
In Vivo Efficacy of Lacticaseibacillus rhamnosus L8020 in a Mouse Model of Oral Candidiasis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Animals
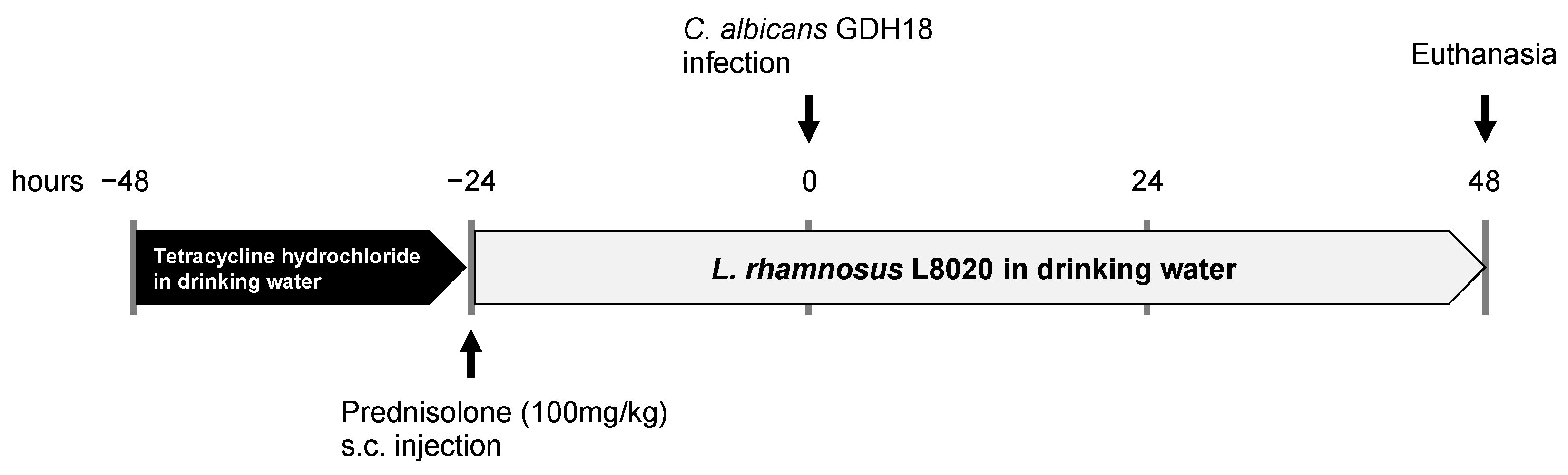
2.3. In Vivo Oral Candidiasis Model
2.4. Histological Assessments
2.5. Real-Time Quantitative RT-PCR Analysis
2.6. Statistical Analysis
3. Results
3.1. Anti-Fungal Efficacy of L. rhamnosus L8020 in a Mouse Model of Oral Candidiasis
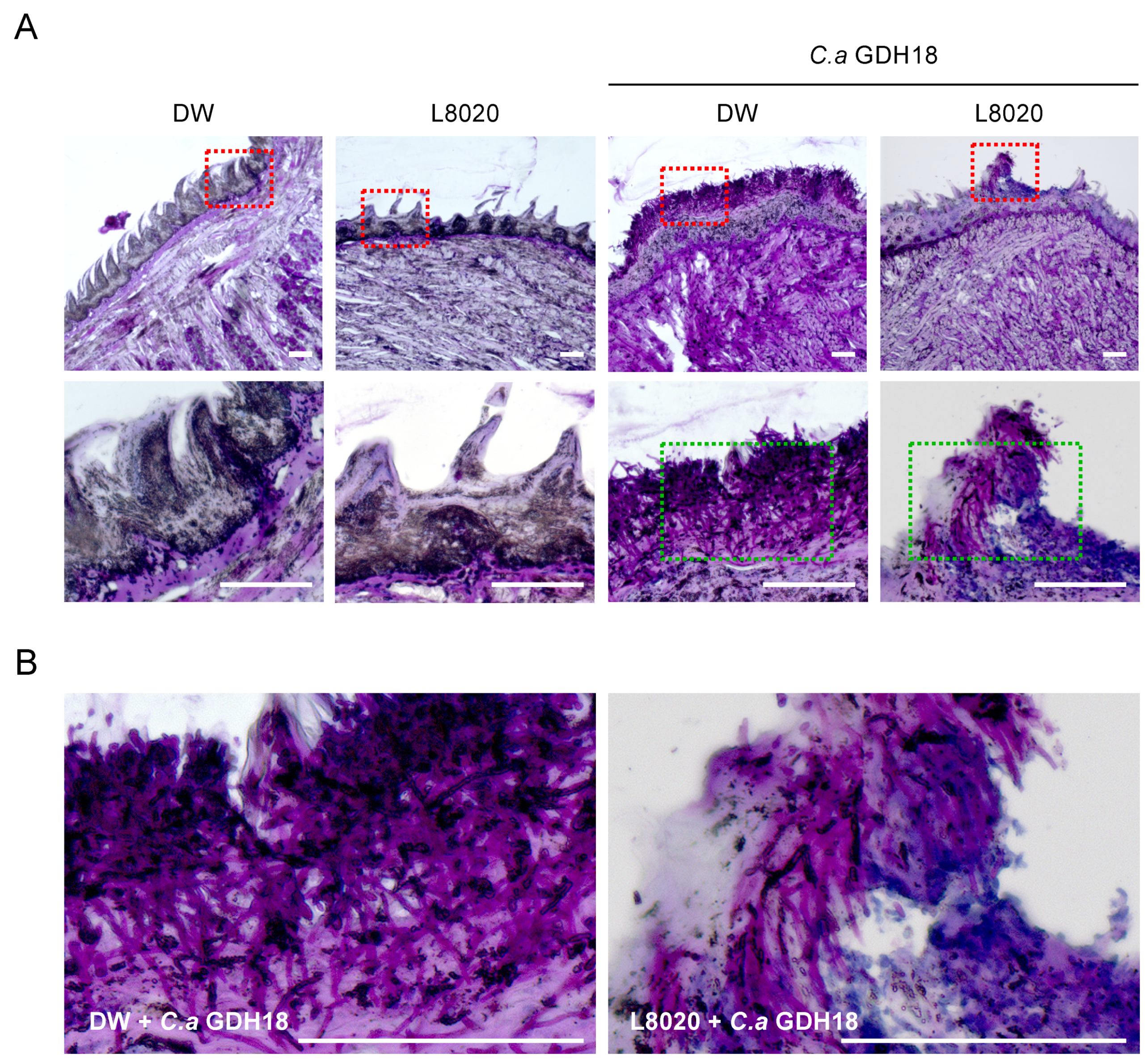
3.2. Histological Assessment of the Tongue Dorsum
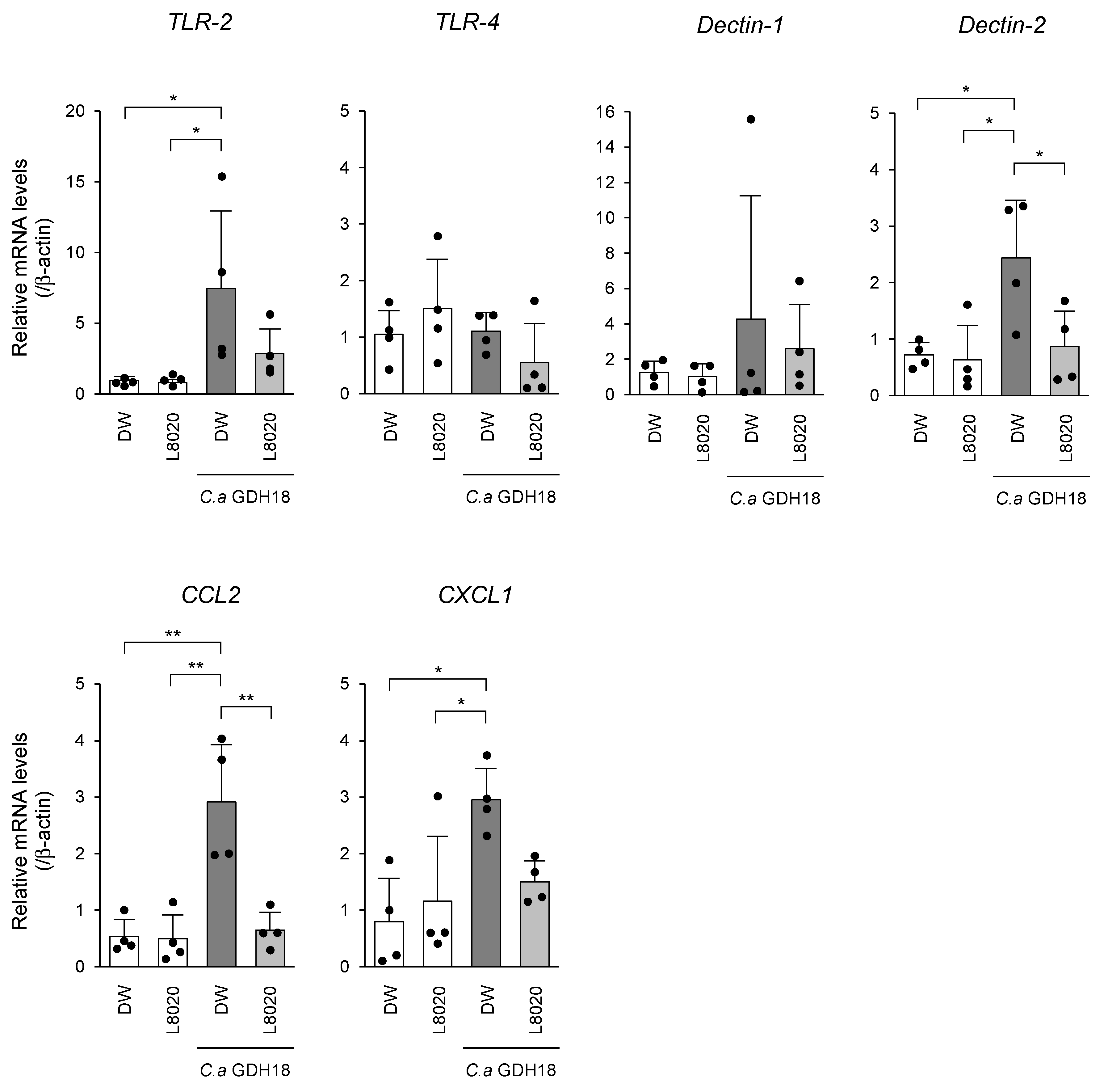
3.3. In Vivo Analyses of PRR and Chemokine Expression Levels
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samaranayake, L.P.; Leung, W.K.; Jin, L. Oral mucosal fungal infections. Periodontology 2000 2009, 49, 39–59. [Google Scholar] [CrossRef]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Fidel, P.L., Jr. Candida-host interactions in HIV disease: Implications for oropharyngeal candidiasis. Adv. Dent. Res. 2011, 23, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Zhou, M.; Young, A.; Zhao, W.; Yan, Z. In vivo effectiveness and safety of probiotics on prophylaxis and treatment of oral candidiasis: A systematic review and meta-analysis. BMC Oral Health 2019, 19, 140. [Google Scholar] [CrossRef] [PubMed]
- Vila, T.; Sultan, A.S.; Montelongo-Jauregui, D.; Jabra-Rizk, M.A. Oral candidiasis: A disease of opportunity. J. Fungi 2020, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO. Report on Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; FAO; WHO: Rome, Italy, 2001; p. 2. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Nikawa, H.; Tomiyama, Y.; Hiramatsu, M.; Yushita, K.; Takamoto, Y.; Ishi, H.; Mimura, S.; Hiyama, A.; Sasahara, H.; Kawahara, K.; et al. Bovine milk fermented with Lactobacillus rhamnosus L8020 decreases the oral carriage of mutans streptococci and the burden of periodontal pathogens. J. Investig. Clin. Dent. 2011, 2, 187–196. [Google Scholar] [CrossRef]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Salvetti, E.; Harris, H.M.B.; Felis, G.E.; O’Toole, P.W. Comparative genomics of the genus lactobacillus reveals robust phylogroups that provide the basis for reclassification. Appl. Environ. Microbiol. 2018, 84, e00993-18. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, V.H.; Silva, E.G.; Paula, C.R.; Ishikawa, K.H.; Nakamae, A.E. Treatment with probiotics in experimental oral colonization by Candida albicans in murine model (DBA/2). Oral Dis. 2012, 18, 260–264. [Google Scholar] [CrossRef]
- Ribeiro, F.C.; Rossoni, R.D.; de Barros, P.P.; Santos, J.D.; Fugisaki, L.R.O.; Leão, M.P.V.; Junqueira, J.C. Action mechanisms of probiotics on Candida spp. and candidiasis prevention: An update. J. Appl. Microbiol. 2020, 129, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Endo, K.; Mine, Y.; Shuto, T.; Taji, T.; Murayama, T.; Nikawa, H. Comprehensive analysis of transcriptional profiles in oral epithelial-like cells stimulated with oral probiotic Lactobacillus spp. Arch. Oral Biol. 2020, 118, 104832. [Google Scholar] [CrossRef]
- Yuki, O.; Furutani, C.; Mizota, Y.; Wakita, A.; Mimura, S.; Kihara, T.; Ohara, M.; Okada, Y.; Okada, M.; Nikawa, H. Effect of bovine milk fermented with Lactobacillus rhamnosus L8020 on periodontal disease in individuals with intellectual disability: A randomized clinical trial. J. Appl. Oral Sci. 2019, 29, e20180564. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Fukunaga, M.; Kuda, T.; Goto, M.; Chiaraluce, G.; Hoshiba, H.; Takahashi, H.; Kimura, B. Detection and isolation of protein susceptible indigenous bacteria affected by dietary milk-casein, albumen and soy-protein in the caecum of ICR mice. Int. J. Biol. Macromol. 2020, 144, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Takakura, N.; Sato, Y.; Ishibashi, H.; Oshima, H.; Uchida, K.; Yamaguchi, H.; Abe, S. A novel murine model of oral candidiasis with local symptoms characteristic of oral thrush. Microbiol. Immunol. 2003, 47, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Ishijima, S.A.; Hayama, K.; Burton, J.P.; Reid, G.; Okada, M.; Matsushita, Y.; Abe, S. Effect of Streptococcus salivarius K12 on the in vitro growth of Candida albicans and its protective effect in an oral candidiasis model. Appl. Environ. Microbiol. 2012, 78, 2190–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.Y.; Samaranayake, Y.H.; Samaranayake, L.P.; Nikawa, H. In vitro susceptibility of Candida species to lactoferrin. Med. Mycol. 1999, 37, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.; Viale, G.; Rescigno, M. Probiotic and postbiotic activity in health and disease: Comparison on a novel polarised ex-vivo organ culture model. Gut 2012, 61, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Carmello, J.C.; Alves, F.; Basso, F.G.; de Souza Costa, C.A.; Bagnato, V.S.; Mima, E.G.D.O.; Pavarina, A.C. Treatment of oral candidiasis using photodithazine®-mediated Photodynamic Therapy in vivo. PLoS ONE 2016, 11, e0156947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leão, M.V.P.; Tavares, T.A.A.; e Silva, C.R.G.; Dos Santos, S.S.F.; Junqueira, J.C.; de Oliveira, L.D.; Jorge, A.O.C. Lactobacillus rhamnosus intake can prevent the development of Candidiasis. Clin. Oral Investig. 2018, 22, 2511–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Sayáns, M.; Beiro-Fuentes, R.; Otero-Rey, E.M.; Chamorro-Petronacci, C.M.; Gándara-Vila, P.; Somoza-Martín, J.M.; García-García, A.; Blanco-Carrión, A. Efficacy of different formulations of nystatin in an experimental model of oral candidiasis in sialoadenectomized rats. J. Dent. Sci. 2021, 16, 123–130. [Google Scholar] [CrossRef]
- Ribeiro, F.C.; Junqueira, J.C.; Dos Santos, J.D.; de Barros, P.P.; Rossoni, R.D.; Shukla, S.; Fuchs, B.B.; Shukla, A.; Mylonakis, E. Development of probiotic formulations for oral candidiasis prevention: Gellan gum as a carrier to deliver lactobacillus paracasei 28.4. Antimicrob. Agents Chemother. 2020, 64, e02323-19. [Google Scholar] [CrossRef]
- Authier, H.; Salon, M.; Rahabi, M.; Bertrand, B.; Blondeau, C.; Kuylle, S.; Holowacz, S.; Coste, A. Oral Administration of Lactobacillus helveticus LA401 and Lactobacillus gasseri LA806 Combination Attenuates Oesophageal and Gastrointestinal Candidiasis and Consequent Gut Inflammation in Mice. J. Fungi 2021, 7, 57. [Google Scholar] [CrossRef]
- Liao, H.; Liu, S.; Wang, H.; Su, H.; Liu, Z. Enhanced antifungal activity of bovine lactoferrin-producing probiotic Lactobacillus casei in the murine model of vulvovaginal candidiasis. BMC Microbiol. 2019, 19, 7. [Google Scholar] [CrossRef]
- De Gregorio, P.R.; Silva, J.A.; Marchesi, A.; Nader-Macías, M.E.F. Anti-Candida activity of beneficial vaginal lactobacilli in in vitro assays and in a murine experimental model. FEMS Yeast Res. 2019, 19, foz008. [Google Scholar] [CrossRef] [Green Version]
- Sanders, M.E. Probiotics: Considerations for human health. Nutr. Rev. 2003, 61, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Moyes, D.L.; Wilson, D.; Richardson, J.P.; Mogavero, S.; Tang, S.X.; Wernecke, J.; Höfs, S.; Gratacap, R.L.; Robbins, J.; Runglall, M.; et al. Candidalysin is a fungal peptide toxin critical for mucosal infection. Nature 2016, 532, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Santos, N.; Gaffen, S.L. Th17 cells in immunity to Candida albicans. Cell Host Microbe 2012, 11, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Bojang, E.; Ghuman, H.; Kumwenda, P.; Hall, R.A. Immune Sensing of Candida albicans. J. Fungi 2021, 7, 119. [Google Scholar] [CrossRef]
- Zhang, S.; Li, J.; Jia, X.; Wu, Y. The expression of toll-like receptor 2 and 4 mRNA in local tissues of model of oropharyngeal candidiasis in mice. J. Huazhong Univ. Sci. Technol. Med. Sci. 2004, 24, 639–641. [Google Scholar] [CrossRef]
- Conti, H.R.; Shen, F.; Nayyar, N.; Stocum, E.; Sun, J.N.; Lindemann, M.J.; Ho, A.W.; Hai, J.H.; Yu, J.J.; Jung, J.W.; et al. Th17 cells and IL-17 receptor signaling are essential for mucosal host defense against oral candidiasis. J. Exp. Med. 2009, 206, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Saunus, J.M.; Wagner, S.A.; Matias, M.A.; Hu, Y.; Zaini, Z.M.; Farah, C.S. Early activation of the interleukin-23-17 axis in a murine model of oropharyngeal candidiasis. Mol. Oral Microbiol. 2010, 25, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Conti, H.R.; Bruno, V.M.; Childs, E.E.; Daugherty, S.; Hunter, J.P.; Mengesha, B.G.; Saevig, D.L.; Hendricks, M.R.; Coleman, B.M.; Brane, L.; et al. IL-17 receptor signaling in oral epithelial cells is critical for protection against oropharyngeal candidiasis. Cell Host Microbe 2016, 20, 606–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, R.; Mine, Y.; Yumisashi, Y.; Yoshioka, R.; Hamaoka, M.; Taji, T.; Murayama, T.; Nikawa, H. In Vivo Efficacy of Lacticaseibacillus rhamnosus L8020 in a Mouse Model of Oral Candidiasis. J. Fungi 2021, 7, 322. https://doi.org/10.3390/jof7050322
Ito R, Mine Y, Yumisashi Y, Yoshioka R, Hamaoka M, Taji T, Murayama T, Nikawa H. In Vivo Efficacy of Lacticaseibacillus rhamnosus L8020 in a Mouse Model of Oral Candidiasis. Journal of Fungi. 2021; 7(5):322. https://doi.org/10.3390/jof7050322
Chicago/Turabian StyleIto, Rei, Yuichi Mine, Yoshie Yumisashi, Reina Yoshioka, Misa Hamaoka, Tsuyoshi Taji, Takeshi Murayama, and Hiroki Nikawa. 2021. "In Vivo Efficacy of Lacticaseibacillus rhamnosus L8020 in a Mouse Model of Oral Candidiasis" Journal of Fungi 7, no. 5: 322. https://doi.org/10.3390/jof7050322