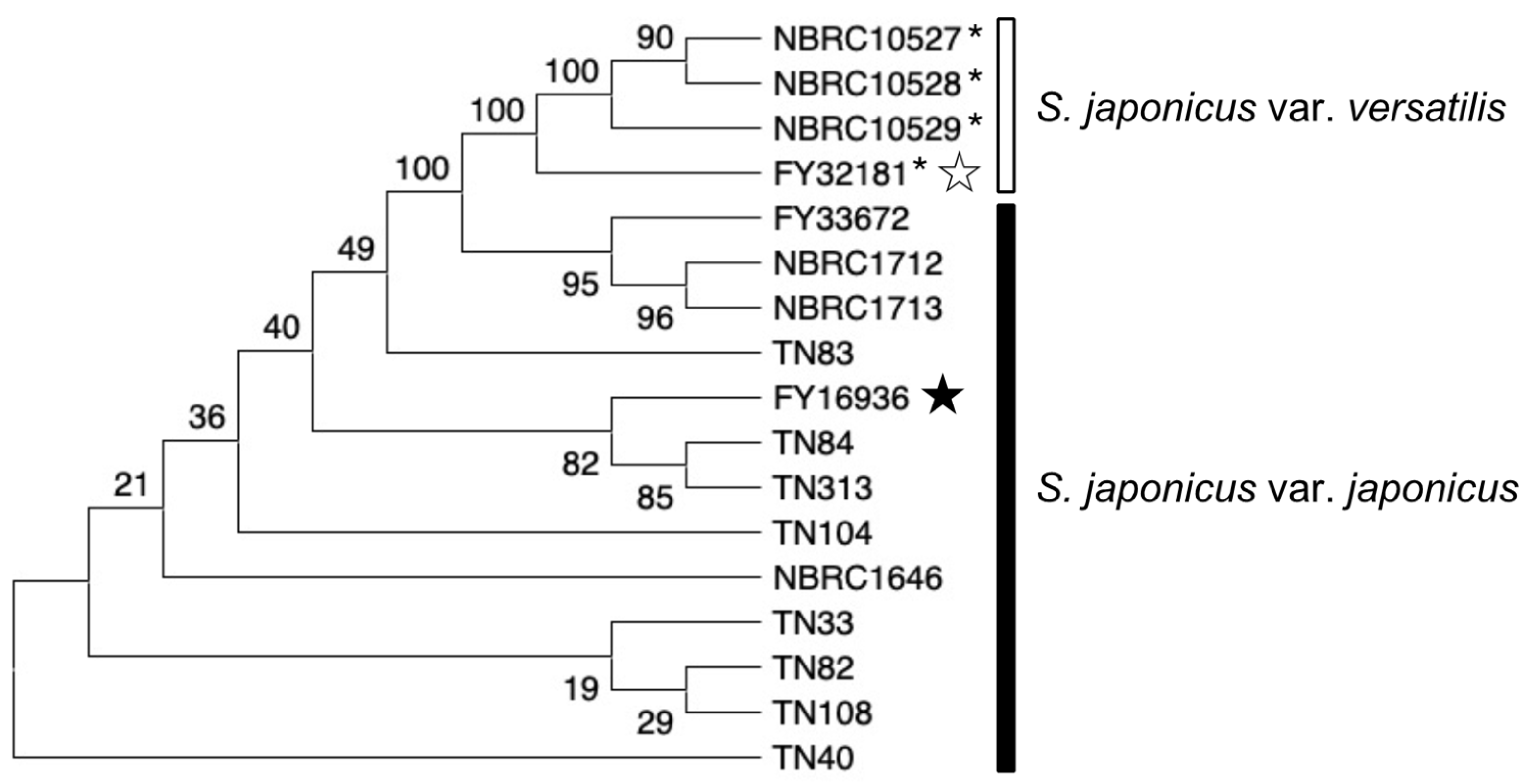
3.7. Diversify of S. japonicus Wild Strains in Nature
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mrak, E.M. The fate of yeast in the digestive tract of drosophila. Am. Nat. 1951, 85, 381–383. [Google Scholar] [CrossRef]
- Sipiczki, M. Where does fission yeast sit on the tree of life? Genome Biol. 2000, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinow, C.; Hyams, J. General cytology of fission yeasts. In Molecular Biology of the Fission Yeast; Elsevier: Amsterdam, The Netherlands, 1989; pp. 273–330. [Google Scholar]
- Aoki, K.; Hayashi, H.; Furuya, K.; Sato, M.; Takagi, T.; Osumi, M.; Kimura, A.; Niki, H. Breakage of the nuclear envelope by an extending mitotic nucleus occurs during anaphase in Schizosaccharomyces japonicus. Genes Cells 2011, 16, 911–926. [Google Scholar] [CrossRef] [PubMed]
- Yam, C.; He, Y.; Zhang, D.; Chiam, K.-H.; Oliferenko, S. Divergent strategies for controlling the nuclear membrane satisfy geometric constraints during nuclear division. Curr. Biol. 2011, 21, 1314–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niki, H. Schizosaccharomyces japonicus: The fission yeast is a fusion of yeast and hyphae. Yeast 2013, 31, 83–90. [Google Scholar] [CrossRef]
- Nemecek, J.C. Global control of dimorphism and virulence in fungi. Science 2006, 312, 583–588. [Google Scholar] [CrossRef]
- Sipiczki, M.; Takeo, K.; Yamaguchi, M.; Yoshida, S.; Miklos, I. Environmentally controlled dimorphic cycle in a fission yeast. Microbiology 1998, 144, 1319–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozaki, S.; Furuya, K.; Niki, H. The Ras1-Cdc42 pathway is involved in hyphal development of Schizosaccharomyces japonicus. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef]
- Kinnaer, C.; Dudin, O.; Martin, S.G. Yeast-to-hypha transition of Schizosaccharomyces japonicus in response to environmental stimuli. Mol. Biol. Cell 2019, 30, 975–991. [Google Scholar] [CrossRef]
- Furuya, K.; Niki, H. The DNA damage checkpoint regulates a transition between yeast and hyphal growth in Schizosaccharomyces japonicus. Mol. Cell. Biol. 2010, 30, 2909–2917. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, S.; Furuya, K.; Nozaki, S.; Aoki, K.; Niki, H. Synchronous activation of cell division by light or temperature stimuli in the dimorphic yeast Schizosaccharomyces japonicus. Eukaryot. Cell 2013, 12, 1235–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaino, T.; Tonoko, K.; Mochizuki, S.; Takashima, Y.; Kawamukai, M. Schizosaccharomyces japonicus has low levels of CoQ10 synthesis, respiration deficiency, and efficient ethanol production. Biosci. Biotechnol. Biochem. 2018, 82, 1031–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickerham, L.J.; Duprat, E. A Remarkable fission yeast, Schizosaccharomyces versatilis nov. sp. J. Bacteriol. 1945, 50, 597–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuya, K.; Niki, H. Isolation of heterothallic haploid and auxotrophic mutants of Schizosaccharomyces japonicus. Yeast 2009, 26, 221–233. [Google Scholar] [CrossRef]
- Yu, C.; Bonaduce, M.J.; Klar, A.J.S. Defining the epigenetic mechanism of asymmetric cell division of Schizosaccharomyces japonicus Yeast. Genetics 2013, 193, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Klar, A.J.S. Schizosaccharomyces japonicus yeast poised to become a favorite experimental organism for eukaryotic research. Genes Genomes Genet. 2013, 3, 1869–1873. [Google Scholar] [CrossRef] [Green Version]
- Aoki, K.; Niki, H. Transformation of Schizosaccharomyces japonicus. Cold Spring Harb. Protoc. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Aoki, K.; Nakajima, R.; Furuya, K.; Niki, H. Novel episomal vectors and a highly efficient transformation procedure for the fission yeast Schizosaccharomyces japonicus. Yeast 2010, 27, 1049–1060. [Google Scholar] [CrossRef] [Green Version]
- Furuya, K.; Niki, H. Mating, spore dissection, and selection of diploid cells in Schizosaccharomyces japonicus. Cold Spring Harb. Protoc. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Rhind, N.; Chen, Z.; Yassour, M.; Thompson, D.A.; Haas, B.J.; Habib, N.; Wapinski, I.; Roy, S.; Lin, M.F.; Heiman, D.I.; et al. Comparative Functional Genomics of the Fission Yeasts. Science 2011, 332, 930–936. [Google Scholar] [CrossRef] [Green Version]
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R.R. The ecology of insect—Yeast rela-tionships and its relevance to human industry. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172733. [Google Scholar] [CrossRef] [Green Version]
- Buzzini, P.; Lachance, M.A.; Yurkov, A. Yeasts in Natural Ecosystems: Diversity; Springer: Cham, Switzerland, 2017; ISBN 9783319626833. [Google Scholar]
- Reuter, M.; Bell, G.; Greig, D. Increased outbreeding in yeast in response to dispersal by an insect vector. Curr. Biol. 2007, 17, R81–R83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanini, I.; Dapporto, L.; Berná, L.; Polsinelli, M.; Turillazzi, S.; Cavalieri, D. Social wasps are a Saccharomyces mating nest. Proc. Natl. Acad. Sci. USA 2016, 113, 2247–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.; Rawling, S.; Stratford, M.; Steels, H.; Novodvorska, M.; Archer, D.; Chandra, S. Yeast identification: Reassessment of assimilation tests as sole universal identifiers. Lett. Appl. Microbiol. 2011, 53, 503–508. [Google Scholar] [CrossRef]
- Moriya, H.; Shimizu-Yoshida, Y.; Kitano, H. In vivo robustness analysis of cell division cycle genes in Saccharomyces cerevisiae. PLoS Genet. 2006, 2, e111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seike, T.; Niki, H. Mating response and construction of heterothallic strains of the fission yeast Schizosaccharomyces octosporus. FEMS Yeast Res. 2017, 17, 1–14. [Google Scholar] [CrossRef]
- Seike, T.; Shimoda, C.; Niki, H. Asymmetric diversification of mating pheromones in fission yeast. PLoS Biol. 2019, 17, e3000101. [Google Scholar] [CrossRef] [Green Version]
- Gutz, H.; Heslot, H.; Leupold, U.L.N. Handbook of Genetics; King, R., Ed.; Plenum Press: New York, NY, USA, 1974. [Google Scholar]
- Richard, E. Fission yeast in general genetics. In The Molecular Biology of Schizosaccharomyces Pombe; Springer: Berlin/Heidelberg, Germany, 1974; pp. 1–12. [Google Scholar]
- Ladds, G.; Rasmussen, M.; Davey, J. Characterisation of Sxa2, a protease involved in pheromone communication in fission yeast. Biochem. Soc. Trans. 1995, 23, 565S. [Google Scholar] [CrossRef] [Green Version]
- Ladds, G.; Rasmussen, E.M.; Young, T.; Nielsen, O.; Davey, J. The sxa2-dependent inactivation of the P-factor mating pheromone in the fission yeast Schizosaccharomyces pombe. Mol. Microbiol. 1996, 20, 35–42. [Google Scholar] [CrossRef]
- Imai, Y.; Yamamoto, M. Schizosaccharomyces pombe sxa1+ and sxa2+ encode putative proteases involved in the mating response. Mol. Cell. Biol. 1992, 12, 1827–1834. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular evolutionary genetics analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Hamby, K.A.; Hernández, A.; Boundy-Mills, K.; Zalom, F.G. Associations of yeasts with spotted-wing drosophila (drosophila suzukii; diptera: Drosophilidae) in cherries and raspberries. Appl. Environ. Microbiol. 2012, 78, 4869–4873. [Google Scholar] [CrossRef] [Green Version]
- Chandler, J.A.; Eisen, J.A.; Kopp, A. Yeast communities of diverse drosophila species: Comparison of two symbiont groups in the same hosts. Appl. Environ. Microbiol. 2012, 78, 7327–7336. [Google Scholar] [CrossRef] [Green Version]
- Koerte, S.; Keesey, I.W.; Easson, M.L.A.E.; Gershenzon, J.; Hansson, B.S.; Knaden, M. Variable dependency on associated yeast communities influences host range in drosophila species. Oikos 2020, 129, 964–982. [Google Scholar] [CrossRef] [Green Version]
- Coluccio, A.E.; Rodriguez, R.K.; Kernan, M.J.; Neiman, A.M. The yeast spore wall enables spores to survive passage through the digestive tract of drosophila. PLoS ONE 2008, 3, e2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briza, P.; Ellinger, A.; Winkler, G.; Breitenbach, M. Chemical composition of the yeast ascospore wall. The second outer layer consists of chitosan. J. Biol. Chem. 1988, 263, 11569–11574. [Google Scholar] [CrossRef]
- Briza, P.; Winkler, G.; Kalchhauser, H.; Breitenbach, M. Dityrosine is a prominent component of the yeast ascospore wall. A proof of its structure. J. Biol. Chem. 1986, 261, 4288–4294. [Google Scholar] [CrossRef]
- Fukunishi, K.; Miyakubi, K.; Hatanaka, M.; Otsuru, N.; Hirata, A.; Shimoda, C.; Nakamura, T. The fission yeast spore is coated by a proteinaceous surface layer comprising mainly Isp3. Mol. Biol. Cell 2014, 25, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Dawes, I.W.; Hardie, I.D. Selective killing of vegetative cells in sporulated yeast cultures by exposure to diethyl ether. Mol. Genet. Genom. 1974, 131, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Briza, P.; Breitenbach, M.; Ellinger, A.; Segall, J. Isolation of two developmentally regulated genes involved in spore wall maturation in Saccharomyces cerevisiae. Genes Dev. 1990, 4, 1775–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Variety | Origin | Source | Reference |
|---|---|---|---|---|
| FY16936 | S. japonicus var. japonicus | Japan, Kyushu University | strawberry | NBRP |
| FY32181 * | S. japonicus var. versatilis | USA | home-canned grape juice | NBRP |
| FY33672 | S. japonicus var. japonicus | Japan, Suzuka College | NBRP | |
| NBRC1646 | S. japonicus var. japonicus | Japan | slime flux of tree | NBRC |
| NBRC1712 | S. japonicus var. japonicus | Unknown | NBRC | |
| NBRC1713 | S. japonicus var. japonicus | Unknown | NBRC | |
| NBRC10527 * | S. japonicus var. versatilis | Unknown | grape juice | NBRC |
| NBRC10528 * | S. japonicus var. versatilis | Portugal | white wine | NBRC |
| NBRC10529 * | S. japonicus var. versatilis | Unknown | slime flux, Ulmus carpinifolia | NBRC |
| TN33 | S. japonicus var. japonicus | Japan, RIKEN | Drosophila | This study |
| TN40 | S. japonicus var. japonicus | Japan, RIKEN | Drosophila | This study |
| TN82 | S. japonicus var. japonicus | Japan, RIKEN | Drosophila | This study |
| TN83 | S. japonicus var. japonicus | Japan, RIKEN | Drosophila | This study |
| TN84 | S. japonicus var. japonicus | Japan, RIKEN | Drosophila | This study |
| TN104 | S. japonicus var. japonicus | Japan, RIKEN | Drosophila | This study |
| TN108 | S. japonicus var. japonicus | Japan, RIKEN | Drosophila | This study |
| TN313 | S. japonicus var. japonicus | Japan, Osaka University | Drosophila | This study |
| Gene | Amino acid Substitution | No. of Strains | Strain |
|---|---|---|---|
| map2 (SJAG_00781) | WT | 15 | FY16936, FY32181 *, FY33672, NBRC1646, NBRC1712, NBRC1713, NBRC10527 *, NBRC10529 *, TN33, TN40, TN82, TN83, TN84, TN104, TN108 |
| S22N/N32V | 1 | NBRC10528 * | |
| A29S | 1 | TN313 | |
| map3 (SJAG_01345) | WT | 11 | FY16936, FY33672, NBRC1646, NBRC1713, TN40, TN82, TN83, TN84, TN104, TN108, TN313 |
| S3P/V4E/M27L/V43L/I54V/I59V/F117L/I121V/I125V/I179V/T222S/M223L/M261L | 4 | FY32181 *, NBRC10527 *, NBRC10528 *, NBRC10529 * | |
| A44S/L169V | 1 | NBRC1712 | |
| I328V | 1 | TN33 | |
| mam2 (SJAG_01928) | WT | 10 | FY16936, FY32181 *, TN33, TN40, TN82, TN83, TN84, TN104, TN108, TN313 |
| A13T/T28V/V170I | 1 | FY33672 | |
| A185G | 1 | NBRC1646 | |
| V170I | 1 | NBRC1712 | |
| A13T/V170I | 2 | NBRC1713, NBRC10527 * | |
| A13T/V170I/R319G | 1 | NBRC10528 * | |
| A13T/V170I/S277W/L283F R319G/S330F | 1 | NBRC10529 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seike, T.; Sakata, N.; Matsuda, F.; Furusawa, C. Elevated Sporulation Efficiency in Fission Yeast Schizosaccharomyces japonicus Strains Isolated from Drosophila. J. Fungi 2021, 7, 350. https://doi.org/10.3390/jof7050350
Seike T, Sakata N, Matsuda F, Furusawa C. Elevated Sporulation Efficiency in Fission Yeast Schizosaccharomyces japonicus Strains Isolated from Drosophila. Journal of Fungi. 2021; 7(5):350. https://doi.org/10.3390/jof7050350
Chicago/Turabian StyleSeike, Taisuke, Natsue Sakata, Fumio Matsuda, and Chikara Furusawa. 2021. "Elevated Sporulation Efficiency in Fission Yeast Schizosaccharomyces japonicus Strains Isolated from Drosophila" Journal of Fungi 7, no. 5: 350. https://doi.org/10.3390/jof7050350