
Do Candida albicans Isolates with Borderline Resistant Micafungin MICs Always Harbor FKS1 Hot Spot Mutations?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. DNA Extraction
2.3. Next-Generation Sequencing and Library Preparation
2.4. Bioinformatic Analysis
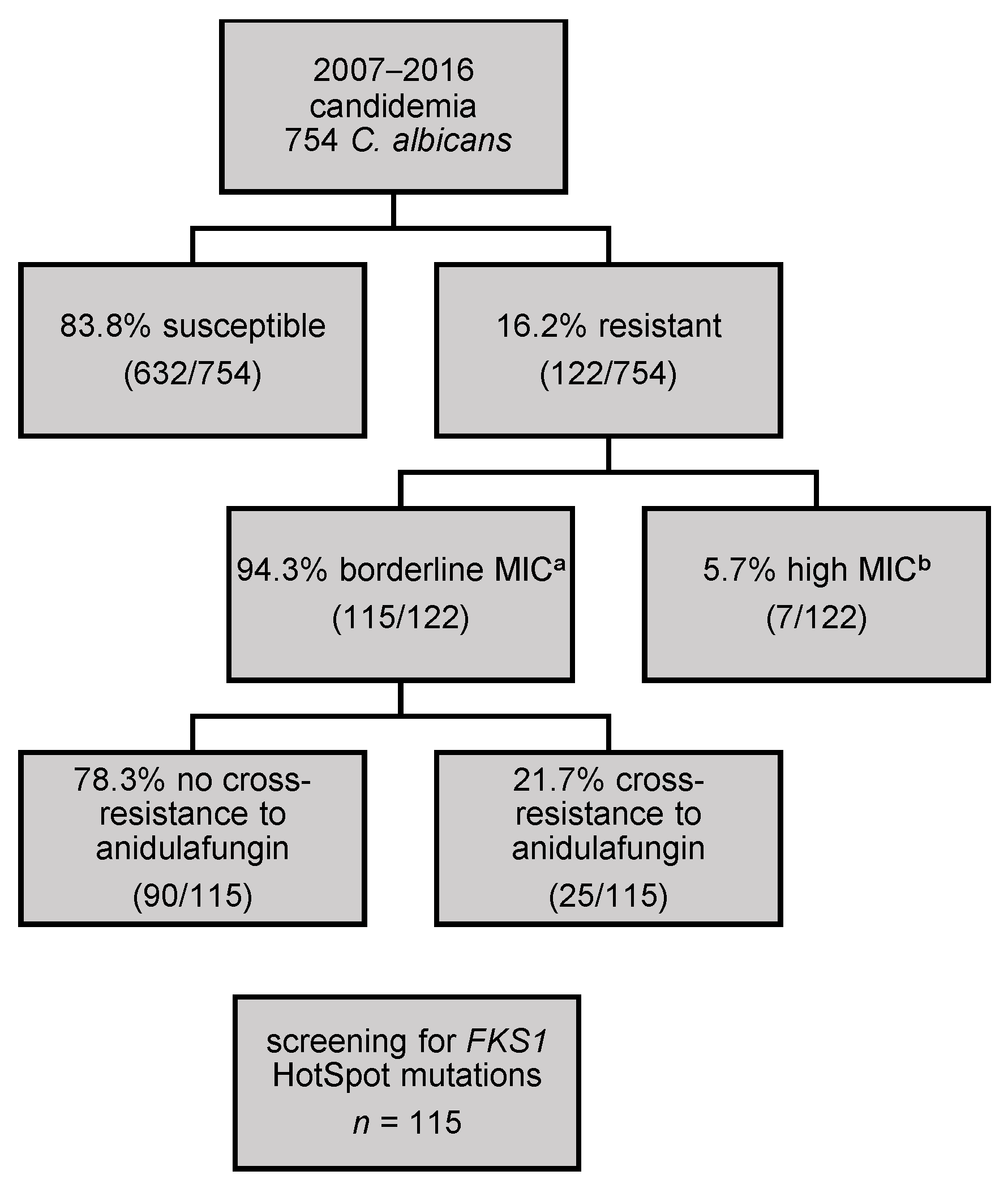
3. Results
3.1. Validation of the Sequencing Run
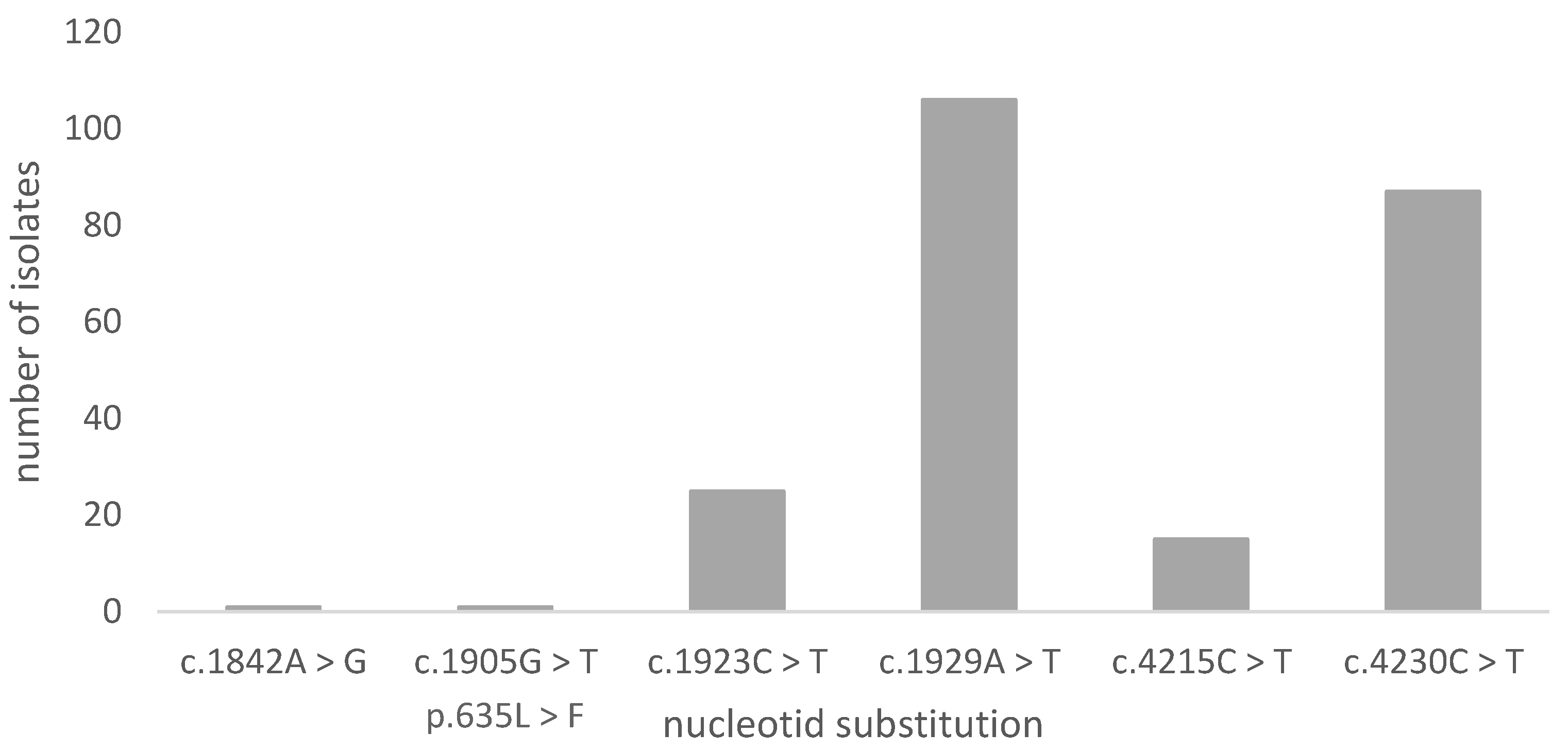
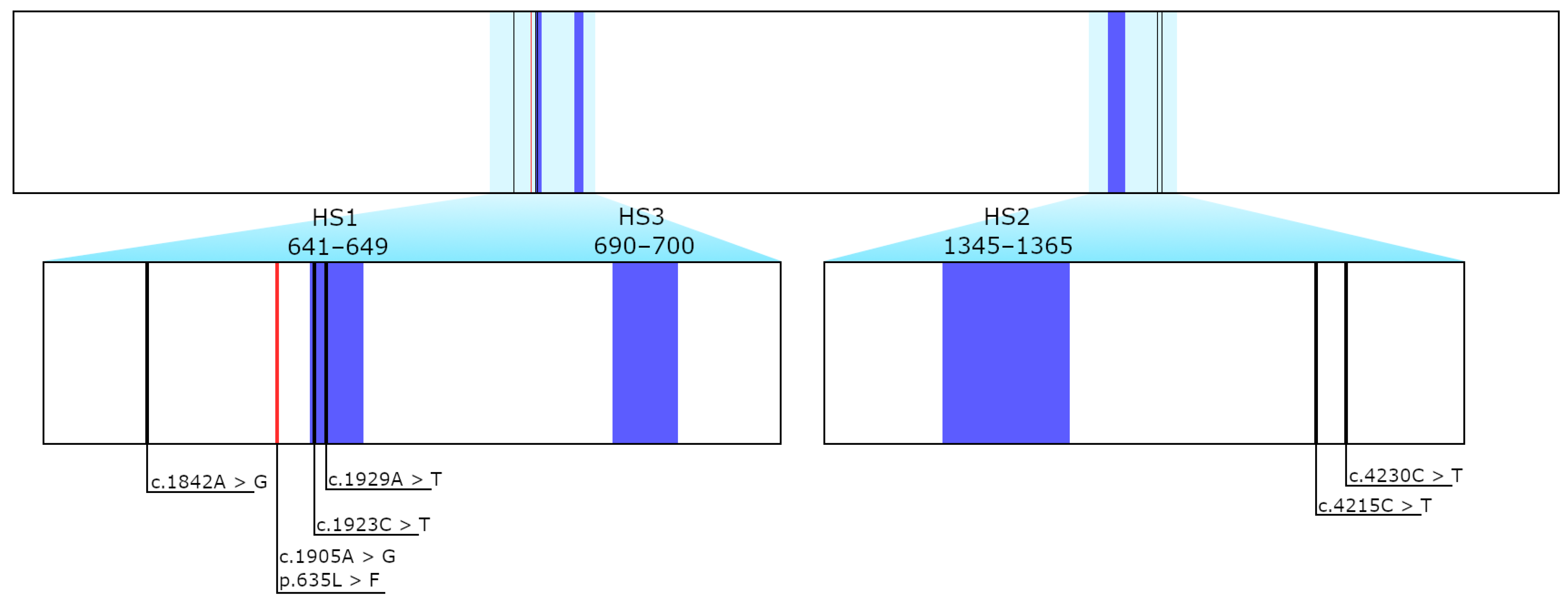
3.2. Sequencing Results of FKS1
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santolaya, M.E.; Thompson, L.; Benadof, D.; Tapia, C.; Legarraga, P.; Cortés, C.; Rabello, M.; Valenzuela, R.; Rojas, P.; Rabagliati, R.; et al. A prospective, multi-center study of Candida bloodstream infections in Chile. PLoS ONE 2019, 14, e0212924. [Google Scholar] [CrossRef] [Green Version]
- Bassetti, M.; Giacobbe, D.R.; Vena, A.; Trucchi, C.; Ansaldi, F.; Antonelli, M.; Adamkova, V.; Alicino, C.; Almyroudi, M.-P.; Atchade, E.; et al. Incidence and outcome of invasive candidiasis in intensive care units (ICUs) in Europe: Results of the EUCANDICU project. Crit. Care 2019, 23, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendrup, M.C.; Boekhout, T.; Akova, M.; Meis, J.F.; Cornely, O.A.; Lortholary, O.; Arikan-Akdagli, S.; Cuenca-Estrella, M.; Dannaoui, E.; van Diepeningen, A.D.; et al. ESCMID† and ECMM‡ joint clinical guidelines for the diagnosis and management of rare invasive yeast infections. Clin. Microbiol. Infect. 2014, 20 (Suppl. 3), 76–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, e1–e50. [Google Scholar] [CrossRef] [PubMed]
- Perlin, D.S. Echinocandin Resistance in Candida. Clin. Infect. Dis. 2015, 61, S612–S617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Kelly, R.; Kahn, J.N.; Robles, J.; Hsu, M.-J.; Register, E.; Li, W.; Vyas, V.; Fan, H.; Abruzzo, G.; et al. Specific Substitutions in the Echinocandin Target Fks1p Account for Reduced Susceptibility of Rare Laboratory and Clinical Candida sp. Isolates. Antimicrob. Agents Chemother. 2005, 49, 3264–3273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.E.; Edlind, T.D. Topological and Mutational Analysis of Saccharomyces cerevisiae Fks1. Eukaryot. Cell 2012, 11, 952–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willinger, B. AURES 2014—Resistenzbericht Hefepilze; Bundesministerium für Gesundheit: Berlin, Germany, 2015; pp. 173–194. ISBN 9783902611987. [Google Scholar]
- Willinger, B. AURES 2015—Resistenzbericht Hefepilze; Bundesministerium für Gesundheit und Frauen: Berlin, Germany, 2016; pp. 186–208. ISBN 9783903099111. [Google Scholar]
- Willinger, B. AURES 2016—Resistenzbericht Hefepilze; Bundesministerium für Gesundheit und Frauen: Berlin, Germany, 2017; pp. 189–211. ISBN 9783903099296. [Google Scholar]
- Willinger, B. AURES 2017—Resistenzbericht Hefepilze; Bundesministerium für Arbeit, Soziales, Gesundheit und Konsumentenschutz: Berlin, Germany, 2018; pp. 255–292. ISBN 9783850105156. [Google Scholar]
- Beyer, R.; Spettel, K.; Zeller, I.; Lass-Flörl, C.; Achleitner, D.; Krause, R.; Apfalter, P.; Buzina, W.; Strauss, J.; Gregori, C.; et al. Antifungal susceptibility of yeast bloodstream isolates collected during a 10-year period in Austria. Mycoses 2019, 62, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Spettel, K.; Barousch, W.; Makristathis, A.; Zeller, I.; Nehr, M.; Selitsch, B.; Lackner, M.; Rath, P.-M.; Steinmann, J.; Willinger, B. Analysis of antifungal resistance genes in Candida albicans and Candida glabrata using next generation sequencing. PLoS ONE 2019, 14, e0210397. [Google Scholar] [CrossRef] [PubMed]
- Umesha, S.; Manukumar, H.M.; Raghava, S. A rapid method for isolation of genomic DNA from food-borne fungal pathogens. 3 Biotech 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illumina Inc. 16S Metagenomic Sequencing Library Preparation—Preparing 16S Ribosomal RNA Gene Amplicons for the Illumina MiSeq System. In 16S Metagenomic Sequencing Library Preparation; Illumina Inc.: San Diego, CA, USA, 2013. [Google Scholar]
- Garnaud, C.; Botterel, F.; Sertour, N.; Bougnoux, M.-E.; Dannaoui, E.; Larrat, S.; Hennequin, C.; Guinea, J.; Cornet, M.; Maubon, D. Next-generation sequencing offers new insights into the resistance of Candida spp. to echinocandins and azoles. J. Antimicrob. Chemother. 2015, 70, 2556–2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 January 2016).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koboldt, D.C.; Chen, K.; Wylie, T.; Larson, D.E.; McLellan, M.D.; Mardis, E.R.; Weinstock, G.M.; Wilson, R.K.; Ding, L. VarScan: Variant detection in massively parallel sequencing of individual and pooled samples. Bioinform. 2009, 25, 2283–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs for Antifungal Agents, version 10.0; 2020. Available online: http://www.eucast.org/astoffungi/clinicalbreakpointsforantifungals/ (accessed on 20 March 2020).
- Arendrup, M.C.; Friberg, N.; Mares, M.; Kahlmeter, G.; Meletiadis, J.; Guinea, J.; Andersen, C.; Arikan-Akdagli, S.; Barchiesi, F.; Chryssanthou, E.; et al. How to interpret MICs of antifungal compounds according to the revised clinical breakpoints v. 10.0 European committee on antimicrobial susceptibility testing (EUCAST). Clin. Microbiol. Infect. 2020, 26, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.A.; Gow, N.A.R.; Munro, C.A. Elevated Chitin Content Reduces the Susceptibility of Candida Species to Caspofungin. Antimicrob. Agents Chemother. 2012, 57, 146–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imtiaz, T.; Lee, K.K.; Munro, C.A.; Maccallum, D.M.; Shankland, G.S.; Johnson, E.M.; MacGregor, M.S.; Bal, A.M. Echinocandin resistance due to simultaneous FKS mutation and increased cell wall chitin in a Candida albicans bloodstream isolate following brief exposure to caspofungin. J. Med. Microbiol. 2012, 61, 1330–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, A.; Ene, I.V.; Bibi, M.; Zakin, S.; Segal, E.S.; Ziv, N.; Dahan, A.M.; Colombo, A.L.; Bennett, R.J.; Berman, J. Antifungal tolerance is a subpopulation effect distinct from resistance and is associated with persistent candidemia. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| ID | Hotspot | Overhang- + Specific Locus Sequence | |
|---|---|---|---|
| 12F | 1, 3 | TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGGAACAAGAGATCAAGAAGATATA | Spettel 2019 |
| 12R | 1, 3 | GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGTGAACGACCAATGGAGAAGA | Spettel 2019 |
| 13F | 2 | TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCTATGGTCATCCAGGTTTCCA | Garnaud 2015 |
| 13R | 2 | GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGCACCAACGGTCAAATCAGTG | Garnaud 2015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spettel, K.; Galazka, S.; Kriz, R.; Camp, I.; Willinger, B. Do Candida albicans Isolates with Borderline Resistant Micafungin MICs Always Harbor FKS1 Hot Spot Mutations? J. Fungi 2021, 7, 93. https://doi.org/10.3390/jof7020093
Spettel K, Galazka S, Kriz R, Camp I, Willinger B. Do Candida albicans Isolates with Borderline Resistant Micafungin MICs Always Harbor FKS1 Hot Spot Mutations? Journal of Fungi. 2021; 7(2):93. https://doi.org/10.3390/jof7020093
Chicago/Turabian StyleSpettel, Kathrin, Sonia Galazka, Richard Kriz, Iris Camp, and Birgit Willinger. 2021. "Do Candida albicans Isolates with Borderline Resistant Micafungin MICs Always Harbor FKS1 Hot Spot Mutations?" Journal of Fungi 7, no. 2: 93. https://doi.org/10.3390/jof7020093