
Innate Resistance and Phosphite Treatment Affect Both the Pathogen’s and Host’s Transcriptomes in the Tanoak-Phytophthora ramorum Pathosystem

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
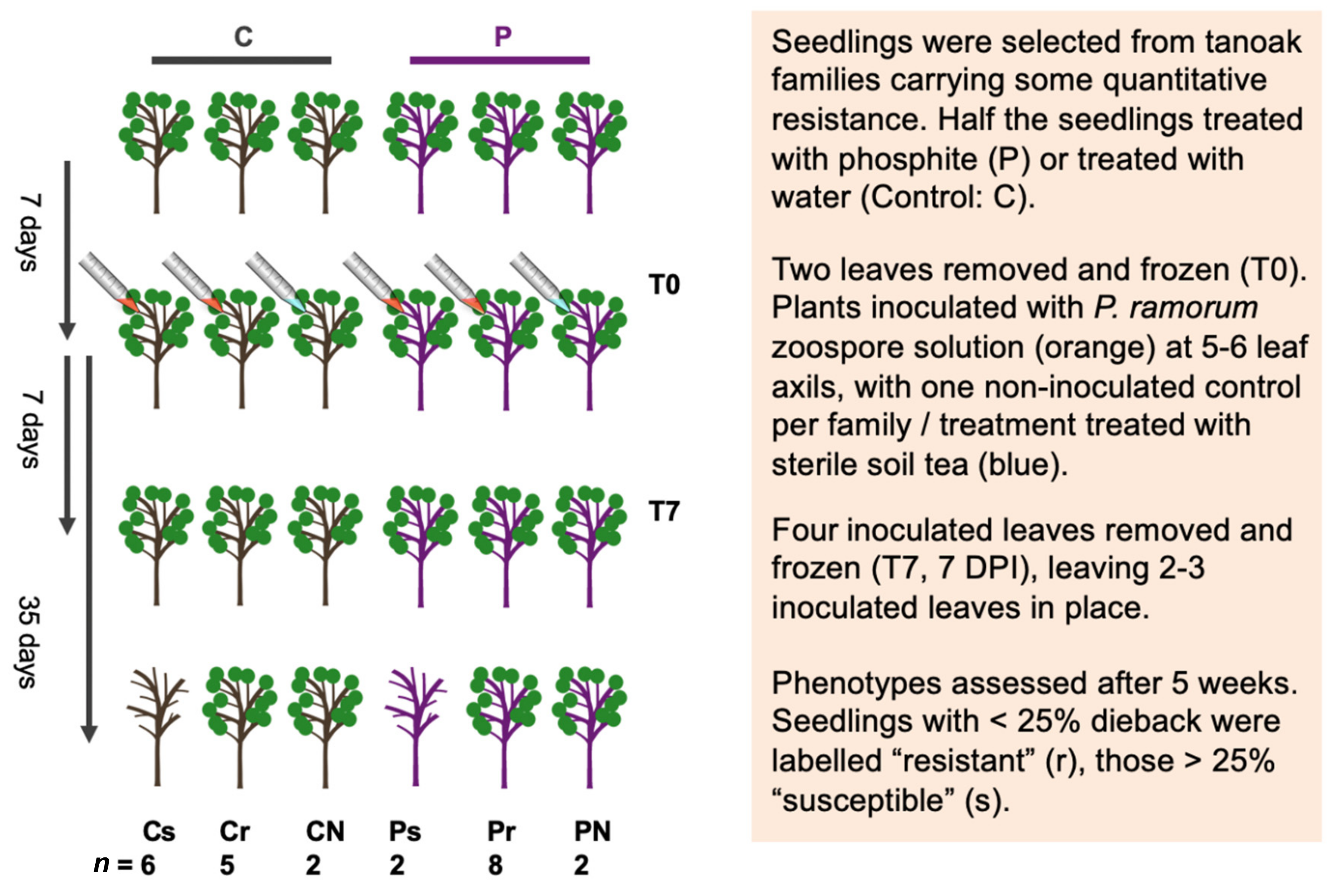
2.1. Experiment Overview
2.2. Experimental Design for Gene Expression Analysis
2.3. Phosphite Treatment
2.4. Inoculum and Leaf Harvest
2.5. RNA Extraction
2.6. Tanoak RNA-Seq cDNA Library Construction
2.7. Bioinformatics Pipeline
2.7.1. Tanoak Expression
2.7.2. P. ramorum Expression
2.7.3. Availability of Data and Material
3. Results
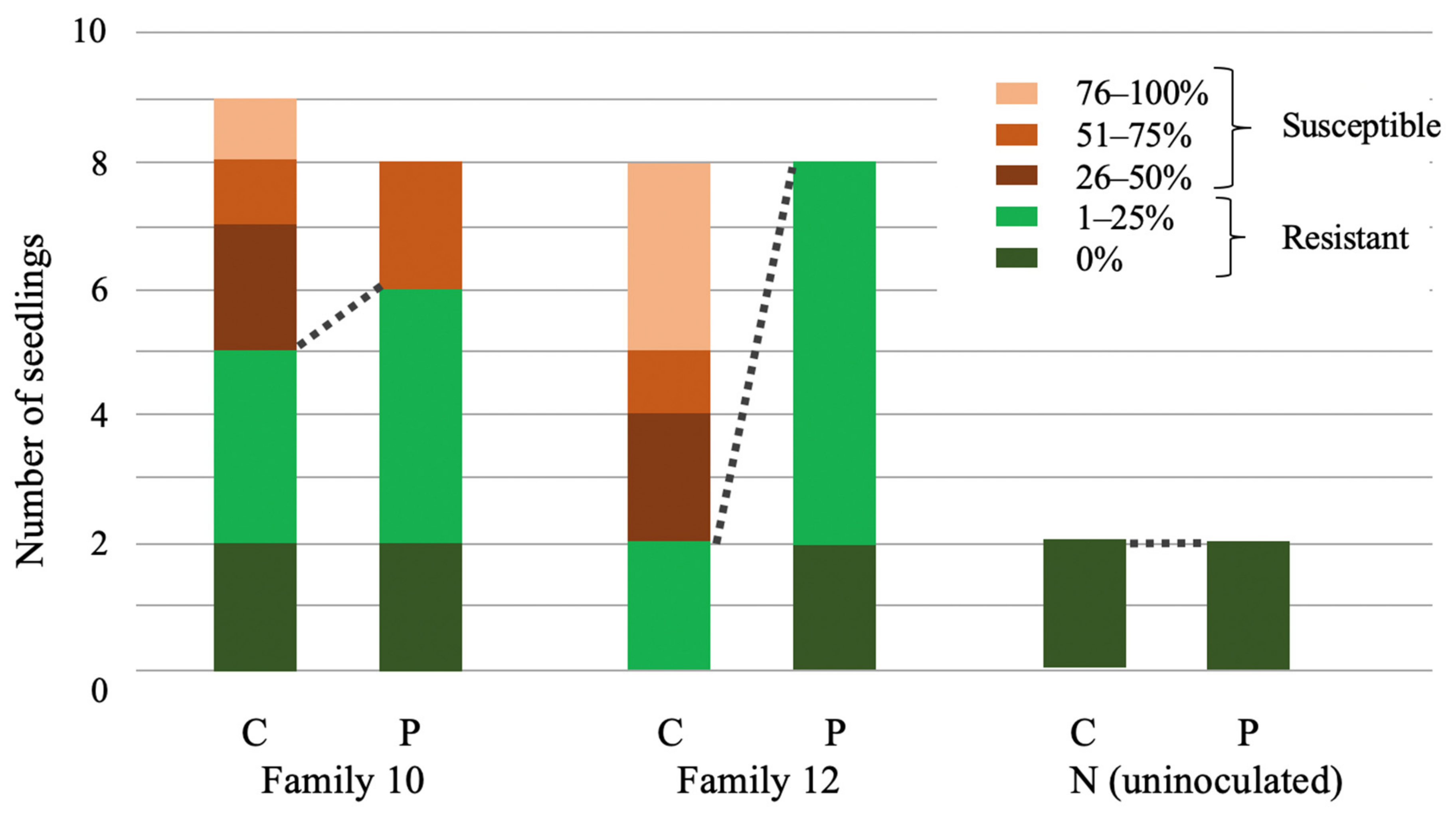
3.1. Tanoak Families Differed in the Presence of Innate Resistance and in the Effectiveness of Phosphite Treatment
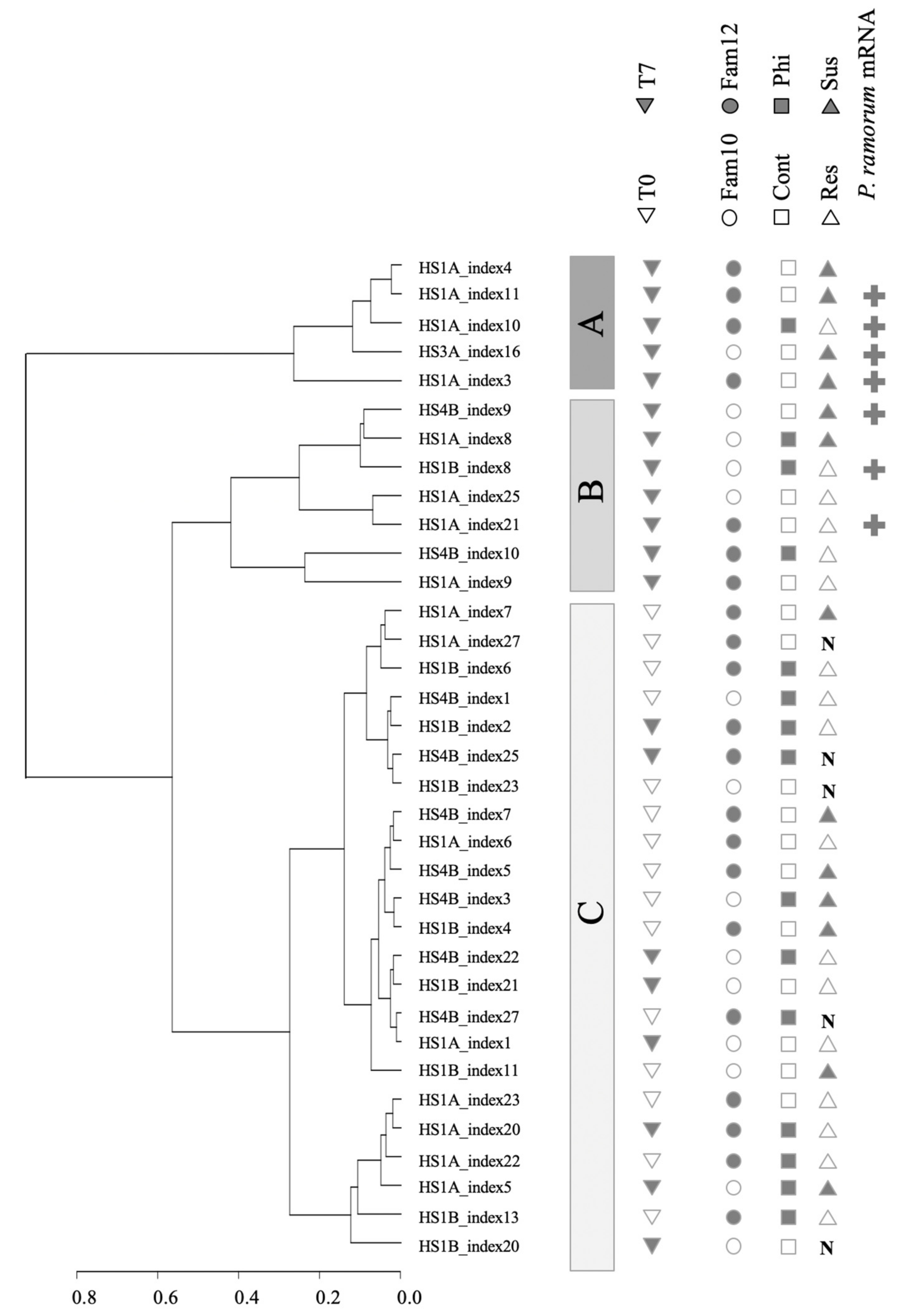
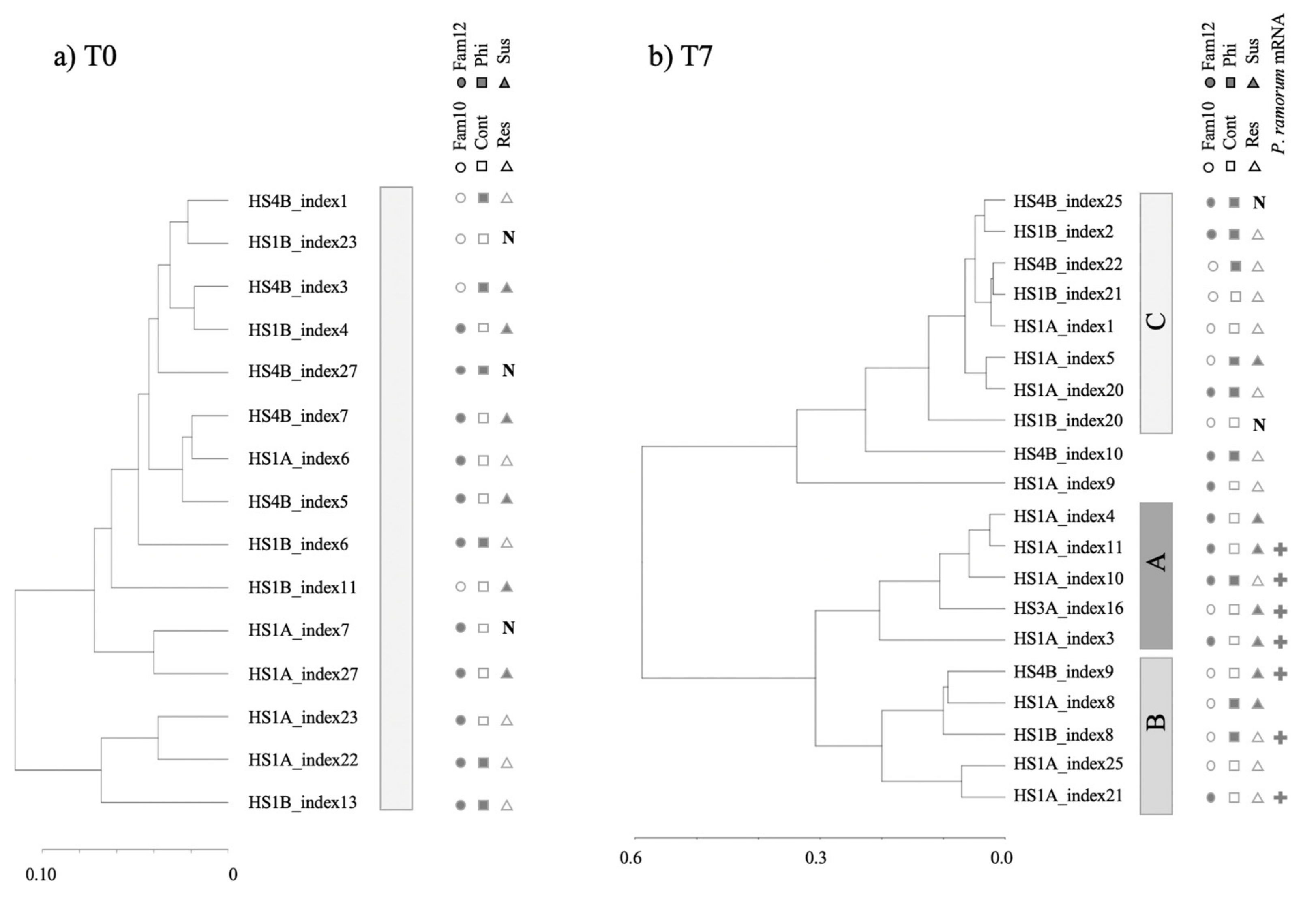
3.2. Clustering of Expression Revealed Factors Influencing Tanoak Transcriptomes
3.3. Search for Tanoak Transcriptome Signatures Associated with Innate Resistance and Phosphite Treatment
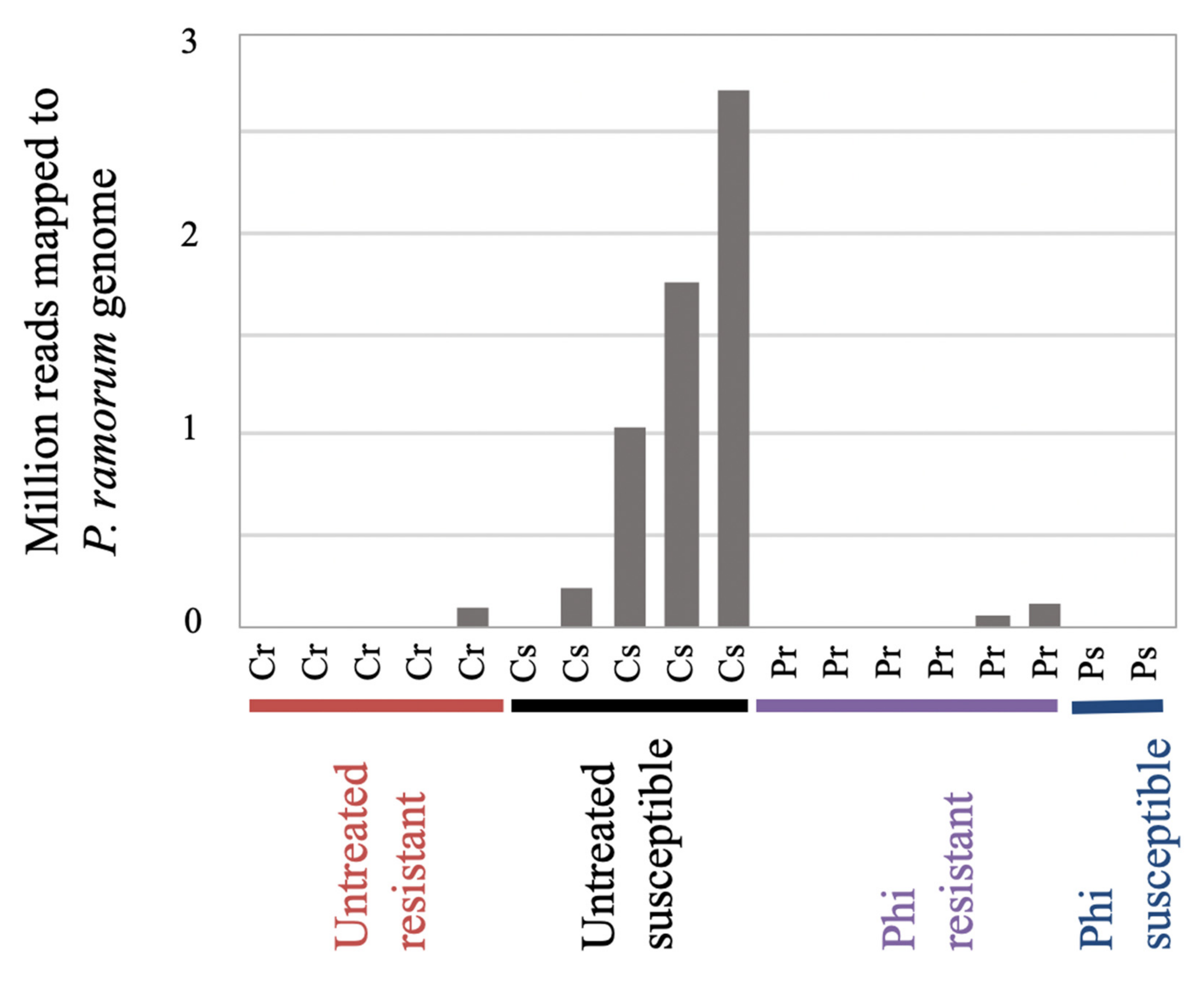
3.4. Analysis of in Planta Phytophthora Ramorum Transcriptomes
4. Discussion
4.1. Use of Organosilicate Surfactant for Phosphite Application
4.2. Genes Involved in the Innate Resistance
4.3. Phosphite-Induced Resistance
4.4. Use of English Oak as a Reference for Tanoak RNA-Seq Analysis
4.5. Phosphite-Induced Transcriptome Changes of P. ramorum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guest, D.; Grant, B. The Complex Action of Phosphonates as Antifungal Agents. Biol. Rev. 1991, 66, 159–187. [Google Scholar] [CrossRef]
- Hardy, G.E.S.J.; Barrett, S.; Shearer, B.L. The Future of Phosphite as a Fungicide to Control the Soilborne Plant Pathogen Phytophthora Cinnamomi in Natural Ecosystems. Australas. Plant Pathol. 2001, 30, 133–139. [Google Scholar] [CrossRef]
- Solla, A.; Moreno, G.; Malewski, T.; Jung, T.; Klisz, M.; Tkaczyk, M.; Siebyla, M.; Pérez, A.; Cubera, E.; Hrynyk, H.; et al. Phosphite Spray for the Control of Oak Decline Induced by Phytophthora in Europe. For. Ecol. Manag. 2021, 485, 118938. [Google Scholar] [CrossRef]
- Dalio, R.J.D.; Fleischmann, F.; Humez, M.; Osswald, W. Phosphite Protects Fagus Sylvatica Seedlings towards Phytophthora Plurivora via Local Toxicity, Priming and Facilitation of Pathogen Recognition. PLoS ONE 2014, 9, e87860. [Google Scholar] [CrossRef] [Green Version]
- Smillie, R.; Grant, B.; Guest, D. The Mode of Action of Phosphite: Evidence for Both Direct and Indirect Modes of Action on 3 Phytophthora Spp in Plants. Phytopathology 1989, 79, 921–926. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, C.J.; Holmes, J.M.; Dell, B.; Tynan, K.M.; McComb, J.A.; Shearer, B.L.; Colquhoun, I.J.; Hardy, G.E.S. Effect of Phosphite on in Planta Zoospore Production of Phytophthora Cinnamomi. Plant Pathol. 2001, 50, 587–593. [Google Scholar] [CrossRef]
- Grant, B.; Dunstan, R.; Griffith, J.; Niere, J.; Smillie, R. The Mechanism of Phosphonic (Phosphorous) Acid Action in Phytophthora. Australas. Plant Pathol. 1990, 19, 115. [Google Scholar] [CrossRef]
- Jackson, T.J.; Burgess, T.; Colquhoun, I.; Hardy, G. Action of the Fungicide Phosphite on Eucalyptus Marginata Inoculated with Phytophthora Cinnamomi. Plant Pathol. 2000, 49, 147–154. [Google Scholar] [CrossRef]
- Massoud, K.; Barchietto, T.; Rudulier, T.L.; Pallandre, L.; Didierlaurent, L.; Garmier, M.; Ambard-Bretteville, F.; Seng, J.-M.; Saindrenan, P. Dissecting Phosphite-Induced Priming in Arabidopsis Infected with Hyaloperonospora Arabidopsidis. Plant Physiol. 2012, 159, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Guest, D.; Bompeix, G. The Complex Mode of Action of Phosphonates. Australas. Plant Pathol. 1990, 19, 113. [Google Scholar] [CrossRef]
- Daniel, R.; Guest, D. Defence Responses Induced by Potassium Phosphonate in Phytophthora Palmivora-Challenged Arabidopsis Thaliana. Physiol. Mol. Plant Pathol. 2005, 67, 194–201. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Garbelotto, M. Sudden Oak Death: Endangering California and Oregon Forest Ecosystems. Front. Ecol. Environ. 2003, 1, 197–204. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Garbelotto, M.; Hansen, E.A. Phytophthora Ramorum: Integrative Research and Management of an Emerging Pathogen in California and Oregon Forests. Annu. Rev. Phytopathol. 2005, 43, 309–335. [Google Scholar] [CrossRef] [Green Version]
- Parke, J.L.; Peterson, E.K. Sudden Oak Death, Sudden Larch Death, and Ramorum Blight. Available online: https://www.apsnet.org/edcenter/disandpath/oomycete/pdlessons/Pages/SuddenOakDeath.aspx (accessed on 8 March 2021).
- Maloney, P.; Lynch, S.; Kane, S.; Jensen, C.; Rizzo, D. Establishment of an Emerging Generalist Pathogen in Redwood Forest Communities. J. Ecol. 2005, 93, 899–905. [Google Scholar] [CrossRef]
- McPherson, B.; Mori, S.; Wood, D.; Storer, A.; Svihra, P.; Kelly, N.; Standiford, R. Sudden Oak Death in California: Disease Progression in Oaks and Tanoaks. For. Ecol. Manag. 2005, 213, 71–89. [Google Scholar] [CrossRef]
- Meentemeyer, R.; Rank, N.; Shoemaker, D.; Oneal, C.; Wickland, A.; Frangioso, K.; Rizzo, D. Impact of Sudden Oak Death on Tree Mortality in the Big Sur Ecoregion of California. Biol. Invasions 2008, 10, 1243–1255. [Google Scholar] [CrossRef]
- Dillon, W.W.; Meentemeyer, R.K.; Vogler, J.B.; Cobb, R.C.; Metz, M.R.; Rizzo, D.M. Range-Wide Threats to a Foundation Tree Species from Disturbance Interactions. Madroño 2013, 60, 139–150. [Google Scholar] [CrossRef]
- Filipe, J.A.N.; Cobb, R.C.; Salmon, M.; Gilligan, C.A. Management Strategies for Conservation of Tanoak in California Forests Threatened by Sudden Oak Death: A Disease-Community Feedback Modelling Approach. Forests 2019, 10, 1103. [Google Scholar] [CrossRef] [Green Version]
- Hayden, K.J.; Garbelotto, M.; Dodd, R.; Wright, J.W. Scaling up from Greenhouse Resistance to Fitness in the Field for a Host of an Emerging Forest Disease. Evol. Appl. 2013, 6, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Garbelotto, M.; Schmidt, D.J. Phosphonate Controls Sudden Oak Death Pathogen for up to 2 Years. Calif. Agric. 2009, 63, 10–17. [Google Scholar] [CrossRef]
- Bowcutt, F. The Tanoak Tree: An Environmental History of a Pacific Coast Hardwood; University of Washington Press: Seattle, 2015; ISBN 978-0-295-99464-2. [Google Scholar]
- Nettel, A.; Dodd, R.; Afzal-Rafii, Z. Genetic Diversity, Structure, and Demographic Change in Tanoak, Lithocarpus Densiflorus (Fagaceae), the Most Susceptible Species to Sudden Oak Death in California. Am. J. Bot. 2009, 96, 2224–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, B.; Kirchner, J. Evolutionary Dynamics of Pathogen Resistance and Tolerance. Evolution 2000, 54, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Tyler, B.; Tripathy, S.; Zhang, X.; Dehal, P.; Jiang, R.; Aerts, A.; Arredondo, F.; Baxter, L.; Bensasson, D.; Beynon, J.; et al. Phytophthora Genome Sequences Uncover Evolutionary Origins and Mechanisms of Pathogenesis. Science 2006, 313, 1261–1266. [Google Scholar] [CrossRef] [Green Version]
- Hayden, K.J.; Garbelotto, M.; Knaus, B.J.; Cronn, R.C.; Rai, H.; Wright, J.W. Dual RNA-Seq of the Plant Pathogen Phytophthora Ramorum and Its Tanoak Host. Tree Genet. Genomes 2014, 10, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Baker, K.F. The U. C. System for Producing Healthy Container-Grown Plants through the Use of Clean Soil, Clean Stock, and Sanitation; University of California, Division of Agricultural Sciences, Agricultural Experiment Station, Extension Service: Berkeley, CA, USA, 1957. [Google Scholar]
- Garbelotto, M.; Schmidt, D.J.; Harnik, T.Y. Phosphite Injections and Bark Application of Phosphite + PentrabarkTM Control Sudden Oak Death in Coast Live Oak. Arboricult. Urban For. 2007, 309–317. [Google Scholar]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press: St. Paul, MN, USA, 1996; ISBN 0-89054-212-0. [Google Scholar]
- Chang, S.; Puryear, J.; Cairney, J. A Simple and Efficient Method for Isolating RNA from Pine Trees. Plant Mol. Biol. Rep. 2007, 11, 113–116. [Google Scholar] [CrossRef]
- Le Provost, G.; Herrera, R.; Paiva, J.; Chaumeil, P.; Salin, F.; Plomion, C. A Micromethod for High Throughput RNA Extraction in Forest Trees. Biol. Res. 2007, 40, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.-S.; Haj-Ahmod, Y. Evaluation of Plant RNA Integrity Number (RIN) Generated Using an Agilent BioAnalyzer 2100. Available online: https://norgenbiotek.com/sites/default/files/resources/plant_microrna_purification_kit_evaluation_of_plant_rna_integrity_number_rin_generated_using_an_agilent_bioanalyzer_2100_application_notes_599.pdf (accessed on 14 January 2020).
- Wang, L.; Nie, J.; Sicotte, H.; Li, Y.; Eckel-Passow, J.E.; Dasari, S.; Vedell, P.T.; Barman, P.; Wang, L.; Weinshiboum, R.; et al. Measure Transcript Integrity Using RNA-Seq Data. BMC Bioinform. 2016, 17, 58. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. The R Package Rsubread Is Easier, Faster, Cheaper and Better for Alignment and Quantification of RNA Sequencing Reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef] [Green Version]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 9 March 2021).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- The Gene Ontology Consortium Expansion of the Gene Ontology Knowledgebase and Resources. Nucleic Acids Res. 2017, 45, D331–D338. [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A Universal Tool for Annotation, Visualization and Analysis in Functional Genomics Research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Plomion, C.; Aury, J.-M.; Amselem, J.; Leroy, T.; Murat, F.; Duplessis, S.; Faye, S.; Francillonne, N.; Labadie, K.; Provost, G.L.; et al. Oak Genome Reveals Facets of Long Lifespan. Nat. Plants 2018, 4, 440. [Google Scholar] [CrossRef] [Green Version]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster Analysis and Display of Genome-Wide Expression Patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Garbelotto, M.; Harnik, T.Y.; Schmidt, D.J. Efficacy of Phosphonic Acid, Metalaxyl-M and Copper Hydroxide against Phytophthora Ramorum in Vitro and in Planta. Plant Pathol. 2009, 58, 111–119. [Google Scholar] [CrossRef]
- Dixon, R.; Paiva, N. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Bilgin, D.D.; Zavala, J.A.; Zhu, J.; Clough, S.J.; Ort, D.R.; DeLucia, E.H. Biotic Stress Globally Downregulates Photosynthesis Genes. Plant Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Dong, X.; Zhang, Z. Network-Based Comparative Analysis of Arabidopsis Immune Responses to Golovinomyces Orontii and Botrytis Cinerea Infections. Sci. Rep. 2016, 6, 19149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; He, F.; Zhang, Z. Large-Scale Transcriptome Analysis Reveals Arabidopsis Metabolic Pathways Are Frequently Influenced by Different Pathogens. Plant Mol. Biol. 2017, 94, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide Emergence of Resistance to Antifungal Drugs Challenges Human Health and Food Security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- French, E.; Kim, B.-S.; Iyer-Pascuzzi, A.S. Mechanisms of Quantitative Disease Resistance in Plants. Semin. Cell Dev. Biol. 2016, 56, 201–208. [Google Scholar]
- Poland, J.A.; Balint-Kurti, P.J.; Wisser, R.J.; Pratt, R.C.; Nelson, R.J. Shades of Gray: The World of Quantitative Disease Resistance. Trends Plant Sci. 2009, 14, 21–29. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.-J.; Reddy, M.S.S.; Wang, L. The Phenylpropanoid Pathway and Plant Defence—a Genomics Perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic Acid Signaling Pathway in Plants. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.-X.; Lou, Y.-G.; Mao, Y.-B.; Lu, S.; Wang, L.-J.; Chen, X.-Y. Plant Terpenoids: Biosynthesis and Ecological Functions. J. Integr. Plant Biol. 2007, 49, 179–186. [Google Scholar] [CrossRef]
- Zumaquero, A.; Martínez-Ferri, E.; Matas, A.J.; Reeksting, B.; Olivier, N.A.; Pliego-Alfaro, F.; Barceló, A.; van den Berg, N.; Pliego, C. Rosellinia Necatrix Infection Induces Differential Gene Expression between Tolerant and Susceptible Avocado Rootstocks. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Kandel, S.L.; Hulse-Kemp, A.M.; Stoffel, K.; Koike, S.T.; Shi, A.; Mou, B.; Van Deynze, A.; Klosterman, S.J. Transcriptional Analyses of Differential Cultivars during Resistant and Susceptible Interactions with Peronospora Effusa, the Causal Agent of Spinach Downy Mildew. Sci. Rep. 2020, 10, 6719. [Google Scholar] [CrossRef] [Green Version]
- Burra, D.D.; Berkowitz, O.; Hedley, P.E.; Morris, J.; Resjö, S.; Levander, F.; Liljeroth, E.; Andreasson, E.; Alexandersson, E. Phosphite-Induced Changes of the Transcriptome and Secretome in Solanum Tuberosum Leading to Resistance against Phytophthora Infestans. BMC Plant Biol. 2014, 14, 254. [Google Scholar] [CrossRef] [Green Version]
- Pagliari, L.; Buoso, S.; Santi, S.; Furch, A.C.; Martini, M.; Degola, F.; Loschi, A.; Bel, V.; Je, A.; Musetti, R. Filamentous Sieve Element Proteins Are Able to Limit Phloem Mass Flow, but Not Phytoplasma Spread. J. Exp. Bot. 2017, 68, 3673–3688. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.B.N.; Moller, E.; Gülz, P.-G. Epicuticular Waxes from Leaves of Quercus Robur. Phytochemistry 1990, 29, 2101–2103. [Google Scholar] [CrossRef]
- Johann, S.; Soldi, C.; Lyon, J.P.; Pizzolatti, M.G.; Resende, M.A. Antifungal Activity of the Amyrin Derivatives and in Vitro Inhibition of Candida Albicans Adhesion to Human Epithelial Cells. Lett. Appl. Microbiol. 2007, 45, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gómez, L.; Boller, T. FLS2: An LRR Receptor-like Kinase Involved in the Perception of the Bacterial Elicitor Flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Mathesius, U. Flavonoid Functions in Plants and Their Interactions with Other Organisms. Plants 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Fenn, M.; Coffey, M. Studies on the in Vitro and in Vivo Antifungal Activity of Fosetyl-Al and Phosphorus Acid. Phytopathology 1984, 74, 606–611. [Google Scholar] [CrossRef]
- King, M.; Reeve, W.; Van der Hoek, M.B.; Williams, N.; McComb, J.; O’Brien, P.A.; Hardy, G.E.S.J. Defining the Phosphite-Regulated Transcriptome of the Plant Pathogen Phytophthora Cinnamomi. Mol. Genet. Genomics 2010, 284, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Patkar, R.N.; Xue, Y.K.; Shui, G.; Wenk, M.R.; Naqvi, N.I. Abc3-Mediated Efflux of an Endogenous Digoxin-like Steroidal Glycoside by Magnaporthe Oryzae Is Necessary for Host Invasion during Blast Disease. PLoS Pathog. 2012, 8, e1002888. [Google Scholar] [CrossRef]
- Fleiβner, A.; Sopalla, C.; Weltring, K.-M. An ATP-Binding Cassette Multidrug-Resistance Transporter Is Necessary for Tolerance of Gibberella Pulicaris to Phytoalexins and Virulence on Potato Tubers. Mol. Plant Microbe Interact. 2002, 15, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (Pyridoxine) and Its Derivatives Are Efficient Singlet Oxygen Quenchers and Potential Fungal Antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Samsatly, J.; Copley, T.R.; Jabaji, S.H. Antioxidant Genes of Plants and Fungal Pathogens Are Distinctly Regulated during Disease Development in Different Rhizoctonia Solani Pathosystems. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | Family | N v N 1 | DE Up 2 | GO and KEGG 3 | DE Down 2 | GO 3 |
|---|---|---|---|---|---|---|
| AB up | AB down | |||||
| AB and C clusters | F10 & F12 | 12 v 23 | 7178 | 226, 40 | 6265 | 143, 8 |
| Cr up | Cr down | |||||
| Cr and Cs plants at T0 | F12 | 4 v 2 | 268 | 104, 4 | 198 | 6, 1 |
| P up | P down | |||||
| P and C plants at T0 | F12 | 6 v 3 | 0 | 0, 0 | 0 | 0, 0 |
| Pr up | Pr down | |||||
| Pr and Cs plants at T0 | F12 | 4 v 3 | 31 | 2, 0 | 16 | 0, 0 |
| GO and KEGG ID 1 | Cluster 2 | Description | False Discovery Rate 3 |
|---|---|---|---|
| map01130 | AB_up | Biosynthesis of antibiotics | 7.76 × 10−17 |
| map00980 | AB_up | Metabolism of xenobiotics by cytochrome P450 | 2.57 × 10−11 |
| map00010 | AB_up | Glycolysis/Gluconeogenesis | 2.57 × 10−9 |
| map00982 | AB_up | Drug metabolism-cytochrome P450 | 2.88 × 10−9 |
| map00480 | AB_up | Glutathione metabolism | 4.26 × 10−9 |
| map00983 | AB_up | Drug metabolism-other enzymes | 6.02 × 10−9 |
| map00520 | AB_up | Amino sugar and nucleotide sugar metabolism | 6.93 × 10−9 |
| map04660 | AB_up | T cell receptor signaling pathway | 1.46 × 10−6 |
| map00020 | AB_up | Citrate cycle (TCA cycle) | 1.56 × 10−6 |
| map04658 | AB_up | Th1 and Th2 cell differentiation | 1.81 × 10−5 |
| map00400 | AB_up | Phenylalanine, tyrosine and tryptophan biosynthesis | 6.29 × 10−5 |
| map00230 | AB_up | Purine metabolism | 6.48 × 10−5 |
| map00940 | AB_up | Phenylpropanoid biosynthesis | 6.63 × 10−5 |
| map00730 | AB_up | Thiamine metabolism | 7.04 × 10−5 |
| map00830 | AB_up | Retinol metabolism | 7.92 × 10−5 |
| map00720 | AB_up | Carbon fixation pathways in prokaryotes | 1.80 × 10−4 |
| map00071 | AB_up | Fatty acid degradation | 2.50 × 10−4 |
| map00592 | AB_up | alpha-Linolenic acid metabolism | 2.89 × 10−4 |
| map00625 | AB_up | Chloroalkane and chloroalkene degradation | 1.29 × 10−3 |
| map00620 | AB_up | Pyruvate metabolism | 1.30 × 10−3 |
| map00680 | AB_up | Methane metabolism | 1.37 × 10−3 |
| map00640 | AB_up | Propanoate metabolism | 1.41 × 10−3 |
| map00760 | AB_up | Nicotinate and nicotinamide metabolism | 2.01 × 10−3 |
| map00626 | AB_up | Naphthalene degradation | 2.58 × 10−3 |
| map00260 | AB_up | Glycine, serine and threonine metabolism | 3.12 × 10−3 |
| map00270 | AB_up | Cysteine and methionine metabolism | 3.48 × 10−3 |
| map00040 | AB_up | Pentose and glucuronate interconversions | 6.21 × 10−3 |
| map00350 | AB_up | Tyrosine metabolism | 6.84 × 10−3 |
| map00051 | AB_up | Fructose and mannose metabolism | 9.69 × 10−3 |
| GO:0016567 | AB_up | protein ubiquitination | 4.16 × 10−9 |
| GO:0010951 | AB_up | negative regulation of endopeptidase activity | 5.62 × 10−8 |
| GO:0006888 | AB_up | endoplasmic reticulum to Golgi vesicle-mediated transport | 1.02 × 10−4 |
| GO:0006749 | AB_up | glutathione metabolic process | 1.68 × 10−4 |
| GO:0006032 | AB_up | chitin catabolic process | 3.14 × 10−4 |
| GO:0032482 | AB_up | Rab protein signal transduction | 3.14 × 10−4 |
| GO:0006468 | AB_up | protein phosphorylation | 3.99 × 10−4 |
| GO:0031640 | AB_up | killing of cells of other organism | 4.86 × 10−4 |
| GO:0009435 | AB_up | NAD biosynthetic process | 1.02 × 10−3 |
| GO:0010200 | AB_up | response to chitin | 1.57 × 10−3 |
| GO:0006099 | AB_up | tricarboxylic acid cycle | 1.57 × 10−3 |
| GO:0016998 | AB_up | cell wall macromolecule catabolic process | 3.51 × 10−3 |
| GO:0006614 | AB_up | SRP-dependent cotranslational protein targeting to membrane | 3.51 × 10−3 |
| GO:0009694 | AB_up | jasmonic acid metabolic process | 3.76 × 10−3 |
| GO:0006096 | AB_up | glycolytic process | 4.47 × 10−3 |
| GO:0006457 | AB_up | protein folding | 6.42 × 10−3 |
| GO:0061025 | AB_up | membrane fusion | 6.54 × 10−3 |
| GO:0002181 | AB_up | cytoplasmic translation | 8.26 × 10−3 |
| GO:0009423 | AB_up | chorismate biosynthetic process | 9.43 × 10−3 |
| GO:0009873 | AB_up | ethylene-activated signaling pathway | 9.66 × 10−3 |
| map00860 | AB_down | Porphyrin and chlorophyll metabolism | 4.23 × 10−5 |
| map00670 | AB_down | One carbon pool by folate | 1.14 × 10−3 |
| map00970 | AB_down | Aminoacyl-tRNA biosynthesis | 2.79 × 10−3 |
| GO:0010088 | AB_down | phloem development | 5.04 × 10−5 |
| GO:0009768 | AB_down | photosynthesis, light harvesting in photosystem I | 1.06 × 10−4 |
| GO:0018298 | AB_down | protein-chromophore linkage | 1.22 × 10−4 |
| GO:0006298 | AB_down | mismatch repair | 1.78 × 10−3 |
| GO:0010206 | AB_down | photosystem II repair | 1.95 × 10−3 |
| GO:0006418 | AB_down | tRNA aminoacylation for protein translation | 2.36 × 10−3 |
| GO:0009234 | AB_down | menaquinone biosynthetic process | 3.43 × 10−3 |
| GO:0045037 | AB_down | protein import into chloroplast stroma | 3.43 × 10−3 |
| GO and KEGG ID 1 | DEG Category 2 | Description | False Discovery Rate |
|---|---|---|---|
| Cr v. Cs | |||
| GO:0010951 | Cr_up | negative regulation of endopeptidase activity | 3.15 × 10−9 |
| GO:0055114 | Cr_up | oxidation-reduction process | 1.35 × 10−6 |
| GO:0042744 | Cr_up | hydrogen peroxide catabolic process | 3.19 × 10−5 |
| GO:0016114 | Cr_up | terpenoid biosynthetic process | 2.08 × 10−4 |
| GO:0098869 | Cr_up | cellular oxidant detoxification | 3.11 × 10−4 |
| GO:0006979 | Cr_up | response to oxidative stress | 3.64 × 10−4 |
| GO:0009423 | Cr_up | chorismate biosynthetic process | 1.17 × 10−2 |
| GO:0031408 | Cr_up | oxylipin biosynthetic process | 1.17 × 10−2 |
| GO:0046129 | Cr_up | purine ribonucleoside biosynthetic process | 1.73 × 10−2 |
| GO:0009072 | Cr_up | aromatic amino acid family metabolic process | 2.47 × 10−2 |
| GO:0006833 | Cr_up | water transport | 3.06 × 10−2 |
| GO:0008654 | Cr_up | phospholipid biosynthetic process | 3.34 × 10−2 |
| GO:0030245 | Cr_up | cellulose catabolic process | 4.40 × 10−2 |
| GO:0009742 | Cr_up | brassinosteroid mediated signaling pathway | 4.40 × 10−2 |
| GO:0046130 | Cr_up | purine ribonucleoside catabolic process | 4.45 × 10−2 |
| GO:0009693 | Cr_up | ethylene biosynthetic process | 4.45 × 10−2 |
| GO:0010087 | Cr_up | phloem or xylem histogenesis | 4.67 × 10−2 |
| map00940 | Cr_up | Phenylpropanoid biosynthesis | 8.47 × 10−10 |
| map01130 | Cr_up | Biosynthesis of antibiotics | 2.10 × 10−6 |
| map00270 | Cr_up | Cysteine and methionine metabolism | 6.46 × 10−3 |
| map00500 | Cr_up | Starch and sucrose metabolism | 4.97 × 10−2 |
| GO:0006417 | Cr_down | regulation of translation | 4.33 × 10−2 |
| GO:0006075 | Cr_down | (1->3)-beta-d-glucan biosynthetic process | 4.33 × 10−2 |
| GO:0009682 | Cr_down | induced systemic resistance | 4.33 × 10−2 |
| map00500 | Cr_down | Starch and sucrose metabolism | 1.13 × 10−4 |
| Cr v. Cs & Cs v. Pr overlap | |||
| GO:0010088 | Cr_up & Pr_up | phloem development | 1.26 × 10−2 |
| GO:0016104 | Cr_up & Pr_up | triterpenoid biosynthetic process | 4.50 × 10−2 |
| DEG Category 1 | Cs and Pr at T0 (Phi-Induced Resistance) | Cs and Cr at T0 (Innate Resistance) | Overlap |
|---|---|---|---|
| Cs_up | 16 | 198 | 3 |
| Cs_down | 31 | 268 | 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasuga, T.; Hayden, K.J.; Eyre, C.A.; Croucher, P.J.P.; Schechter, S.; Wright, J.W.; Garbelotto, M. Innate Resistance and Phosphite Treatment Affect Both the Pathogen’s and Host’s Transcriptomes in the Tanoak-Phytophthora ramorum Pathosystem. J. Fungi 2021, 7, 198. https://doi.org/10.3390/jof7030198
Kasuga T, Hayden KJ, Eyre CA, Croucher PJP, Schechter S, Wright JW, Garbelotto M. Innate Resistance and Phosphite Treatment Affect Both the Pathogen’s and Host’s Transcriptomes in the Tanoak-Phytophthora ramorum Pathosystem. Journal of Fungi. 2021; 7(3):198. https://doi.org/10.3390/jof7030198
Chicago/Turabian StyleKasuga, Takao, Katherine J. Hayden, Catherine A. Eyre, Peter J. P. Croucher, Shannon Schechter, Jessica W. Wright, and Matteo Garbelotto. 2021. "Innate Resistance and Phosphite Treatment Affect Both the Pathogen’s and Host’s Transcriptomes in the Tanoak-Phytophthora ramorum Pathosystem" Journal of Fungi 7, no. 3: 198. https://doi.org/10.3390/jof7030198