
Genetic Diversity and Pathogenicity of Rhizoctonia spp. Isolates Associated with Red Cabbage in Samsun (Turkey)

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation of Rhizoctonia Isolates
2.2. DNA Extraction
2.3. ITS Sequencing
2.4. iPBS Amplification Analyses
2.5. Pathogenicity Assays
2.6. Resistance Response Classification
2.7. Statistical Analysis
3. Results
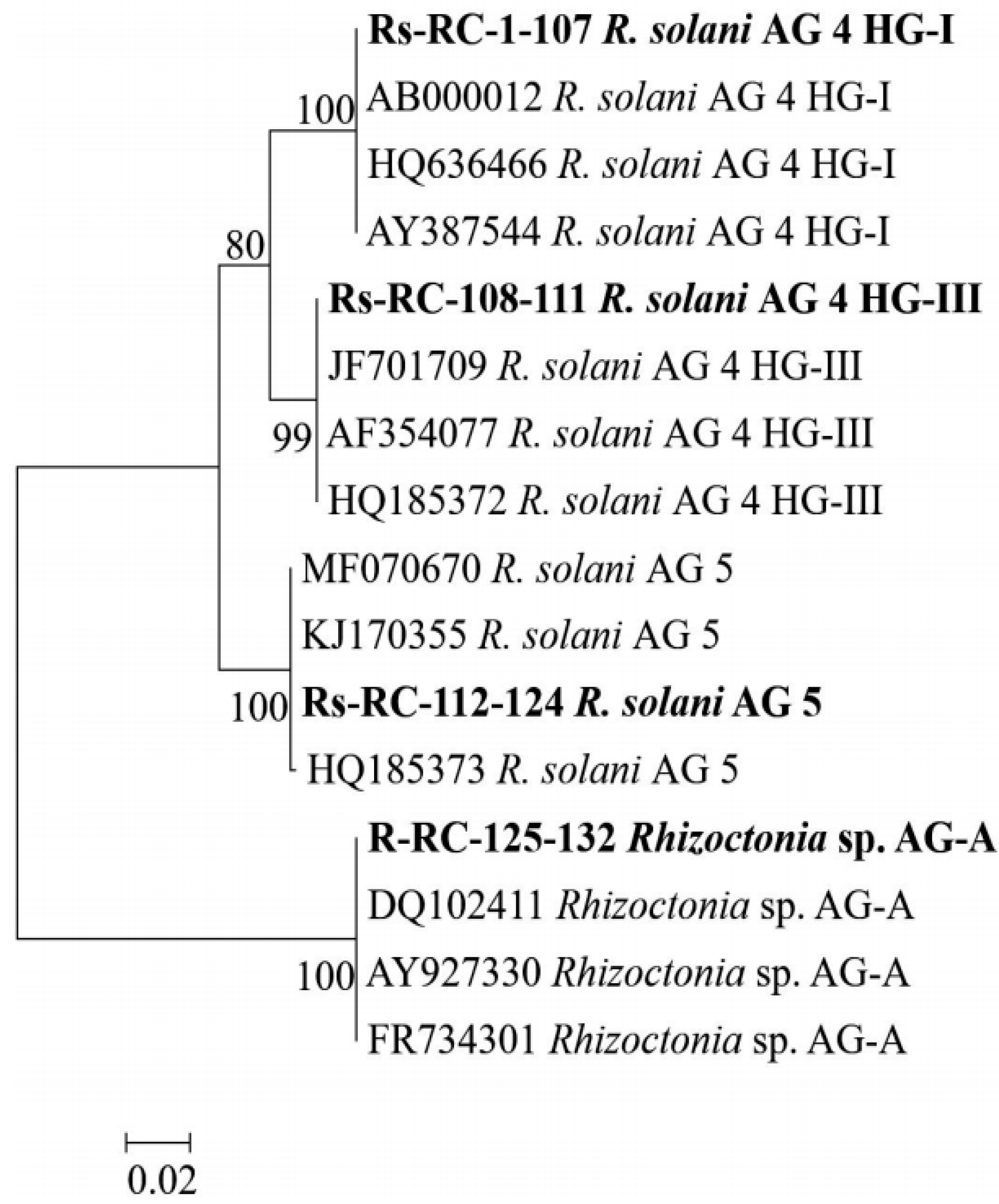
3.1. Rhizoctonia Species and AGs on Red Cabbage
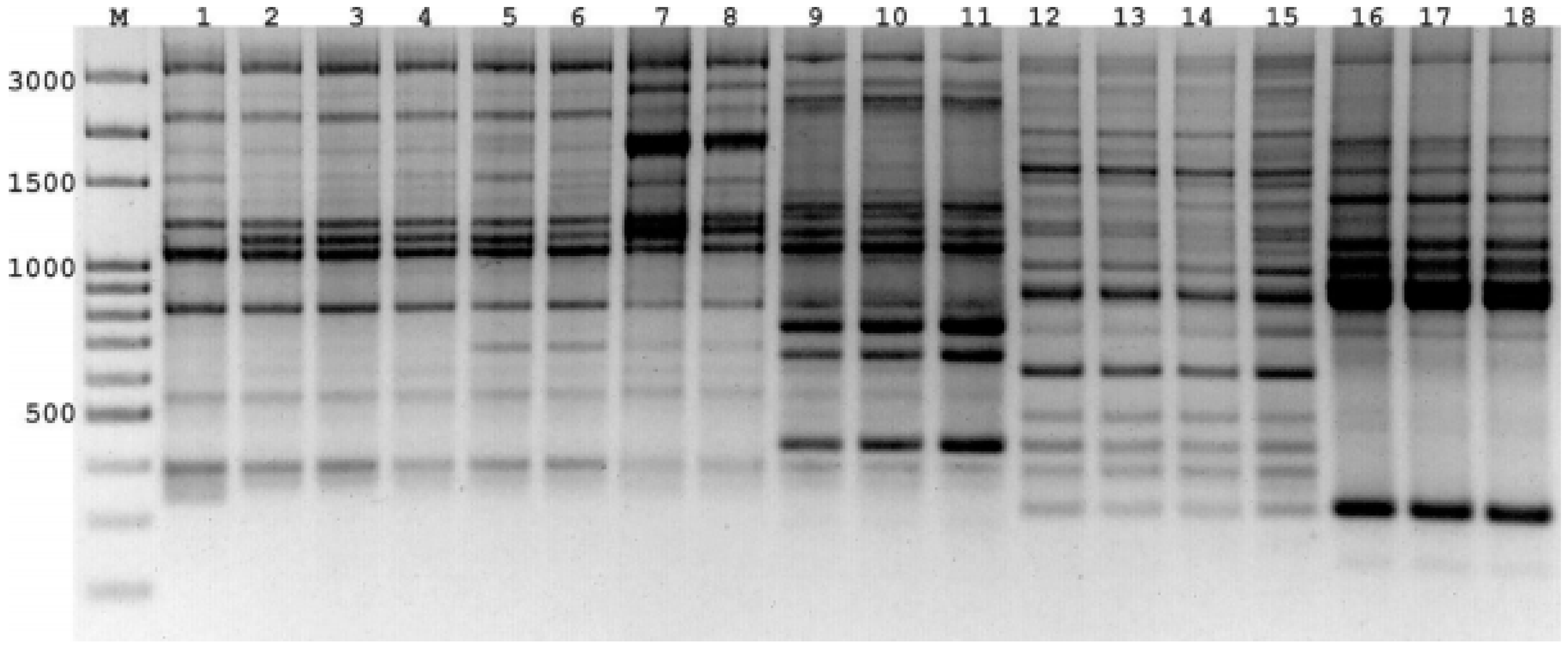
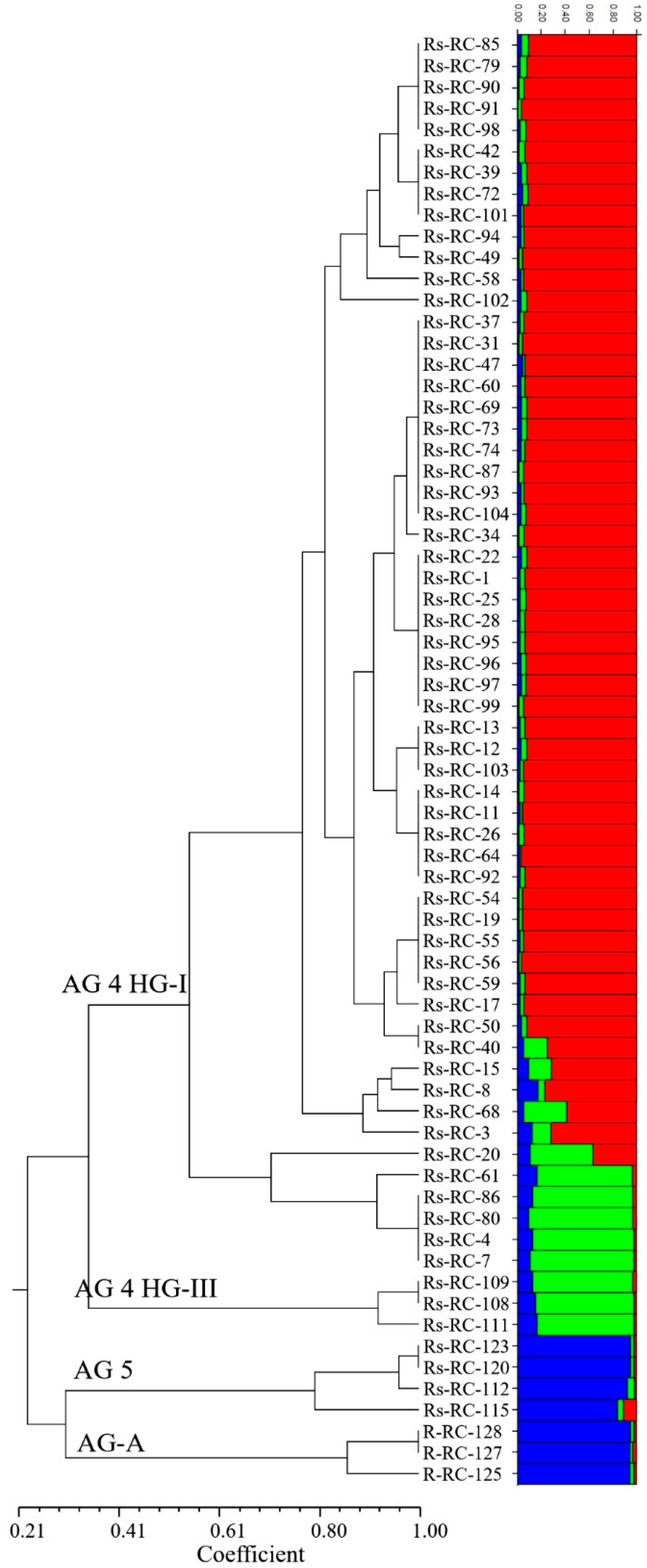
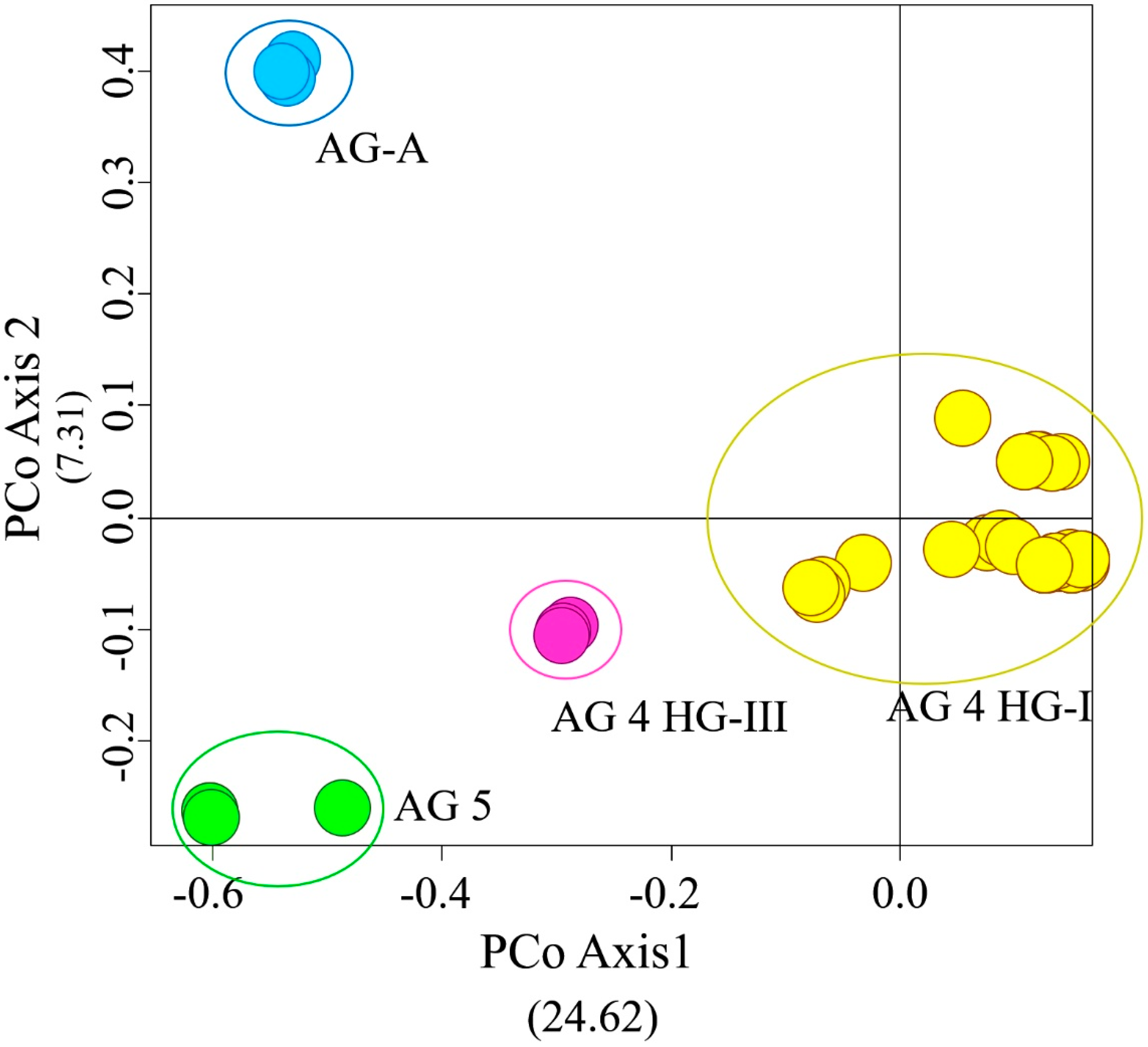
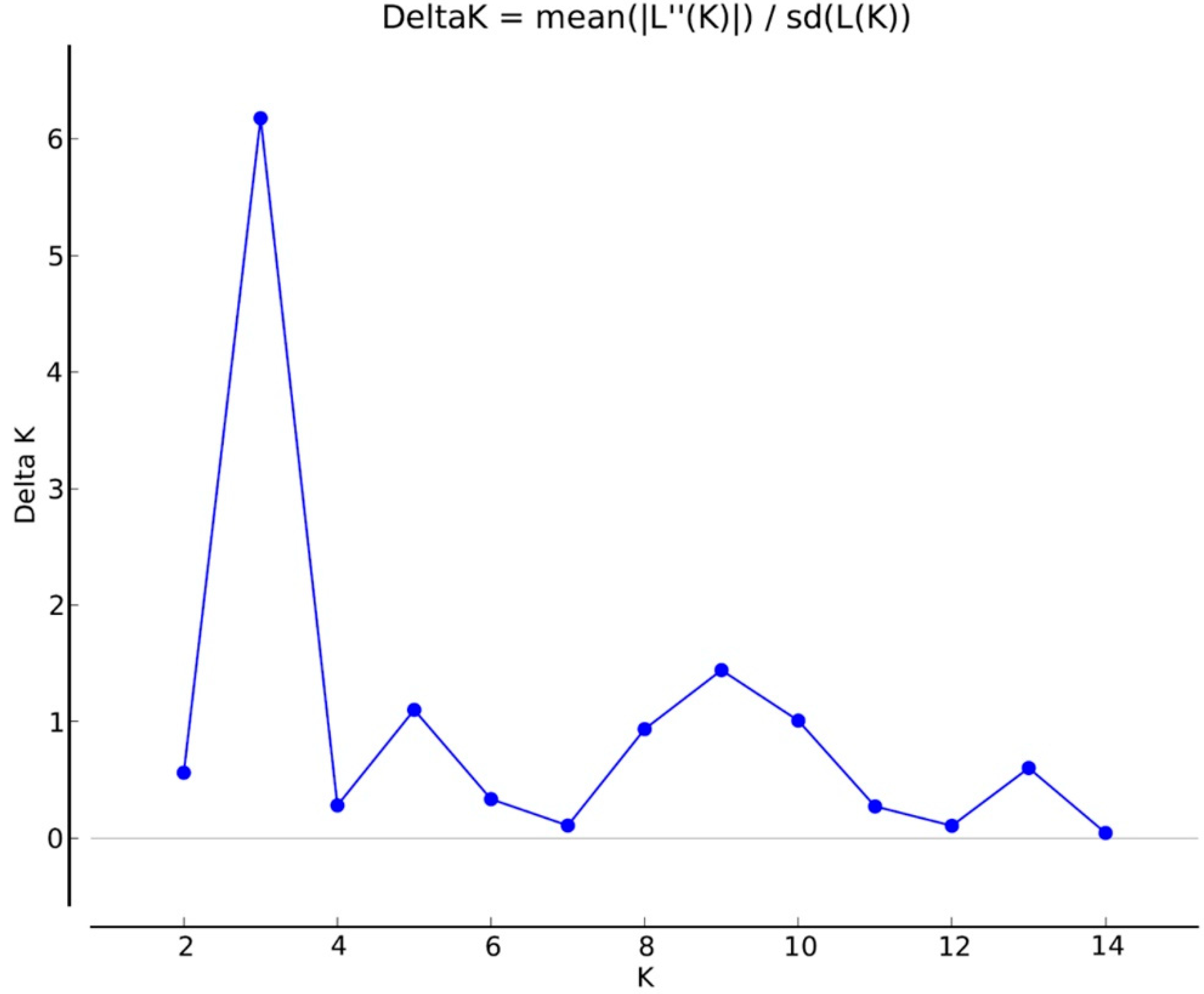
3.2. iPBS Amplification Analysis
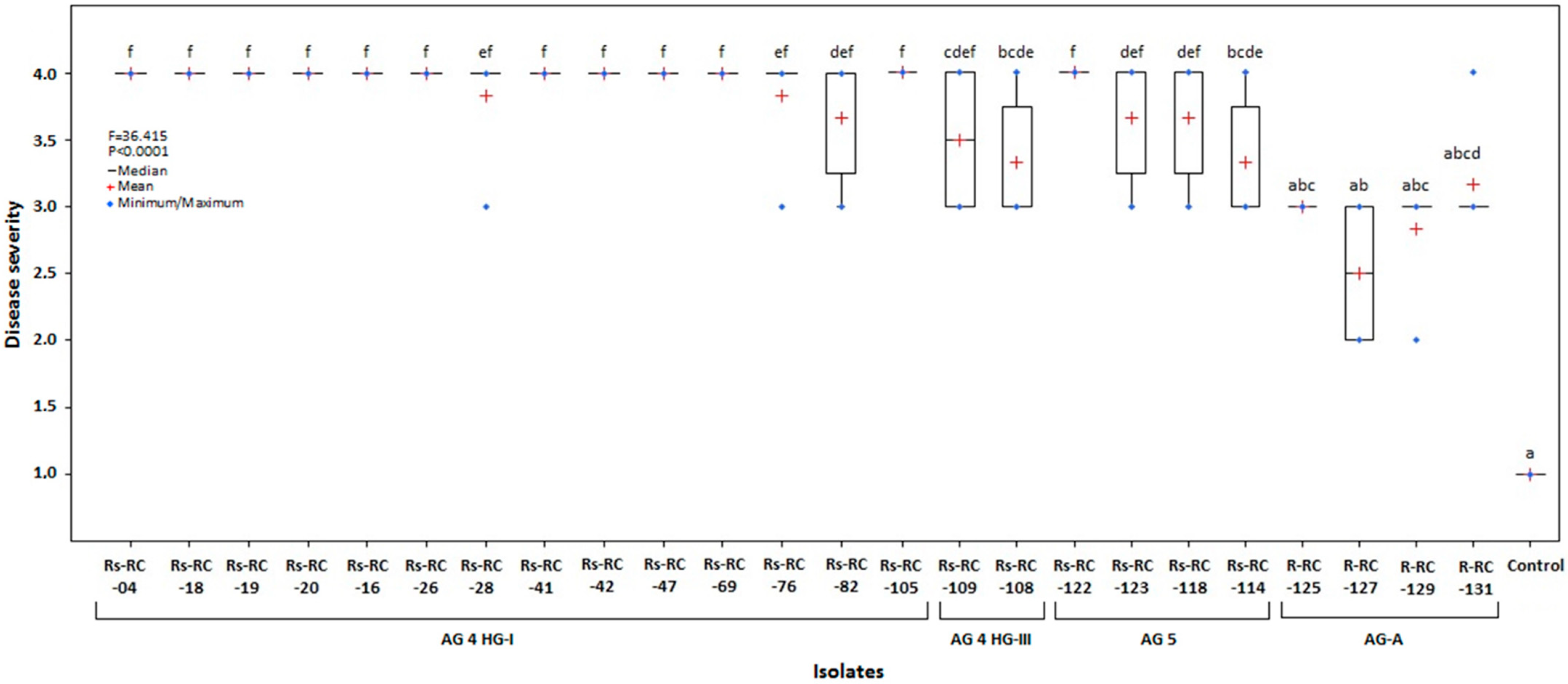
3.3. Virulence of the Isolates
3.4. Host Response of Some Commercial Cultivars to Different AGs of Rhizoctonia
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#data (accessed on 1 March 2021).
- Monteiro, A.; Lunn, T. Trends and Perspectives of Vegetable Brassica Breeding World-Wide. In Proceedings of the WCHR-World Conference on Horticultural Research 495, Rome, Italy, 17–20 June 1998; pp. 273–280. [Google Scholar]
- Balkaya, A.; Yanmaz, R.; Apaydin, A.; Kar, H. Morphological characterisation of white head cabbage (Brassica oleracea var. capitata subvar. alba) genotypes in turkey. N. Z. J. Crop Hortic. Sci. 2005, 33, 333–341. [Google Scholar] [CrossRef] [Green Version]
- TUIK. Turkish Statistical Institute: Crop Production Statistics. Available online: https://biruni.tuik.gov.tr/medas (accessed on 1 March 2021).
- Rimmer, S.R.; Shattuck, V.I.; Buchwaldt, L. Compendium of Brassica Diseases; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2007. [Google Scholar]
- Sneh, B.; Jabaji-Hare, S.; Neate, S.; Dijst, G. Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1996. [Google Scholar] [CrossRef]
- Yang, G.H.; Chen, J.Y.; Pu, W.Q. First report of head rot of cabbage and web blight of snap bean caused by Rhizoctonia solani AG-4 HGI. Plant Pathol. 2007, 56, 351. [Google Scholar] [CrossRef]
- Hua, G.K.; Bertier, L.; Soltaninejad, S.; Hofte, M. Cropping systems and cultural practices determine the Rhizoctonia anastomosis groups associated with Brassica spp. in Vietnam. PLoS ONE 2014, 9, e111750. [Google Scholar] [CrossRef] [Green Version]
- Misawa, T.; Kubota, M.; Sasaki, J.; Kuninaga, S. First report of broccoli foot rot caused by Rhizoctonia solani AG-2-2 IV and pathogenicity comparison of the pathogen with related pathogens. J. Gen. Plant Pathol. 2014, 81, 15–23. [Google Scholar] [CrossRef]
- Saygi, S.; Erper, I.; Ozer, G.; Turkkan, M. First report of Rhizoctonia solani AG 4 HG-I causing damping off and wirestem of white cabbage in Turkey. J. Plant Pathol. 2019, 102, 587. [Google Scholar] [CrossRef] [Green Version]
- Türkkan, M.; Kılıçoğlu, M.Ç.; Erper, I. Characterization and pathogenicity of Rhizoctonia isolates collected from Brassica oleracea var. acephala in Ordu, Turkey. Phytoparasitica 2020, 48, 273–286. [Google Scholar] [CrossRef]
- Carling, D.E.; Kuninaga, S.; Brainard, K.A. Hyphal anastomosis reactions, rDNA-internal transcribed spacer sequences, and virulence levels among subsets of Rhizoctonia solani anastomosis group-2 (AG-2) and AG-BI. Phytopathology 2002, 92, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharon, M.; Sneh, B.; Kuninaga, S.; Hyakumachi, M.; Naito, S. Classification of Rhizoctonia spp. using rDNA-ITS sequence analysis supports the genetic basis of the classical anastomosis grouping. Mycoscience 2008, 49, 93–114. [Google Scholar] [CrossRef]
- Aiello, D.; Guarnaccia, V.; Formica, P.T.; Hyakumachi, M.; Polizzi, G. Occurrence and characterisation of Rhizoctonia species causing diseases of ornamental plants in Italy. Eur. J. Plant Pathol. 2017, 148, 967–982. [Google Scholar] [CrossRef]
- Dong, W.; Li, Y.; Duan, C.; Li, X.; Naito, S.; Conner, R.L.; Yang, G.; Li, C. Identification of AG-V, a new anastomosis group of binucleate Rhizoctonia spp. from taro and ginger in Yunnan province. Eur. J. Plant Pathol. 2017, 148, 895–906. [Google Scholar] [CrossRef]
- Zhao, C.; Li, Y.; Liu, H.; Li, S.; Han, C.; Wu, X. A binucleate Rhizoctonia anastomosis group (AG-W) is the causal agent of sugar beet seedling damping-off disease in China. Eur. J. Plant Pathol. 2019, 155, 53–69. [Google Scholar] [CrossRef]
- Ogoshi, A. Ecology and pathogenicity of anastomosis and intraspecific groups of Rhizoctonia solani Kuhn. Annu. Rev. Phytopathol. 1987, 25, 125–143. [Google Scholar] [CrossRef]
- Kuramae, E.E. Identification of Rhizoctonia solani AG 1-IB in lettuce, AG 4 HG-I in tomato and melon, and AG 4 HG-III in broccoli and spinach, in Brazil. Eur. J. Plant Pathol. 2003, 109, 391–395. [Google Scholar] [CrossRef]
- Khangura, R.K.; Barbetti, M.J.; Sweetingham, M.W. Characterization and pathogenicity of Rhizoctonia species on canola. Plant Dis. 1999, 83, 714–721. [Google Scholar] [CrossRef] [Green Version]
- Pannecoucque, J.; Van Beneden, S.; Höfte, M. Characterization and pathogenicity of Rhizoctonia isolates associated with cauliflower in belgium. Plant Pathol. 2008, 57, 737–746. [Google Scholar] [CrossRef]
- Yang, J. Characterization, virulence, and genetic variation of Rhizoctonia solani AG-9 in Alberta. Plant Dis. 1996, 80. [Google Scholar] [CrossRef]
- Sayama, A. Occurrence of damping-off disease caused by Rhizoctonia solani ag 2-2-iiib on cabbage plug seedlings. Annu. Rep. Soc. Plant Prot. N. Jpn. 2000, 51, 54–57. [Google Scholar]
- Misawa, T.; Aoki, M. First report of Rhizoctonia solani AG-1 IC causing head rot of cabbage in Japan. New Dis. Rep. 2017, 36, 12. [Google Scholar] [CrossRef] [Green Version]
- Keinath, A.P.; Farnham, M.W. Differential cultivars and criteria for evaluating resistance to Rhizoctonia solani in seedling Brassica oleracea. Plant Dis. 1997, 81, 946–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulitz, T.C.; Okubara, P.A.; Schillinger, W.F. First report of damping-off of canola caused by Rhizoctonia solani AG 2-1 in washington state. Plant Dis. 2006, 90, 829. [Google Scholar] [CrossRef]
- Budge, G.E.; Shaw, M.W.; Colyer, A.; Pietravalle, S.; Boonham, N. Molecular tools to investigate Rhizoctonia solani distribution in soil. Plant Pathol. 2009, 58, 1071–1080. [Google Scholar] [CrossRef]
- Kalendar, R.; Shustov, A.V.; Seppanen, M.M.; Schulman, A.H.; Stoddard, F.L. Palindromic sequence-targeted (PST) PCR: A rapid and efficient method for high-throughput gene characterization and genome walking. Sci. Rep. 2019, 9, 17707. [Google Scholar] [CrossRef]
- Kalendar, R.; Muterko, A.; Boronnikova, S. Retrotransposable elements: DNA fingerprinting and the assessment of genetic diversity. Methods Mol. Biol. 2021, 2222, 263–286. [Google Scholar] [CrossRef] [PubMed]
- Ghonaim, M.; Kalendar, R.; Barakat, H.; Elsherif, N.; Ashry, N.; Schulman, A.H. High-throughput retrotransposon-based genetic diversity of maize germplasm assessment and analysis. Mol. Biol. Rep. 2020, 47, 1589–1603. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Ramakrishnan, M.; Vinod, K.K.; Kalendar, R.; Yrjälä, K.; Zhou, M. Development and deployment of high-throughput retrotransposon-based markers reveal genetic diversity and population structure of Asian bamboo. Forests 2019, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Kalendar, R.; Raskina, O.; Belyayev, A.; Schulman, A.H. Long tandem arrays of Cassandra retroelements and their role in genome dynamics in plants. Int. J. Mol. Sci. 2020, 21, 2931. [Google Scholar] [CrossRef]
- Kalendar, R.; Amenov, A.; Daniyarov, A. Use of retrotransposon-derived genetic markers to analyse genomic variability in plants. Funct. Plant Biol. 2018, 46, 15–29. [Google Scholar] [CrossRef]
- Kalendar, R.; Antonius, K.; Smýkal, P.; Schulman, A.H. IPBS: A universal method for DNA fingerprinting and retrotransposon isolation. Theor. Appl. Genet. 2010, 121, 1419–1430. [Google Scholar] [CrossRef]
- Milovanov, A.; Zvyagin, A.; Daniyarov, A.; Kalendar, R.; Troshin, L. Genetic analysis of the grapevine genotypes of the Russian Vitis ampelographic collection using iPBS markers. Genetica 2019, 147, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Pourmahdi, A.; Taheri, P. Genetic diversity of thanatephorus cucumeris infecting tomato in Iran. J. Phytopathol. 2015, 163, 19–32. [Google Scholar] [CrossRef]
- Özer, G.; Bayraktar, H.; Baloch, F.S. IPBS retrotransposons ‘a universal retrotransposons’ now in molecular phylogeny of fungal pathogens. Biochem. Syst. Ecol. 2016, 68, 142–147. [Google Scholar] [CrossRef]
- Ozer, G.; Sameeullah, M.; Bayraktar, H.; Göre, M.E. Genetic diversity among phytopathogenic Sclerotiniaceae, based on retrotransposon molecular markers. Phytopathol. Mediterr. 2017, 56, 251–258. [Google Scholar]
- Özer, G.; Bayraktar, H. Genetic diversity of Fusarium oxysporum f. sp. cumini isolates analyzed by vegetative compatibility, sequences analysis of the rDNA IGS region and iPBS retrotransposon markers. J. Plant Pathol. 2018, 100, 225–232. [Google Scholar] [CrossRef]
- Aydin, F.; Ozer, G.; Alkan, M.; Cakir, I. The utility of iPBS retrotransposons markers to analyze genetic variation in yeast. Int. J. Food Microbiol. 2020, 325, 108647. [Google Scholar] [CrossRef] [PubMed]
- Turzhanova, A.; Khapilina, O.N.; Tumenbayeva, A.; Shevtsov, V.; Raiser, O.; Kalendar, R. Genetic diversity of Alternaria species associated with black point in wheat grains. PeerJ 2020, 8, e9097. [Google Scholar] [CrossRef]
- Amaradasa, B.S.; Horvath, B.J.; Lakshman, D.K.; Warnke, S.E. DNA fingerprinting and anastomosis grouping reveal similar genetic diversity in species infecting turfgrasses in the transition zone of USA. Mycologia 2013, 105, 1190–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaradasa, B.S.; Keenan Amundsen, D.L. AFLP fingerprinting for identification of infra-species groups of Rhizoctonia solani and Waitea circinata. J. Plant Pathol. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Doungous, O.; Kalendar, R.; Filippova, N.; Ngane, B.K. Utility of iPBS retrotransposons markers for molecular characterization of African Gnetum species. Plant Biosyst. 2019, 154, 587–592. [Google Scholar] [CrossRef]
- Khapilina, O.; Raiser, O.; Danilova, A.; Shevtsov, V.; Turzhanova, A.; Kalendar, R. DNA profiling and assessment of genetic diversity of relict species Allium altaicum pall. on the territory of Altai. PeerJ 2021, 9, e10674. [Google Scholar] [CrossRef]
- Demirci, E.; Döken, M. Anastomosis groups and pathogenicity of Rhizoctonia solani Kuehn isolates from potatoes in Erzurum- Turkey. J. Turk. Phytopathol. 1993, 22, 95–102. [Google Scholar]
- Erper, I.; Karaca, G.H.; Turkkan, M.; Ozkoc, I. Characterization and pathogenicity of Rhizoctonia spp. from onion in Amasya, Turkey. J. Phytopathol. 2006, 154, 75–79. [Google Scholar] [CrossRef]
- Erper, I.; Kilicoglu, M.C.; Turkkan, M.; Onder, H. Characterization and pathogenicity of Rhizoctonia spp. isolated from winter squash in the Black Sea region of Turkey. Eur. J. Plant Pathol. 2016, 146, 683–697. [Google Scholar] [CrossRef]
- Özer, G.; Bayraktar, H. Characterization and sensitivity to fungicides of Rhizoctonia spp. recovered from potato plants in Bolu, Turkey. J. Phytopathol. 2015, 163, 11–18. [Google Scholar] [CrossRef]
- Erper, I.; Yildirim, E. Characterization and pathogenicity of Rhizoctonia spp. isolated from vegetable crops grown in greenhouses in Samsun province, Turkey. Biosci. J. 2017, 33. [Google Scholar] [CrossRef] [Green Version]
- Ogoshi, A. Grouping of Rhizoctonia solani Kühn and their perfect stages. Bull. Nat. Inst. Agric. Sci. Ser. C 1975, 30, 1–63. [Google Scholar]
- Bandoni, R.J. Safranin o as a rapid nuclear stain for fungi. Mycologia 2018, 71, 873–874. [Google Scholar] [CrossRef]
- Kalendar, R.; Boronnikova, S.; Seppanen, M. Isolation and purification of DNA from complicated biological samples. Methods Mol. Biol. 2021, 2222, 57–67. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Clustal W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Roldán-Ruiz, I.; Dendauw, J.; Van Bockstaele, E.; Depicker, A.; De Loose, M. AFLP markers reveal high polymorphic rates in ryegrasses (Lolium spp.). Mol. Breed. 2000, 6, 125–134. [Google Scholar] [CrossRef]
- Prevost, A.; Wilkinson, M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]
- Rohlf, F.J. Ntsys-Pc: Numerical Taxonomy and Multivariate Analysis System Version 2.1; Exeter Publishing Setauket: New York, NY, USA, 2000. [Google Scholar]
- R-Team. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 March 2021).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Earl, D.A.; vonHoldt, B.M. Structure harvester: A website and program for visualizing structure output and implementing the evanno method. Conserv. Genet. Resour. 2011, 4, 359–361. [Google Scholar] [CrossRef]
- Brayford, D. Methods for Research on Soilborne Phytopathogenic Fungi; Singleton, L.L., Mihail, J.D., Rush, C.M., Eds.; APS Press: St. Paul, MN, USA, 1993; pp. 103–106. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Rollins, P.A.; Keinath, A.P.; Farnham, M.W. Effect of inoculum type and anastomosis group of Rhizoctonia solani causing wirestem of cabbage seedlings in a controlled environment. Can. J. Plant Pathol. 1999, 21, 119–124. [Google Scholar] [CrossRef]
- Yitbarek, S.M.; Verma, P.R.; Morrall, R.A.A. Anastomosis groups, pathogenicity, and specificity of Rhizoctonia solani isolates from seedling and adult rapeseed/canola plants and soils in Saskatchewan. Can. J. Plant Pathol. 1987, 9, 6–13. [Google Scholar] [CrossRef]
- Verma, P.R. Biology and control of Rhizoctonia solani on rapeseed: A review. Phytoprotection 2005, 77, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Balali, G.R.; Neate, S.M.; Scott, E.S.; Whisson, D.L.; Wicks, T.J. Anastomosis group and pathogenicity of isolates of Rhizoctonia solani from potato crops in South Australia. Plant Pathol. 1995, 44, 1050–1057. [Google Scholar] [CrossRef]
- Carling, D.E. Effect of temperature on virulence of Rhizoctonia solani and other Rhizoctonia on potato. Phytopathology 1990, 80, 930–934. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Yang, X.B. Recovery of anastomosis groups of Rhizoctonia solani from different latitudinal positions and influence of temperatures on their growth and survival. Plant Dis. 2004, 88, 817–823. [Google Scholar] [CrossRef] [Green Version]
- Yitbarek, S.M.; Verma, P.R.; Gugel, R.K.; Morrall, R.A.A. Effect of soil temperature and inoculum density on pre-emergence damping-off of canola caused by Rhizoctonia solani. Can. J. Plant Pathol. 1988, 10, 93–98. [Google Scholar] [CrossRef]
- Erper, I.; Hatat Karaca, G.; Özkoc, I. Characterization of Rhizoctonia species causing root-rot of cucumber plants grown in greenhouses in Samsun, Turkey. Acta Hortic. 2002, 531–534. [Google Scholar] [CrossRef]
- Erper, I.; Ozkoc, I.; Karaca, G.H. Identification and pathogenicity of Rhizoctonia species isolated from bean and soybean plants in Samsun, Turkey. Arch. Phytopathol. Plant Prot. 2011, 44, 78–84. [Google Scholar] [CrossRef]
- Keinath, A.P. Relationships between inoculum density of Rhizoctonia solani, wirestem incidence and severity, and growth of cabbage. Phytopathology 1995, 85, 1487–1492. [Google Scholar] [CrossRef]
- Cubeta, M.A.; Vilgalys, R. Population biology of the Rhizoctonia solani complex. Phytopathology 1997, 87, 480–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharon, M.; Sneh, B.; Kuninaga, S.; Hyakumachi, M. The advancing identification and classification of Rhizoctonia spp. using molecular and biotechnological methods compared with the classical anastomosis grouping. Mycoscience 2006, 47, 299–316. [Google Scholar] [CrossRef]
- Ozer, G.; Imren, M.; Alkan, M.; Paulitz, T.C.; Bayraktar, H.; Palacioğlu, G.; Mehdiyev, I.; Muminjanov, H.; Dababat, A.A. Molecular and pathogenic characterization of Cochliobolus anamorphs associated with common root rot of wheat in Azerbaijan. Phytopathol. Mediterr. 2020, 59, 147–158. [Google Scholar]
- Šķipars, V.; Siaredzich, M.; Belevich, V.; Bruņeviča, N.; Brūna, L.; Ruņģis, D.E. Genetic differentiation of Phoma sp. isolates using retrotransposon-based iPBS assays. Environ. Exp. Biol. 2018, 16, 307–314. [Google Scholar] [CrossRef]
- Ogoshi, A. Introduction—The genus Rhizoctonia. In Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control; Springer: Dordrecht, The Netherlands, 1996; pp. 1–9. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Primer Sequences (5′–3′) | Ta (°C) | GC (%) | TB | PB | PIC | RP |
|---|---|---|---|---|---|---|---|
| 2078 | GCGGAGTCGCCA | 62.8 | 75.0 | 17 | 15 | 0.16 | 3.26 |
| 2095 | GCTCGGATACCA | 53.7 | 58.3 | 13 | 12 | 0.18 | 2.82 |
| 2219 | GAACTTATGCCGATACCA | 53.0 | 44.4 | 11 | 9 | 0.24 | 3.41 |
| 2222 | ACTTGGATGCCGATACCA | 53.0 | 50.0 | 12 | 10 | 0.22 | 3.26 |
| 2237 | CCCCTACCTGGCGTGCCA | 55.0 | 72.2 | 11 | 10 | 0.16 | 2.00 |
| 2239 | ACCTAGGCTCGGATGCCA | 55.0 | 61.1 | 12 | 11 | 0.14 | 1.94 |
| 2395 | TCCCCAGCGGAGTCGCCA | 52.8 | 72.2 | 30 | 29 | 0.15 | 5.32 |
| Total | 106 | 96 | |||||
| Avg. | 15.14 | 13.71 | 0.18 | 3.15 |
| Anastomosis Group | Isolate Denomination * | Location | GenBank Accession Nos. |
|---|---|---|---|
| AG 4 HG-I | Rs-RC-1 | Ağıllar | MT068296 |
| Rs-RC-2 | Ağıllar | MT068297 | |
| Rs-RC-3 | Ağıllar | MT068298 | |
| Rs-RC-4 | Ağıllar | MT068299 | |
| Rs-RC-5 | Ağıllar | MT068300 | |
| Rs-RC-6 | Ağıllar | MT068301 | |
| Rs-RC-7 | Altınova | MT068302 | |
| Rs-RC-8 | Altınova | MT068303 | |
| Rs-RC-9 | Altınova | MT068304 | |
| Rs-RC-10 | Altınova | MT068305 | |
| Rs-RC-11 | Altınova | MT068306 | |
| Rs-RC-12 | Altınova | MT068307 | |
| Rs-RC-13 | Altınova | MT068308 | |
| Rs-RC-14 | Altınova | MT068309 | |
| Rs-RC-15 | Balıklar | MT068310 | |
| Rs-RC-16 | Balıklar | MT068311 | |
| Rs-RC-17 | Çetinkaya | MT068312 | |
| Rs-RC-18 | Çetinkaya | MT068313 | |
| Rs-RC-19 | Dedeli | MT068314 | |
| Rs-RC-20 | Dereköy | MT068315 | |
| Rs-RC-21 | Dereköy | MT068316 | |
| Rs-RC-22 | Doğanca | MT068317 | |
| Rs-RC-23 | Doğanca | MT068318 | |
| Rs-RC-24 | Doğanca | MT068319 | |
| Rs-RC-25 | Doğanca | MT068320 | |
| Rs-RC-26 | Karpuzlu | MT068321 | |
| Rs-RC-27 | Karpuzlu | MT068322 | |
| Rs-RC-28 | Karpuzlu | MT068323 | |
| Rs-RC-29 | Karpuzlu | MT068324 | |
| Rs-RC-30 | Karpuzlu | MT068325 | |
| Rs-RC-31 | Karpuzlu | MT068326 | |
| Rs-RC-32 | Karpuzlu | MT068327 | |
| Rs-RC-33 | Karpuzlu | MT068328 | |
| Rs-RC-34 | Karpuzlu | MT068329 | |
| Rs-RC-35 | Karpuzlu | MT068330 | |
| Rs-RC-36 | Karpuzlu | MT068331 | |
| Rs-RC-37 | Karpuzlu | MT068332 | |
| Rs-RC-38 | Karpuzlu | MT068333 | |
| Rs-RC-39 | Karpuzlu | MT068334 | |
| Rs-RC-40 | Karpuzlu | MT068335 | |
| Rs-RC-41 | Karpuzlu | MT068336 | |
| Rs-RC-42 | Kaygusuz | MT068337 | |
| Rs-RC-43 | Kaygusuz | MT068338 | |
| Rs-RC-44 | Kaygusuz | MT068339 | |
| Rs-RC-45 | Kaygusuz | MT068340 | |
| Rs-RC-46 | Kaygusuz | MT068341 | |
| Rs-RC-47 | Kuşçular | MT068342 | |
| Rs-RC-48 | Kuşçular | MT068343 | |
| Rs-RC-49 | Osmanbeyli | MT068344 | |
| Rs-RC-50 | Osmanbeyli | MT068345 | |
| Rs-RC-51 | Osmanbeyli | MT068346 | |
| Rs-RC-52 | Osmanbeyli | MT068347 | |
| Rs-RC-53 | Osmanbeyli | MT068348 | |
| Rs-RC-54 | Osmanbeyli | MT068349 | |
| Rs-RC-55 | Osmanbeyli | MT068350 | |
| Rs-RC-56 | Osmanbeyli | MT068351 | |
| Rs-RC-57 | Üçpınar | MT068352 | |
| Rs-RC-58 | Osmanbeyli | MT068353 | |
| Rs-RC-59 | Osmanbeyli | MT068354 | |
| Rs-RC-60 | Osmanbeyli | MT068355 | |
| Rs-RC-61 | Osmanbeyli | MT068356 | |
| Rs-RC-62 | Osmanbeyli | MT068357 | |
| Rs-RC-63 | Osmanbeyli | MT068358 | |
| Rs-RC-64 | Osmanbeyli | MT068359 | |
| Rs-RC-65 | Osmanbeyli | MT068360 | |
| Rs-RC-66 | Osmanbeyli | MT068361 | |
| Rs-RC-67 | Osmanbeyli | MT068362 | |
| Rs-RC-68 | Şeyhören | MT068363 | |
| Rs-RC-69 | Şeyhören | MT068364 | |
| Rs-RC-70 | Şeyhören | MT068365 | |
| Rs-RC-71 | Şeyhören | MT068366 | |
| Rs-RC-72 | Türbe | MT068367 | |
| Rs-RC-73 | Türbe | MT068368 | |
| Rs-RC-74 | Türbe | MT068369 | |
| Rs-RC-75 | Türbe | MT068370 | |
| Rs-RC-76 | Türbe | MT068371 | |
| Rs-RC-77 | Türbe | MT068372 | |
| Rs-RC-78 | Türbe | MT068373 | |
| Rs-RC-79 | Türbe | MT068374 | |
| Rs-RC-80 | Türbe | MT068375 | |
| Rs-RC-81 | Türbe | MT068376 | |
| Rs-RC-82 | Türbe | MT068377 | |
| Rs-RC-83 | Türbe | MT068378 | |
| Rs-RC-84 | Türbe | MT068379 | |
| Rs-RC-85 | Türbe | MT068380 | |
| Rs-RC-86 | Türbe | MT068381 | |
| Rs-RC-87 | Türbe | MT068382 | |
| Rs-RC-88 | Türbe | MT068383 | |
| Rs-RC-89 | Türbe | MT068384 | |
| Rs-RC-90 | Üçpınar | MT068385 | |
| Rs-RC-91 | Üçpınar | MT068386 | |
| Rs-RC-92 | Üçpınar | MT068387 | |
| Rs-RC-93 | Üçpınar | MT068388 | |
| Rs-RC-94 | Üçpınar | MT068389 | |
| Rs-RC-95 | Üçpınar | MT068390 | |
| Rs-RC-96 | Üçpınar | MT068391 | |
| Rs-RC-97 | Üçpınar | MT068392 | |
| Rs-RC-98 | Üçpınar | MT068393 | |
| Rs-RC-99 | Üçpınar | MT068394 | |
| Rs-RC-100 | Üçpınar | MT068395 | |
| Rs-RC-101 | Yeşilyazı | MT068396 | |
| Rs-RC-102 | Yeşilyazı | MT068397 | |
| Rs-RC-103 | Yeşilyazı | MT068398 | |
| Rs-RC-104 | Yeşilyazı | MT068399 | |
| Rs-RC-105 | Yeşilyazı | MT068400 | |
| Rs-RC-106 | Yeşilyazı | MT068401 | |
| Rs-RC-107 | Yeşilyazı | MT068402 | |
| AG 4 HG-III | Rs-RC-108 | Koşuköy | MT068403 |
| Rs-RC-109 | Türbe | MT068404 | |
| Rs-RC-110 | Türbe | MT068405 | |
| Rs-RC-111 | Altınova | MT068406 | |
| AG 5 | Rs-RC-112 | Koşuköy | MT068407 |
| Rs-RC-113 | Koşuköy | MT068408 | |
| Rs-RC-114 | Koşuköy | MT068409 | |
| Rs-RC-115 | Balıklar | MT068410 | |
| Rs-RC-116 | Balıklar | MT068411 | |
| Rs-RC-117 | Balıklar | MT068412 | |
| Rs-RC-118 | Balıklar | MT068413 | |
| Rs-RC-119 | Balıklar | MT068414 | |
| Rs-RC-120 | Ağıllar | MT068415 | |
| Rs-RC-121 | Ağıllar | MT068416 | |
| Rs-RC-122 | Ağıllar | MT068417 | |
| Rs-RC-123 | Ağıllar | MT068418 | |
| Rs-RC-124 | Ağıllar | MT068419 | |
| AG-A | R-RC-125 | Balıklar | MT053135 |
| R-RC-126 | Koşuköy | MT053136 | |
| R-RC-127 | Koşuköy | MT053137 | |
| R-RC-128 | Türbe | MT053138 | |
| R-RC-129 | Türbe | MT053139 | |
| R-RC-130 | Türbe | MT053140 | |
| R-RC-131 | Türbe | MT053141 | |
| R-RC-132 | Türbe | MT053142 |
| Anastomosis Group | Isolate a | Root | Shoot Dry Weight (g) | Plant Height (cm) | |
|---|---|---|---|---|---|
| Dry Weight (g) | Length (cm) | ||||
| AG 4 HG-I | Rs-RC-04 | 0.013 b ± 0.002 c cde d | 2.083 ± 0.523 h | 0.127 ± 0.014 h | 4.883 ± 0.190 ijk |
| Rs-RC-16 | 0.014 ± 0.004 cde | 3.083 ± 0.688 c–h | 0.200 ± 0.016 def | 4.700 ± 0.191 k | |
| Rs-RC-18 | 0.010 ± 0.005 e | 2.000 ± 0.129 h | 0.148 ± 0.032 gh | 4.833 ± 0.211 jk | |
| Rs-RC-19 | 0.013 ± 0.007 cde | 2.333 ± 0.247 gh | 0.182 ± 0.020 e-h | 5.000 ± 0.289 h–k | |
| Rs-RC-20 | 0.017 ± 0.007 cde | 2.583 ± 0.300 e–h | 0.185 ± 0.020 efg | 5.333 ± 0.167 e–k | |
| Rs-RC-26 | 0.020 ± 0.012 cde | 2.717 ± 0.415 d–h | 0.232 ± 0.023 b–e | 5.167 ± 0.333 f–k | |
| Rs-RC-28 | 0.021 ± 0.005 cde | 3.167 ± 0.105 b–h | 0.212 ± 0.017 def | 5.250 ± 0.214 f–k | |
| Rs-RC-41 | 0.019 ± 0.005 cde | 3.367 ± 0.851 b–h | 0.202 ± 0.016 def | 5.083 ± 0.539 g–k | |
| Rs-RC-42 | 0.019 ± 0.007 cde | 3.500 ± 0.129 b-g | 0.195 ± 0.023 d–g | 5.667 ± 0.279 c–i | |
| Rs-RC-47 | 0.013 ± 0.004 cde | 3.083 ± 0.455 b–h | 0.243 ± 0.010 b–e | 5.833 ± 0.307 b–f | |
| Rs-RC-69 | 0.010 ± 0.002 e | 2.417 ± 0.375 fgh | 0.212 ± 0.017 def | 5.417 ± 0.300 d–k | |
| Rs-RC-76 | 0.029 ± 0.005 bc | 3.583 ± 0.271 b–g | 0.237 ± 0.029 b–e | 5.750 ± 0.423 c–h | |
| Rs-RC-82 | 0.023 ± 0.008 b | 3.750 ± 0.382 b–f | 0.220 ± 0.015 c–f | 5.917 ± 0.271 b–f | |
| Rs-RC-105 | 0.013 ± 0.005 cde | 3.000 ± 0.258 c–h | 0.178 ± 0.033 fgh | 4.950 ± 0.263 h–k | |
| AG 4 HG-III | Rs-RC-108 | 0.025 ± 0.009 bcd | 3.833 ± 0.211 a–e | 0.218 ± 0.010 c-f | 5.667 ± 0.401 c–i |
| Rs-RC-109 | 0.026 ± 0.002 bcd | 4.083 ± 0.490 a–d | 0.198 ± 0.022 def | 5.550 ± 0.293 c–j | |
| AG 5 | Rs-RC-114 | 0.027 ± 0.009 bcd | 3.917 ± 0.712 a–e | 0.205 ± 0.020 def | 5.917 ± 0.300 b–f |
| Rs-RC-118 | 0.018 ± 0.005 cde | 3.833 ± 0.833 b–f | 0.263 ± 0.023 bcd | 5.800 ± 0.163 b–g | |
| Rs-RC-122 | 0.018 ± 0.007 cde | 3.167 ± 0.558 b–h | 0.257 ± 0.021 bcd | 6.333 ± 0.247 abc | |
| Rs-RC-123 | 0.026 ± 0.013 bcd | 3.950 ± 0.419 a–e | 0.262 ± 0.013 bcd | 6.667 ± 0.401 ab | |
| AG-A | R-RC-125 | 0.029 ± 0.006 bc | 4.000 ± 0.606 a–e | 0.297 ± 0.042 bc | 6.200 ± 0.100 a–d |
| R-RC-127 | 0.057 ± 0.013 b | 4.500 ± 0.548 ab | 0.315 ± 0.041 b | 6.083 ± 0.239 a–e | |
| R-RC-129 | 0.046 ± 0.005 b | 4.167 ± 0.792 a–d | 0.292 ± 0.026 bc | 6.250 ± 0.310 a–d | |
| R-RC-131 | 0.027 ± 0.007 bcd | 4.250 ± 0.250 abc | 0.232 ± 0.031 cde | 6.183 ± 0.290 a–d | |
| Control | Control | 0.203 ± 0.018 a | 5.333 ± 0.558 a | 0.558 ± 0.049a | 6.917 ± 0.271 a |
| Isolate | Disease Severity Observed on Variety a | |||||
|---|---|---|---|---|---|---|
| Rescue | Travero | Integro | Remala | Redriver | Rondale | |
| AG 4 HG-I | 4.0 b ± 0.00 c b d | 4.0 ± 0.00 b | 4.0 ± 0.00 b | 4.0 ± 0.00 c | 4.0 ± 0.00 c | 4.0 ± 0.00 c |
| AG 4 HG-III | 4.0 ± 0.00 b B e | 4.0 ± 0.00 b B | 4.0 ± 0.00 b B | 3.3 ± 0.21 bc A | 3.5 ± 0.22 bc AB | 3.5 ± 0.22 bc AB |
| AG 5 | 4.0 ± 0.00 b B | 4.0 ± 0.00 b B | 4.0 ± 0.00 b B | 3.0 ± 0.00 b A | 2.5 ± 0.22 b A | 4.0 ± 0.00 c B |
| AG-A | 3.0 ± 0.26 a B | 3.0 ± 0.00 a B | 3.2 ± 0.31 b B | 2.7 ± 0.21 b AB | 2.3 ± 0.21 ab A | 3.2 ± 0.17 ab B |
| Control | 1.0 ± 0.00 a | 1.0 ± 0.00 a | 1.0 ± 0.00 a | 1.0 ± 0.00 a | 1.0 ± 0.00 a | 1.0 ± 0.00 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erper, I.; Ozer, G.; Kalendar, R.; Avci, S.; Yildirim, E.; Alkan, M.; Turkkan, M. Genetic Diversity and Pathogenicity of Rhizoctonia spp. Isolates Associated with Red Cabbage in Samsun (Turkey). J. Fungi 2021, 7, 234. https://doi.org/10.3390/jof7030234
Erper I, Ozer G, Kalendar R, Avci S, Yildirim E, Alkan M, Turkkan M. Genetic Diversity and Pathogenicity of Rhizoctonia spp. Isolates Associated with Red Cabbage in Samsun (Turkey). Journal of Fungi. 2021; 7(3):234. https://doi.org/10.3390/jof7030234
Chicago/Turabian StyleErper, Ismail, Goksel Ozer, Ruslan Kalendar, Sirin Avci, Elif Yildirim, Mehtap Alkan, and Muharrem Turkkan. 2021. "Genetic Diversity and Pathogenicity of Rhizoctonia spp. Isolates Associated with Red Cabbage in Samsun (Turkey)" Journal of Fungi 7, no. 3: 234. https://doi.org/10.3390/jof7030234