
Complete Utilization of the Major Carbon Sources Present in Sugar Beet Pulp Hydrolysates by the Oleaginous Red Yeasts Rhodotorula toruloides and R. mucilaginosa

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Rhodotorula mucilaginosa IST 390 from Sugar Beet Pulp
2.2. Yeast Strains, Growth Media and Conditions
2.3. Effect of Acetic Acid Concentration on Yeast Growth and Carbon Source Catabolism
2.4. Preparation of Sugar Beet Pulp Hydrolysate
2.5. Determination of Concentrations of Sugars and Acetic Acid
2.6. Assessment of Lipid Production
2.7. Assessment of Carotenoid Production
2.8. Statistical Analysis
3. Results
3.1. Isolation of Rhodotorula mucilaginosa IST 390 from Sugar Beet Pulp
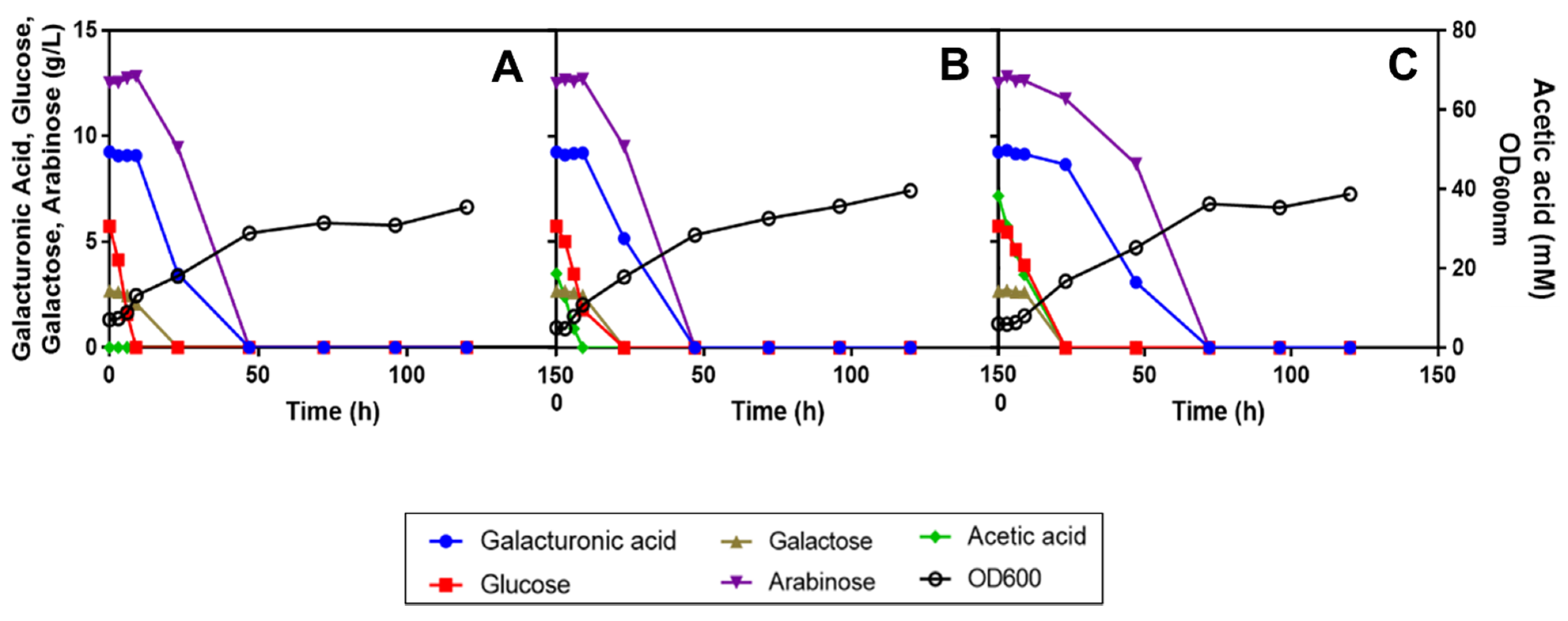
3.2. Differential Utilization of the Carbon Sources Present in SBP Hydrolysate during R. mucilaginosa IST 390 and R. toruloides PYCC 5615 Cultivation
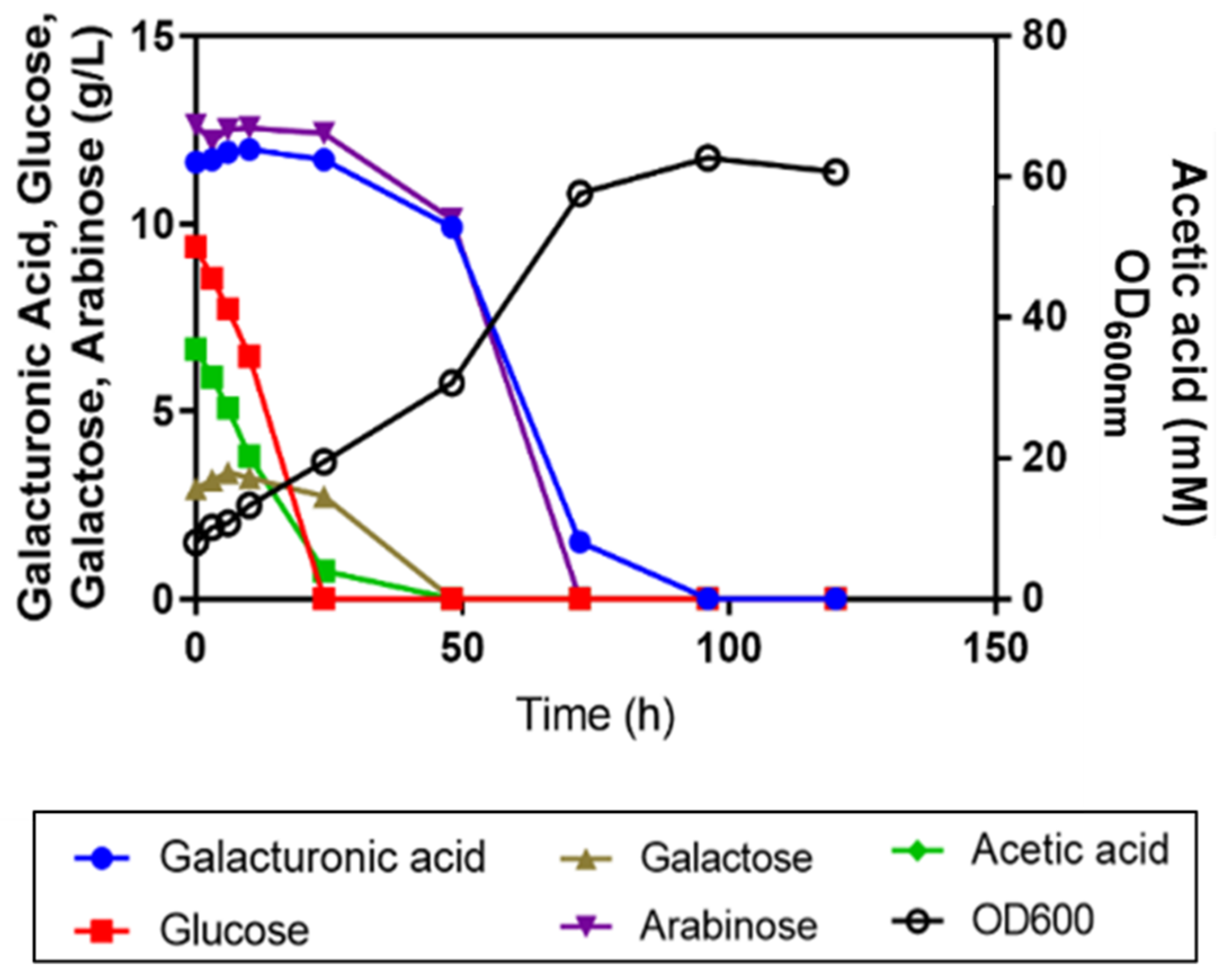
3.3. Sugar Utilization from a Synthetic SBP Hydrolysate Medium Is Affected by Acetic Acid and Improved by Amino Acid Supplementation and Inoculum Increase
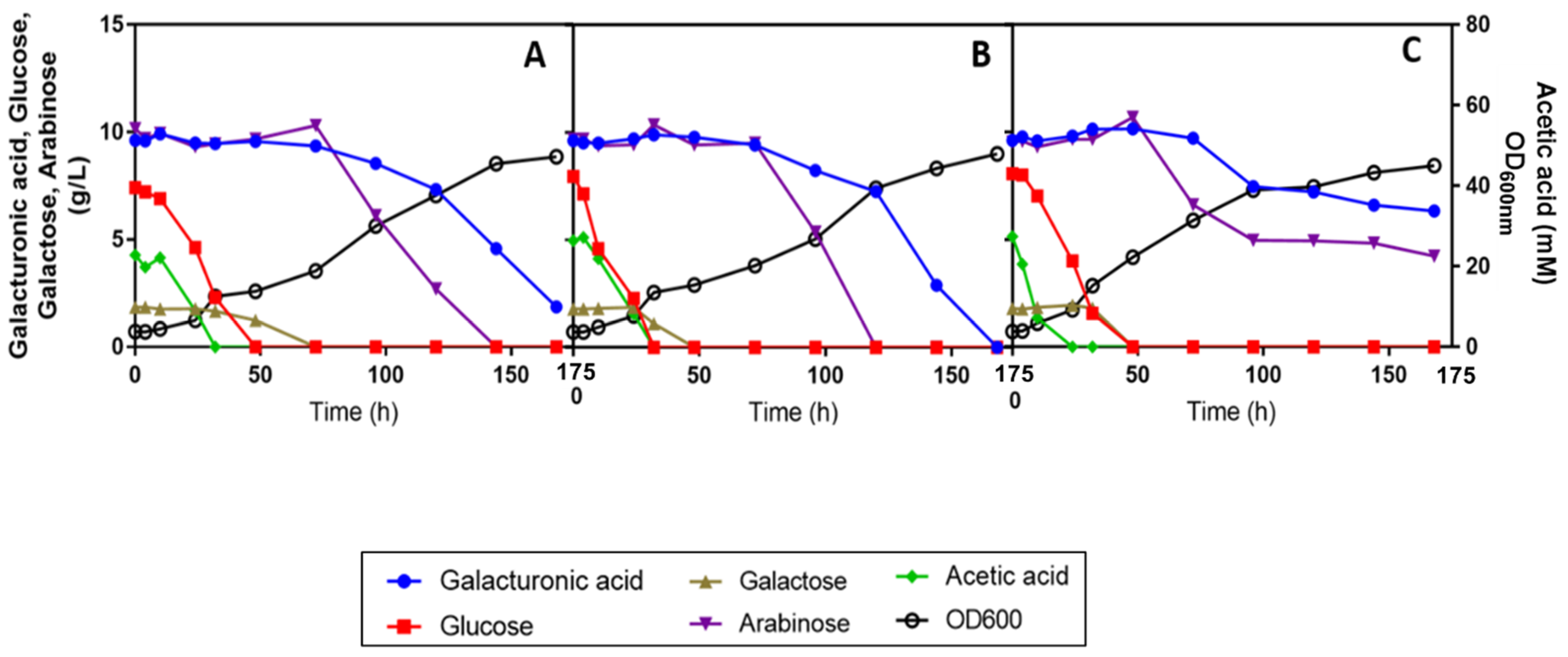
3.4. Performance of R. Toruloides PYCC 5615 in SBP Hydrolysate Supplemented with Commercial YNB with Amino Acids and Effect of Ammonium Sulphate Concentration
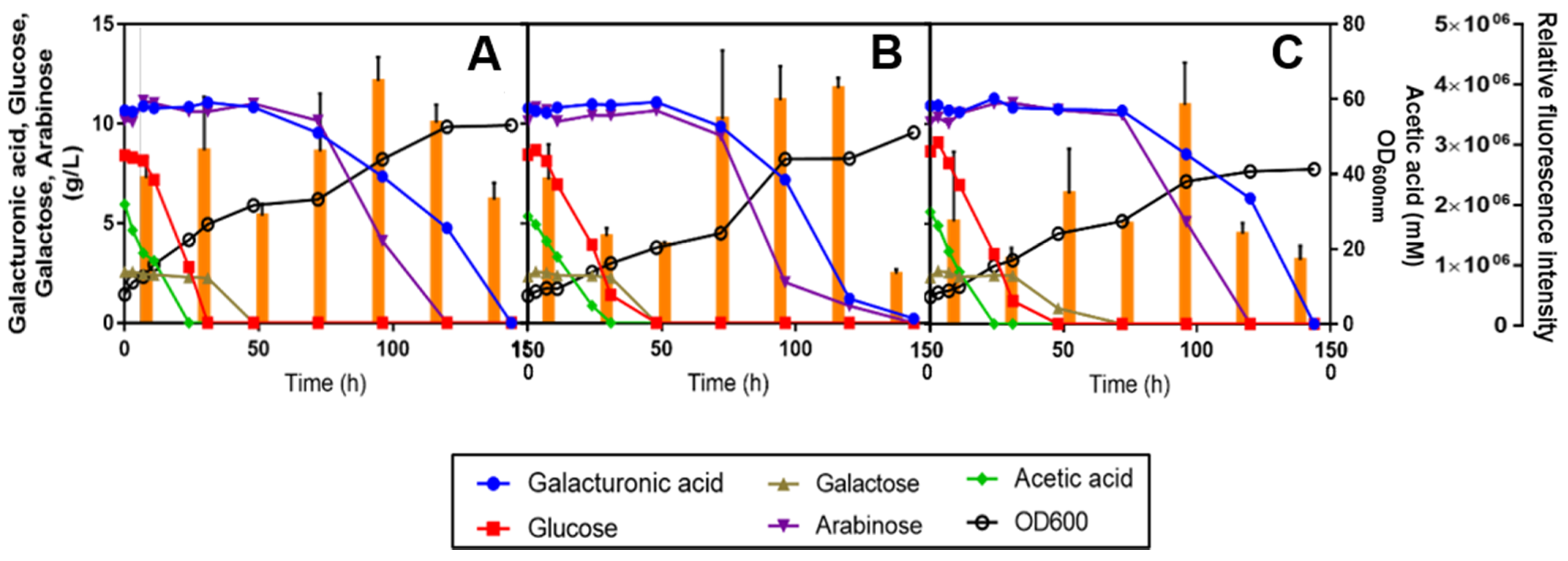
3.5. Preliminary Assessment of Lipid and Carotenoid Production by Three Oleaginous Red Yeast Strains during SBP Bioconversion
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuin, V.G.; Segatto, M.L.; Ramin, L.Z. Plants as resources for organic molecules: Facing the green and sustainable future today. Curr. Opin. Green Sustain. Chem. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Rebello, S.; Abraham, A.; Madhavan, A.; Sindhu, R.; Binod, P.; Babu, A.K.; Aneesh, E.M.; Pandey, A. Non-conventional Yeast cell factories for sustainable bioprocesses. FEMS Microbiol. Lett. 2018, 365, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.C.; Monteiro, C.C.; Semedo, P.M.; Sá-correia, I. Valorisation of pectin-rich agro-industrial residues by yeasts: Potential and challenges. Appl. Microbiol. Biotechnol. 2020, 104, 6527–6547. [Google Scholar] [CrossRef]
- Cherubini, F. The biorefinery concept: Using biomass instead of oil for producing energy and chemicals. Energy Convers. Manag. 2010, 51, 1412–1421. [Google Scholar] [CrossRef]
- Dahiya, S.; Kumar, A.N.; Shanthi Sravan, J.; Chatterjee, S.; Sarkar, O.; Mohan, S.V. Food waste biorefinery: Sustainable strategy for circular bioeconomy. Bioresour. Technol. 2018, 248, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Martani, F.; Maestroni, L.; Torchio, M.; Ami, D.; Natalello, A.; Lotti, M.; Porro, D.; Branduardi, P. Conversion of sugar beet residues into lipids by Lipomyces starkeyi for biodiesel production. Microb. Cell Fact. 2020, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Peters, D. Raw materials. Adv. Biochem. Eng. Biotechnol. 2006, 105, 1–30. [Google Scholar]
- Berlowska, J.; Binczarski, M.; Dziugan, P.; Wilkowska, A.; Kregiel, D.; Witonska, I. Sugar Beet Pulp as a Source of Valuable Biotechnological Products. In Advances in Biotechnology for Food Industry; Elsevier: Amsterdam, The Netherlands, 2018; Volume 14, pp. 359–392. ISBN 9780128114957. [Google Scholar]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Leijdekkers, A.G.M.; Bink, J.P.M.; Geutjes, S.; Schols, H.A.; Gruppen, H. Enzymatic saccharification of sugar beet pulp for the production of galacturonic acid and arabinose; a study on the impact of the formation of recalcitrant oligosaccharides. Bioresour. Technol. 2013, 128, 518–525. [Google Scholar] [CrossRef]
- Hamley-Bennett, C.; Lye, G.J.; Leak, D.J. Selective fractionation of Sugar Beet Pulp for release of fermentation and chemical feedstocks; optimisation of thermo-chemical pre-treatment. Bioresour. Technol. 2016, 209, 259–264. [Google Scholar] [CrossRef] [Green Version]
- Berlowska, J.; Pielech-Przybylska, K.; Balcerek, M.; Cieciura, W.; Borowski, S.; Kregiel, D. Integrated bioethanol fermentation/anaerobic digestion for valorization of sugar beet pulp. Energies 2017, 10, 1255. [Google Scholar] [CrossRef] [Green Version]
- Sakai, T.; Sakamoto, T.; Hallaert, J.; Vandamme, E.J. Pectin, Pectinase, and Protopectinase: Production, Properties, and Applications. Adv. Appl. Microbiol. 1993, 39, 213–294. [Google Scholar] [CrossRef]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Teixeira, M.C.; Mira, N.P.; Sá-Correia, I. A genome-wide perspective on the response and tolerance to food-relevant stresses in Saccharomyces cerevisiae. Curr. Opin. Biotechnol. 2011, 22, 150–156. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, S.C.; Sá-Correia, I. Yeast toxicogenomics: Lessons from a eukaryotic cell model and cell factory. Curr. Opin. Biotechnol. 2015, 33, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Satyanarayana, T.; Kunze, G. Yeast Biotechnology: Diversity and Applications; Springer: Dordrecht, The Netherlands, 2009; ISBN 978-1-4020-8291-7. [Google Scholar]
- Nielsen, J. Yeast Systems Biology: Model Organism and Cell Factory. Biotechnol. J. 2019, 14, 1800421. [Google Scholar] [CrossRef] [Green Version]
- Radecka, D.; Mukherjee, V.; Mateo, R.Q.; Stojiljkovic, M.; Foulquié-Moreno, M.R.; Thevelein, J.M. Looking beyond Saccharomyces: The potential of non-conventional yeast species for desirable traits in bioethanol fermentation. FEMS Yeast Res. 2015, 15, 53. [Google Scholar] [CrossRef] [Green Version]
- Diaz, A.B.; Blandino, A.; Caro, I. Value added products from fermentation of sugars derived from agro-food residues. Trends Food Sci. Technol. 2018, 71, 52–64. [Google Scholar] [CrossRef]
- Micard, V.; Renard, C.M.G.C.; Thibault, J.-F. Enzymatic saccharification of sugar-beet pulp. Enzym. Microb. Technol. 1996, 19, 162–170. [Google Scholar] [CrossRef]
- Kayikci, Ö.; Nielsen, J. Glucose repression in Saccharomyces cerevisiae. FEMS Yeast Res. 2015, 15, fov068. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Shen, X.; Yuan, Q.; Yan, Y. Metabolic Engineering Strategies for Co-Utilization of Carbon Sources in Microbes. Bioengineering 2016, 3, 10. [Google Scholar] [CrossRef]
- Huisjes, E.H.; de Hulster, E.; van Dam, J.C.; Pronk, J.T.; van Maris, A.J.A. Galacturonic Acid Inhibits the Growth of Saccharomyces cerevisiae on Galactose, Xylose, and Arabinose. Appl. Environ. Microbiol. 2012, 78, 5052–5059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, T.; Hamada, S.; Wakabayashi, A.; Kishida, M. Fermentative production of l-galactonate by using recombinant Saccharomyces cerevisiae containing the endogenous galacturonate reductase gene from Cryptococcus diffluens. J. Biosci. Bioeng. 2016, 122, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Benz, J.; Protzko, R.J.; Andrich, J.M.; Bauer, S.; Dueber, J.E.; Somerville, C.R. Identification and characterization of a galacturonic acid transporter from Neurospora crassa and its application for Saccharomyces cerevisiae fermentation processes. Biotechnol. Biofuels 2014, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, D.; Ye, S.; Park, H.; Kim, S.R. Simultaneous fermentation of galacturonic acid and five-carbon sugars by engineered Saccharomyces cerevisiae. Bioresour. Technol. 2020, 295, 122259. [Google Scholar] [CrossRef] [PubMed]
- Protzko, R.J.; Latimer, L.N.; Martinho, Z.; de Reus, E.; Seibert, T.; Benz, J.P.; Dueber, J.E. Engineering Saccharomyces cerevisiae for co-utilization of d-galacturonic acid and d-glucose from citrus peel waste. Nat. Commun. 2018, 9, 5059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biz, A.; Sugai-Guérios, M.H.; Kuivanen, J.; Maaheimo, H.; Krieger, N.; Mitchell, D.A.; Richard, P. The introduction of the fungal d-galacturonate pathway enables the consumption of d-galacturonic acid by Saccharomyces cerevisiae. Microb. Cell Fact. 2016, 15, 144. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.; Jeong, D.; Shon, J.C.; Liu, K.H.; Kim, K.H.; Shin, M.; Kim, S.R. Deletion of PHO13 improves aerobic l-arabinose fermentation in engineered Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2019, 46, 1725–1731. [Google Scholar] [CrossRef] [PubMed]
- Wisselink, H.W.; Toirkens, M.J.; del Berriel, M.F.R.; Winkler, A.A.; van Dijken, J.P.; Pronk, J.T.; van Maris, A.J.A. Engineering of Saccharomyces cerevisiae for Efficient Anaerobic Alcoholic Fermentation of L-arabinose. Appl. Environ. Microbiol. 2007, 73, 4881–4891. [Google Scholar] [CrossRef] [Green Version]
- Richard, P.; Hilditch, S. d-Galacturonic acid catabolism in microorganisms and its biotechnological relevance. Appl. Microbiol. Biotechnol. 2009, 82, 597–604. [Google Scholar] [CrossRef]
- Protzko, R.J.; Hach, C.A.; Coradetti, S.T.; Hackhofer, M.A.; Magosch, S.; Thieme, N.; Geiselman, G.M.; Arkin, A.P.; Skerker, J.M.; Dueber, J.E.; et al. Genomewide and Enzymatic Analysis Reveals Efficient d-galacturonic Acid Metabolism in the Basidiomycete Yeast Rhodosporidium toruloides. mSystems 2019, 4, e00389-19. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.-K.; Nicaud, J.-M.; Ledesma-Amaro, R. The Engineering Potential of Rhodosporidium toruloides as a Workhorse for Biotechnological Applications. Trends Biotechnol. 2018, 36, 304–317. [Google Scholar] [CrossRef]
- Cárdenas-Fernández, M.; Bawn, M.; Hamley-Bennett, C.; Bharat, P.K.V.; Subrizi, F.; Suhaili, N.; Ward, D.P.; Bourdin, S.; Dalby, P.A.; Hailes, H.C.; et al. An integrated biorefinery concept for conversion of sugar beet pulp into value-added chemicals and pharmaceutical intermediates. Faraday Discuss. 2017, 202, 415–431. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.D.; Boundy-Mills, K.L.; Barla, G.F.; Kumar, S.; Ubanwa, B.; Balan, V. Microbial Lipid Alternatives to Plant Lipids; Humana: New York, NY, USA, 2019; Volume 1995, ISBN 9781493994847. [Google Scholar]
- Kot, A.M.; Błazejak, S.; Gientka, I.; Kieliszek, M.; Bryś, J. Torulene and torularhodin: “New” fungal carotenoids for industry? Microb. Cell Fact. 2018, 17, 49. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, C.S. Preparation of Yeast DNA. Curr. Protoc. Mol. Biol. 1997, 39, 11–14. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek. Int. J. Gen. Mol. Microbiol. 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Banno, I. Studies on the sexuality of Rhodotorula. J. Gen. Appl. Microbiol. 1967, 13, 167–196. [Google Scholar] [CrossRef]
- Dinh, H.V.; Suthers, P.F.; Chan, S.H.J.; Shen, Y.; Xiao, T.; Deewan, A.; Jagtap, S.S.; Zhao, H.; Rao, C.V.; Rabinowitz, J.D.; et al. A comprehensive genome-scale model for Rhodosporidium toruloides IFO0880 accounting for functional genomics and phenotypic data. Metab. Eng. Commun. 2019, 9, e00101. [Google Scholar] [CrossRef]
- Kumar, K.K.; Deeba, F.; Negi, Y.S.; Gaur, N.A. Harnessing pongamia shell hydrolysate for triacylglycerol agglomeration by novel oleaginous yeast Rhodotorula pacifica INDKK. Biotechnol. Biofuels 2020, 13, 175. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Amaya, D.B.; Kimura, M. HarvestPlus Handbook for Carotenoid Analysis; HarvestPlus; International Food Policy Research Institute (IFPRI): Washington, DC, USA, 2004. [Google Scholar]
- Martens-Uzunova, E.S.; Schaap, P.J. An evolutionary conserved d-galacturonic acid metabolic pathway operates across filamentous fungi capable of pectin degradation. Fungal Genet. Biol. 2008, 45, 1449–1457. [Google Scholar] [CrossRef] [PubMed]
- Taskin, M.; Ortucu, S.; Aydogan, M.N.; Arslan, N.P. Lipid production from sugar beet molasses under non-aseptic culture conditions using the oleaginous yeast Rhodotorula glutinis TR29. Renew. Energy 2016, 99, 198–204. [Google Scholar] [CrossRef]
- Carota, E.; Petruccioli, M.; D’Annibale, A.; Gallo, A.M.; Crognale, S. Orange peel waste–based liquid medium for biodiesel production by oleaginous yeasts. Appl. Microbiol. Biotechnol. 2020, 104, 4617–4628. [Google Scholar] [CrossRef]
- Sellick, C.A.; Campbell, R.N.; Reece, R.J. Galactose Metabolism in Yeast—Structure and Regulation of the Leloir Pathway Enzymes and the Genes Encoding Them. In International Review of Cell and Molecular Biology; Faculty of Sciences Academic Press, University of Manchester: Manchester, UK, 2008; Volume 269, pp. 111–150. ISBN 9780123745545. [Google Scholar]
- Maza, D.D.; Viñarta, S.C.; Su, Y.; Guillamón, J.M.; Aybar, M.J. Growth and lipid production of Rhodotorula glutinis R4, in comparison to other oleaginous yeasts. J. Biotechnol. 2020, 310, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Lopes, H.J.S.; Bonturi, N.; Kerkhoven, E.J.; Miranda, E.A.; Lahtvee, P.J. C/N ratio and carbon source-dependent lipid production profiling in Rhodotorula toruloides. Appl. Microbiol. Biotechnol. 2020, 104, 2639–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, B.; Schmidt, C.; Daum, G. Storage lipids of yeasts: A survey of nonpolar lipid metabolism in Saccharomyces cerevisiae, Pichia pastoris, and Yarrowia lipolytica. FEMS Microbiol. Rev. 2014, 38, 892–915. [Google Scholar] [CrossRef] [Green Version]
- Van Zutphen, T.; Todde, V.; De Boer, R.; Kreim, M.; Hofbauer, H.F.; Wolinski, H.; Veenhuis, M.; Van Der Klei, I.J.; Kohlwein, S.D. Lipid droplet autophagy in the yeast Saccharomyces cerevisiae. Mol. Biol. Cell 2014, 25, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mata-Gómez, L.C.; Montañez, J.C.; Méndez-Zavala, A.; Aguilar, C.N. Biotechnological production of carotenoids by yeasts: An overview. Microb. Cell Fact. 2014, 13, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, K.D.; Shida, Y.; Miyata, A.; Takamizawa, T.; Suzuki, Y.; Ara, S.; Yamazaki, H.; Masaki, K.; Mori, K.; Aburatani, S.; et al. Effect of light on carotenoid and lipid production in the oleaginous yeast Rhodosporidium toruloides. Biosci. Biotechnol. Biochem. 2020, 84, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Shen, P.; Hu, R.; Xue, T.; Jiang, X.; Qin, L.; Chen, Y.; Huang, J. Carotenoids and lipid production from Rhodosporidium toruloides cultured in tea waste hydrolysate. Biotechnol. Biofuels 2020, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jeong, D.; Park, H.; Jang, B.-K.; Ju, Y.; Shin, M.H.; Oh, E.J.; Lee, E.J.; Kim, S.R. Recent advances in the biological valorization of citrus peel waste into fuels and chemicals. Bioresour. Technol. 2021, 323, 124603. [Google Scholar] [CrossRef]
- Müller-Maatsch, J.; Bencivenni, M.; Caligiani, A.; Tedeschi, T.; Bruggeman, G.; Bosch, M.; Petrusan, J.; Van Droogenbroeck, B.; Elst, K.; Sforza, S. Pectin content and composition from different food waste streams. Food Chem. 2016, 201, 37–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mota, M.N.; Martins, L.C.; Sá-Correia, I. The Identification of Genetic Determinants of Methanol Tolerance in Yeast Suggests Differences in Methanol and Ethanol Toxicity Mechanisms and Candidates for Improved Methanol Tolerance Engineering. J. Fungi 2021, 7, 90. [Google Scholar] [CrossRef] [PubMed]
- Sitepu, I.; Selby, T.; Lin, T.; Zhu, S.; Boundy-Mills, K. Carbon source utilization and inhibitor tolerance of 45 oleaginous yeast species. J. Ind. Microbiol. Biotechnol. 2014, 41, 1061–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, M.M.; Morrissey, J.P. Kluyveromyces marxianus: A yeast emerging from its sister’s shadow. Fungal Biol. Rev. 2010, 24, 17–26. [Google Scholar] [CrossRef]
- Palma, M.; Guerreiro, J.F.; Sá-Correia, I. Adaptive Response and Tolerance to Acetic Acid in Saccharomyces cerevisiae and Zygosaccharomyces bailii: A Physiological Genomics Perspective. Front. Microbiol. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Wamelink, M.M.C.; Struys, E.A.; Jakobs, C. The biochemistry, metabolism and inherited defects of the pentose phosphate pathway: A review. J. Inherit. Metab. Dis. 2008, 31, 703–717. [Google Scholar] [CrossRef]
- Fonseca, C.; Romão, R.; de Sousa, H.R.; Hahn-Hägerdal, B.; Spencer-Martins, I. Arabinose transport and catabolism in yeast. FEBS J. 2007, 274, 3589–3600. [Google Scholar] [CrossRef]
- Xu, P.; Bura, R.; Doty, S.L. Genetic analysis of D-xylose metabolism by endophytic yeast strains of Rhodotorula graminis and Rhodotorula mucilaginosa. Genet. Mol. Biol. 2011, 34, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Hu, J.; Fan, L.; Chen, Q. RNA-Seq-based transcriptomic and metabolomic analysis reveal stress responses and programmed cell death induced by acetic acid in Saccharomyces cerevisiae. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef]
- Orij, R.; Brul, S.; Smits, G.J. Intracellular pH is a tightly controlled signal in yeast. Biochim. Biophys. Acta Gen. Subj. 2011, 1810, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Croft, T.; Venkatakrishnan, P.; Lin, S.-J. NAD+ Metabolism and Regulation: Lessons from Yeast. Biomolecules 2020, 10, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, J. It Is All about Metabolic Fluxes. J. Bacteriol. 2003, 185, 7031–7035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, K.; Vowinckel, J.; Keller, M.A.; Ralser, M. Methionine Metabolism Alters Oxidative Stress Resistance via the Pentose Phosphate Pathway. Antioxid. Redox Signal. 2016, 24, 543–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratledge, C.; Wynn, J.P. The biochemistry and molecular biology of lipid accumulation in oleaginous microorganisms. Adv. Appl. Microbiol. 2002, 51, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Braunwald, T.; Schwemmlein, L.; Graeff-Hönninger, S.; French, W.T.; Hernandez, R.; Holmes, W.E.; Claupein, W. Effect of different C/N ratios on carotenoid and lipid production by Rhodotorula glutinis. Appl. Microbiol. Biotechnol. 2013, 97, 6581–6588. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme | d-Glucose g/L | d-Galactose g/L | l-Arabinose g/L | d-Galacturonic Acid g/L | Acetic Acid mM (g/L) |
|---|---|---|---|---|---|
| H11 | 9.4 | 4.8 | 13.1 | 11.3 | 33.5 (2.0) |
| H13 | 6.7 | 2.2 | 9.3 | 9.4 | 32.0 (1.9) |
| Yeast Strain | Total Carotenoids | |
|---|---|---|
| (μg/g Dry Biomass) | (mg/L) | |
| R. mucilaginosa IST 390 | 81 | 1.4 |
| R. toruloides PYCC 5615 | 255 | 5.4 |
| R. toruloides IFO 0880 | 158 | 2.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, L.C.; Palma, M.; Angelov, A.; Nevoigt, E.; Liebl, W.; Sá-Correia, I. Complete Utilization of the Major Carbon Sources Present in Sugar Beet Pulp Hydrolysates by the Oleaginous Red Yeasts Rhodotorula toruloides and R. mucilaginosa. J. Fungi 2021, 7, 215. https://doi.org/10.3390/jof7030215
Martins LC, Palma M, Angelov A, Nevoigt E, Liebl W, Sá-Correia I. Complete Utilization of the Major Carbon Sources Present in Sugar Beet Pulp Hydrolysates by the Oleaginous Red Yeasts Rhodotorula toruloides and R. mucilaginosa. Journal of Fungi. 2021; 7(3):215. https://doi.org/10.3390/jof7030215
Chicago/Turabian StyleMartins, Luís C., Margarida Palma, Angel Angelov, Elke Nevoigt, Wolfgang Liebl, and Isabel Sá-Correia. 2021. "Complete Utilization of the Major Carbon Sources Present in Sugar Beet Pulp Hydrolysates by the Oleaginous Red Yeasts Rhodotorula toruloides and R. mucilaginosa" Journal of Fungi 7, no. 3: 215. https://doi.org/10.3390/jof7030215