
Specificities and Synergistic Actions of Novel PL8 and PL7 Alginate Lyases from the Marine Fungus Paradendryphiella salina




Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrates
2.2. Sequence Analysis
2.3. Phylogenic Analysis of PL8 Sequences
2.4. Genes, Cloning, Expression, and Purification of PsAlg7A, -B, -C, and PsMan8A Alginate Lyases
2.5. Standard Assay Conditions
2.6. Biochemical Characterization
2.7. Kinetics of PsAlg7A, PsAlg7B, PsAlg7C, and PsMan8A
2.8. Synergy of PsAlg7A and PsMan8A
2.9. Action Pattern and Degradation Product Analysis by LC-MS
3. Results
3.1. Genetic Environment
3.2. Sequence Analysis
3.3. Blastp Analysis
3.4. Phylogenetic Analysis
3.5. Expression and Purification
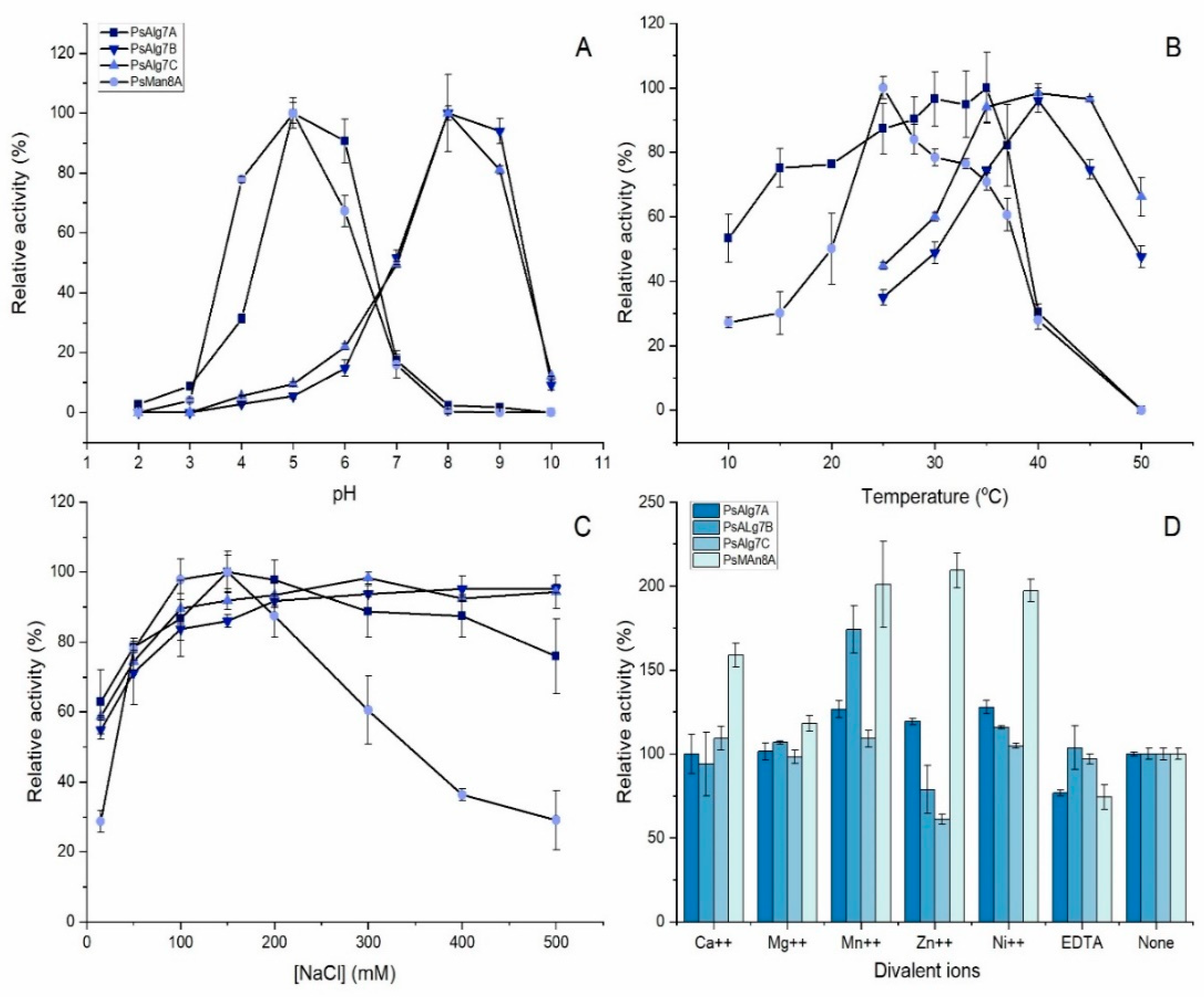
3.6. Biochemical Characterization
3.7. Kinetic Parameters of PsAlg7A, PsAlg7B, PsAlg7C, and PsMan8A
3.8. Synergy of PsAlg7A and PsMan8A
3.9. Product Profiles of P. salina Alginate Lyases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gladfelter, A.S.; James, T.Y.; Amend, A.S. Marine Fungi. Curr. Biol. 2019, 29, R191–R195. [Google Scholar] [CrossRef]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.H.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; et al. Fungi in the Marine Environment: Open Questions and Unsolved Problems. mBio 2019, 10, e01189-18. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.B.G. Are There More Marine Fungi to Be Described? Bot. Mar. 2011, 54, 343–354. [Google Scholar] [CrossRef]
- Margulis, L.; Chapman, M.J. Chapter Four—Kingdom Fungi. In Kingdoms and Domains, 4th ed.; Margulis, L., Chapman, M.J., Eds.; Academic Press: London, UK, 2009; pp. 379–409. ISBN 978-0-12-373621-5. [Google Scholar]
- Schoch, C.L.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.W.A.; Burgess, T.I.; de Gruyter, J.; de Hoog, G.S.; Dixon, L.J.; Grube, M.; Gueidan, C.; et al. A Class-Wide Phylogenetic Assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1–15. [Google Scholar] [CrossRef]
- Sutherland, G.K. Marine Fungi Imperfecti. New Phytol. 1916, 15, 35–48. [Google Scholar] [CrossRef]
- Jones, E.B.G. Marine Fungi. Trans. Br. Mycol. Soc. 1962, 45, 93–114. [Google Scholar] [CrossRef]
- Schaumann, K.; Weide, G. Enzymatic Degradation of Alginate by Marine Fungi. Hydrobiologia 1990, 204–205, 589–596. [Google Scholar] [CrossRef]
- Zuccaro, A.; Schoch, C.L.; Spatafora, J.W.; Kohlmeyer, J.; Draeger, S.; Mitchell, J.I. Detection and Identification of Fungi Intimately Associated with the Brown Seaweed Fucus Serratus. Appl. Environ. Microbiol. 2008, 74, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Gostinčar, C.; Muggia, L.; Grube, M. Polyextremotolerant Black Fungi: Oligotrophism, Adaptive Potential, and a Link to Lichen Symbioses. Front. Microbiol. 2012, 3, 390. [Google Scholar] [CrossRef] [Green Version]
- Rabillé, H.; Torode, T.A.; Tesson, B.; Le Bail, A.; Billoud, B.; Rolland, E.; Le Panse, S.; Jam, M.; Charrier, B. Alginates along the Filament of the Brown Alga Ectocarpus Help Cells Cope with Stress. Sci. Rep. 2019, 9, 12956. [Google Scholar] [CrossRef] [Green Version]
- Fourest, E.; Volesky, B. Alginate Properties and Heavy Metal Biosorption by Marine Algae. Appl. Biochem. Biotechnol. 1997, 67, 215–226. [Google Scholar] [CrossRef]
- Rhein-Knudsen, N.; Ale, M.T.; Meyer, A.S. Seaweed Hydrocolloid Production: An Update on Enzyme Assisted Extraction and Modification Technologies. Mar. Drugs 2015, 13, 3340–3359. [Google Scholar] [CrossRef]
- Draget, K.I.; Smidsrød, O.; Skjåk-Bræk, G. Alginates from Algae. Part 6. Polysaccharides. In Biopolymers Online; Wiley Online Library, Wiley-VCH GmbH: Weinheim, Germany, 2005; ISBN 978-3-527-60003-8. [Google Scholar] [CrossRef]
- Rye, P.D.; Tøndervik, A.; Sletta, H.; Pritchard, M.; Kristiansen, A.; Dessen, A.; Thomas, D.W. Alginate Oligomers and Their Use as Active Pharmaceutical Drugs. In Alginates and Their Biomedical Applications; Rehm, B.H.A., Moradali, M.F., Eds.; Springer Series in Biomaterials Science and Engineering; Springer: Singapore, 2018; pp. 237–256. ISBN 978-981-10-6910-9. [Google Scholar]
- Lombard, V.; Bernard, T.; Rancurel, C.; Brumer, H.; Coutinho, P.M.; Henrissat, B. A Hierarchical Classification of Polysaccharide Lyases for Glycogenomics. Biochem. J. 2010, 432, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Mashino, C.; Uji, T.; Saga, N.; Mikami, K.; Ojima, T. Characterization of an Eukaryotic PL-7 Alginate Lyase in the Marine Red Alga Pyropia yezoensis. Curr. Biotechnol. 2015, 4, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Ojima, T. Functional Identification of Alginate Lyase from the Brown Alga Saccharina japonica. Sci. Rep. 2019, 9, 4937. [Google Scholar] [CrossRef] [Green Version]
- Suda, K.; Tanji, Y.; Hori, K.; Unno, H. Evidence for a Novel Chlorella Virus-Encoded Alginate Lyase. FEMS Microbiol. Lett. 1999, 180, 45–53. [Google Scholar] [CrossRef]
- Sil’chenko, A.S.; Kusaikin, M.I.; Zakharenko, A.M.; Zvyagintseva, T.N. Isolation from the Marine Mollusk Lambis Sp. and Catalytic Properties of an Alginate Lyase with Rare Substrate Specificity. Chem. Nat. Compd. 2013, 49, 215–218. [Google Scholar] [CrossRef]
- Zhu, B.; Yin, H. Alginate Lyase: Review of Major Sources and Classification, Properties, Structure-Function Analysis and Applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Pilgaard, B.; Wilkens, C.; Herbst, F.-A.; Vuillemin, M.; Rhein-Knudsen, N.; Meyer, A.S.; Lange, L. Proteomic Enzyme Analysis of the Marine Fungus Paradendryphiella salina Reveals Alginate Lyase as a Minimal Adaptation Strategy for Brown Algae Degradation. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. DbCAN2: A Meta Server for Automated Carbohydrate-Active Enzyme Annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-Scale Protein Function Classification. Bioinform. Oxf. Engl. 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Käll, L.; Krogh, A.; Sonnhammer, E.L.L. Advantages of Combined Transmembrane Topology and Signal Peptide Prediction—the Phobius Web Server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Jung, E.; Brunak, S. Prediction of N-Glycosylation Sites in Human Proteins. 2004. Available online: http://www.cbs.dtu.dk/services2002 (accessed on 15 December 2020).
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum Likelihood-Based Phylogenetic Analyses with Thousands of Taxa and Mixed Models. Bioinform. Oxf. Engl. 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An Improved General Amino Acid Replacement Matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), IEEE, New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Wu, S.; Letchworth, G.J. High Efficiency Transformation by Electroporation of Pichia pastoris Pretreated with Lithium Acetate and Dithiothreitol. BioTechniques 2004, 36, 152–154. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. ISBN 978-1-58829-343-5. [Google Scholar]
- Brooke, D.; Movahed, N.; Bothner, B. Universal Buffers for Use in Biochemistry and Biophysical Experiments. Biophysics 2015, 2, 336–342. [Google Scholar] [CrossRef]
- Swift, S.M.; Hudgens, J.W.; Heselpoth, R.D.; Bales, P.M.; Nelson, D.C. Characterization of AlgMsp, an Alginate Lyase from Microbulbifer sp. 6532A. PLoS ONE 2014, 9, e112939. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, Y.; Iwasaki, K.; Muramatsu, T. Action of Poly (Alpha-Lguluronate) Lyase from Corynebacterium sp. ALY-1 Strain on Saturated Oligoguluronates. Biosci. Biotechnol. Biochem. 1998, 62, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.P.; Borch, K.; Westh, P. Endo/Exo-Synergism of Cellulases Increases with Substrate Conversion. Biotechnol. Bioeng. 2017, 114, 696–700. [Google Scholar] [CrossRef] [PubMed]
- Helbert, W.; Poulet, L.; Drouillard, S.; Mathieu, S.; Loiodice, M.; Couturier, M.; Lombard, V.; Terrapon, N.; Turchetto, J.; Vincentelli, R.; et al. Discovery of Novel Carbohydrate-Active Enzymes through the Rational Exploration of the Protein Sequences Space. Proc. Natl. Acad. Sci. USA 2019, 116, 6063–6068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and Sequencing of Alginate Lyase Genes from Deep-Sea Strains of Vibrio and Agarivorans and Characterization of a New Vibrio Enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a Novel Alginate Lyase from Nitratiruptor sp. SB155-2 Thriving at Deep-Sea Hydrothermal Vents and Identification of the Residues Responsible for Its Heat Stability. J. Biol. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, G.; Tonon, T.; Scornet, D.; Cock, J.M.; Kloareg, B. The Cell Wall Polysaccharide Metabolism of the Brown Alga Ectocarpus siliculosus. Insights into the Evolution of Extracellular Matrix Polysaccharides in Eukaryotes. New Phytol. 2010, 188, 82–97. [Google Scholar] [CrossRef]
- Balabanova, L.; Slepchenko, L.; Son, O.; Tekutyeva, L. Biotechnology Potential of Marine Fungi Degrading Plant and Algae Polymeric Substrates. Front. Microbiol. 2018, 9, 1527. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Gupta, V.; Kumari, P.; Kumar, M.; Reddy, C.R.K.; Prasad, K.; Jha, B. Purification and Partial Characterization of an Extracellular Alginate Lyase from Aspergillus oryzae Isolated from Brown Seaweed. J. Appl. Phycol. 2011, 23, 755–762. [Google Scholar] [CrossRef]
- Shimokawa, T.; Yoshida, S.; Takeuchi, T.; Murata, K.; Kobayashi, H.; Kusakabe, I. Purification and Characterization of Extracellular Poly(β -D-1,4-Mannuronide) Lyase from Dendryphiella salina IFO 32139. Biosci. Biotechnol. Biochem. 1997, 61, 636–640. [Google Scholar] [CrossRef]
- dela Cruz, T.E.E.; Schulz, B.E.; Kubicek, C.P.; Druzhinina, I.S. Carbon Source Utilization by the Marine Dendryphiella Species D. arenaria and D. salina. FEMS Microbiol. Ecol. 2006, 58, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Zhang, Z.; Linhardt, R.J. Chapter 3—Glycosaminoglycans. In Handbook of Glycomics; Cummings, R.D., Pierce, J.M., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 59–80. ISBN 978-0-12-373600-0. [Google Scholar]
- Hay, I.D.; Rehman, Z.U.; Moradali, M.F.; Wang, Y.; Rehm, B.H.A. Microbial Alginate Production, Modification and Its Applications. Microb. Biotechnol. 2013, 6, 637–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millero, F.J.; Feistel, R.; Wright, D.G.; McDougall, T.J. The Composition of Standard Seawater and the Definition of the Reference-Composition Salinity Scale. Deep Sea Res. Part I: Oceanogr. Res. Pap. 2008, 55, 50–72. [Google Scholar] [CrossRef]
- Nogueira, K.M.V.; Mendes, V.; Carraro, C.B.; Taveira, I.C.; Oshiquiri, L.H.; Gupta, V.K.; Silva, R.N. Sugar Transporters from Industrial Fungi: Key to Improving Second-Generation Ethanol Production. Renew. Sustain. Energy Rev. 2020, 131, 109991. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Parameters | Alginate | PolyM | PolyG |
|---|---|---|---|---|
| PsAlg7A | SA (U mg−1) | 245 ± 4.4 | 1263 ± 6.5 | n/a |
| Km (mM) | 3.9 ± 0.1 | 17.4 ± 0.3 | n/a | |
| kcat (s−1) | 0.65 ± 0.01 | 3.4 ± 0.02 | n/a | |
| kcat/Km | 0.16 | 0.19 | n/a | |
| PsAlg7B | SA (U mg−1) | 1008 ± 21 | 1459 ± 6.8 | 832 ± 22 |
| Km (mM) | 1.3 ± 0.1 | 1.7 ± 0.1 | 5.0 ± 0.1 | |
| kcat (s−1) | 2.6 ± 0.05 | 3.8 ± 0.02 | 2.1 ± 0.06 | |
| kcat/Km | 2.07 | 2.16 | 0.43 | |
| PsAlg7C | SA (U mg−1) | 2889 ± 31.1 | 4481 ± 47 | 3267 ± 39 |
| Km (mM) | 1.8 ± 0.1 | 2.4 ± 0.1 | 22.0 ± 0.1 | |
| kcat (s−1) | 7.3 ± 0.079 | 11 ± 0.12 | 8.3 ± 0.10 | |
| kcat/Km | 4.1 | 4.7 | 0.4 | |
| PsMan8A | SA (U mg−1) | nd. | 1093 ± 17 | nd. |
| Km (mM) | nd. | 21.0 ± 0.7 | nd. | |
| kcat (s−1) | nd. | 9.7 ± 0.2 | nd. | |
| kcat/Km | nd. | 0.5 | nd. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pilgaard, B.; Vuillemin, M.; Holck, J.; Wilkens, C.; Meyer, A.S. Specificities and Synergistic Actions of Novel PL8 and PL7 Alginate Lyases from the Marine Fungus Paradendryphiella salina. J. Fungi 2021, 7, 80. https://doi.org/10.3390/jof7020080
Pilgaard B, Vuillemin M, Holck J, Wilkens C, Meyer AS. Specificities and Synergistic Actions of Novel PL8 and PL7 Alginate Lyases from the Marine Fungus Paradendryphiella salina. Journal of Fungi. 2021; 7(2):80. https://doi.org/10.3390/jof7020080
Chicago/Turabian StylePilgaard, Bo, Marlene Vuillemin, Jesper Holck, Casper Wilkens, and Anne S. Meyer. 2021. "Specificities and Synergistic Actions of Novel PL8 and PL7 Alginate Lyases from the Marine Fungus Paradendryphiella salina" Journal of Fungi 7, no. 2: 80. https://doi.org/10.3390/jof7020080