
Effects of the Fungal Bioherbicide, Alternaria cassia on Peroxidase, Pectinolytic and Proteolytic Activities in Sicklepod Seedlings

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sources of Plant Tissue and Production of A. cassiae Spores
2.2. Extraction and Enzyme Assays
3. Results
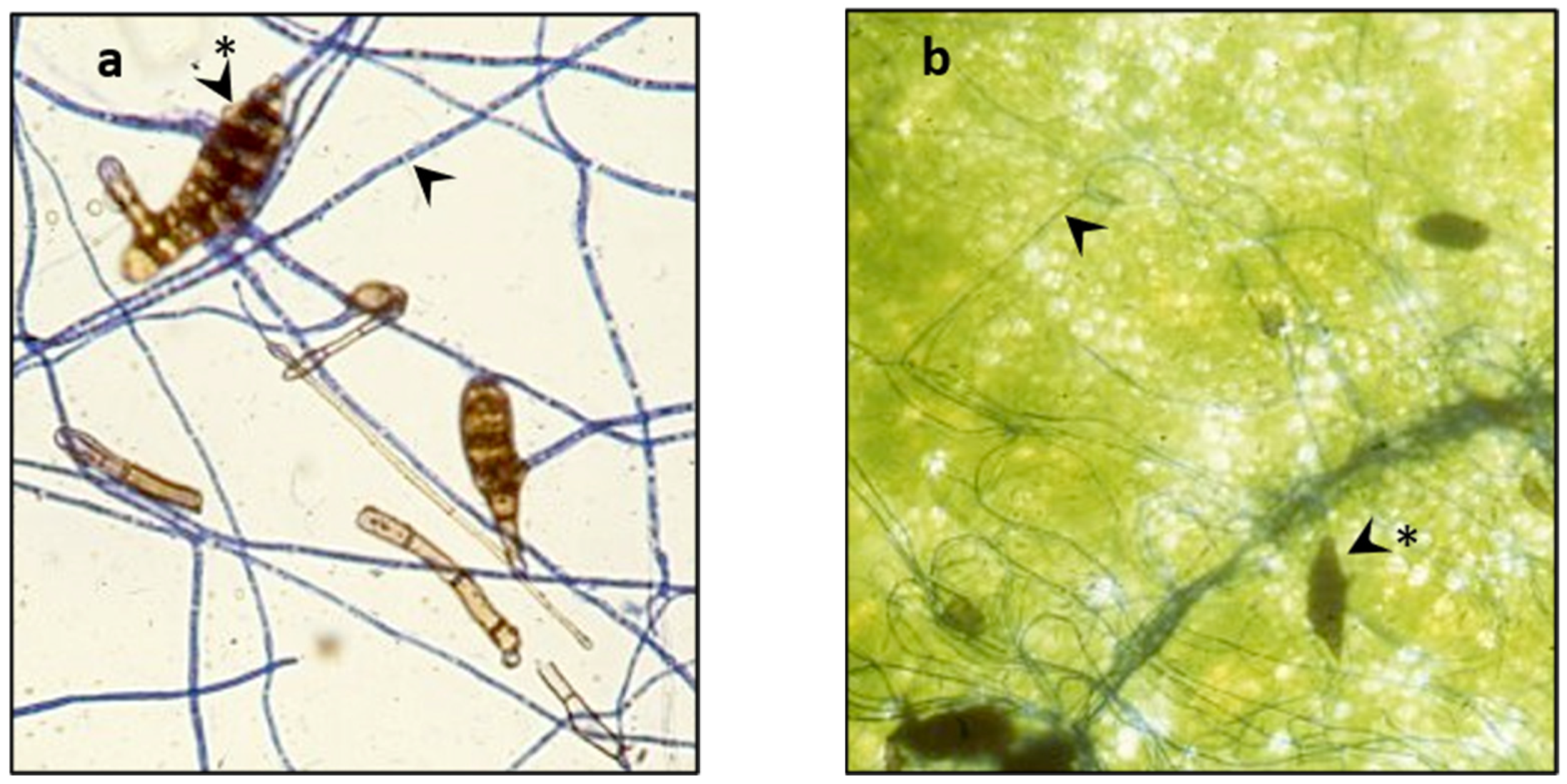
3.1. Germination, Growth, and Infection of A. cassia Spores
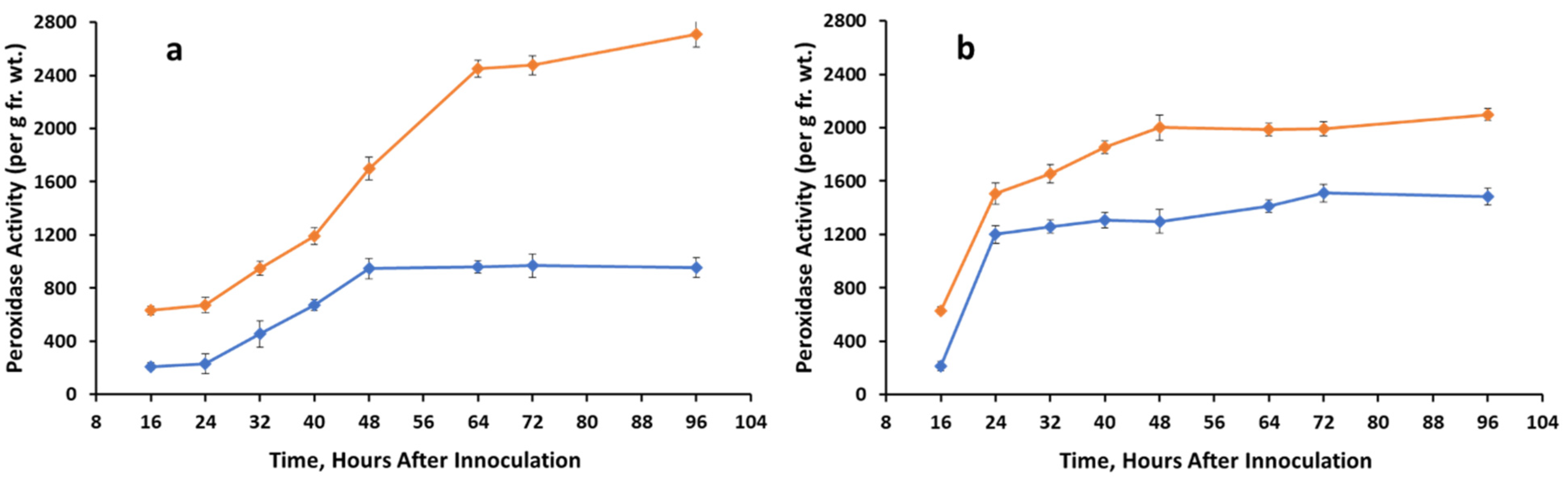
3.2. Peroxidase Activity
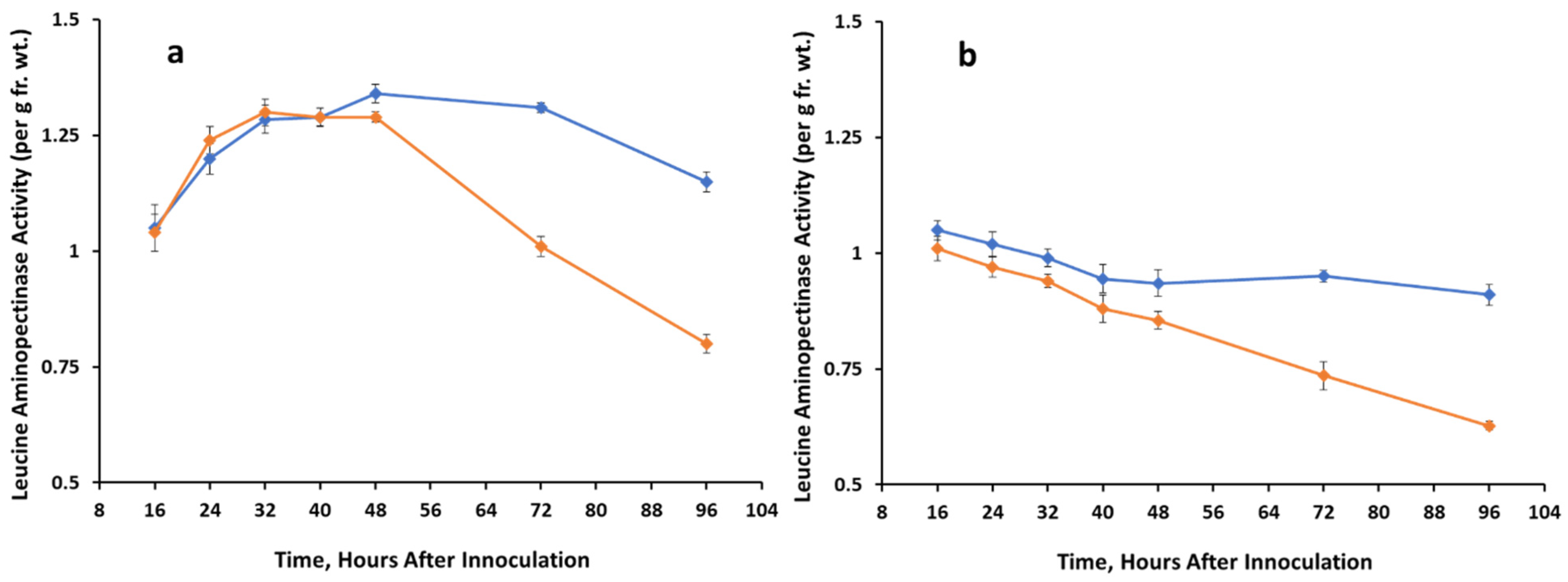
3.3. Proteolytic Activity of Two Substrates
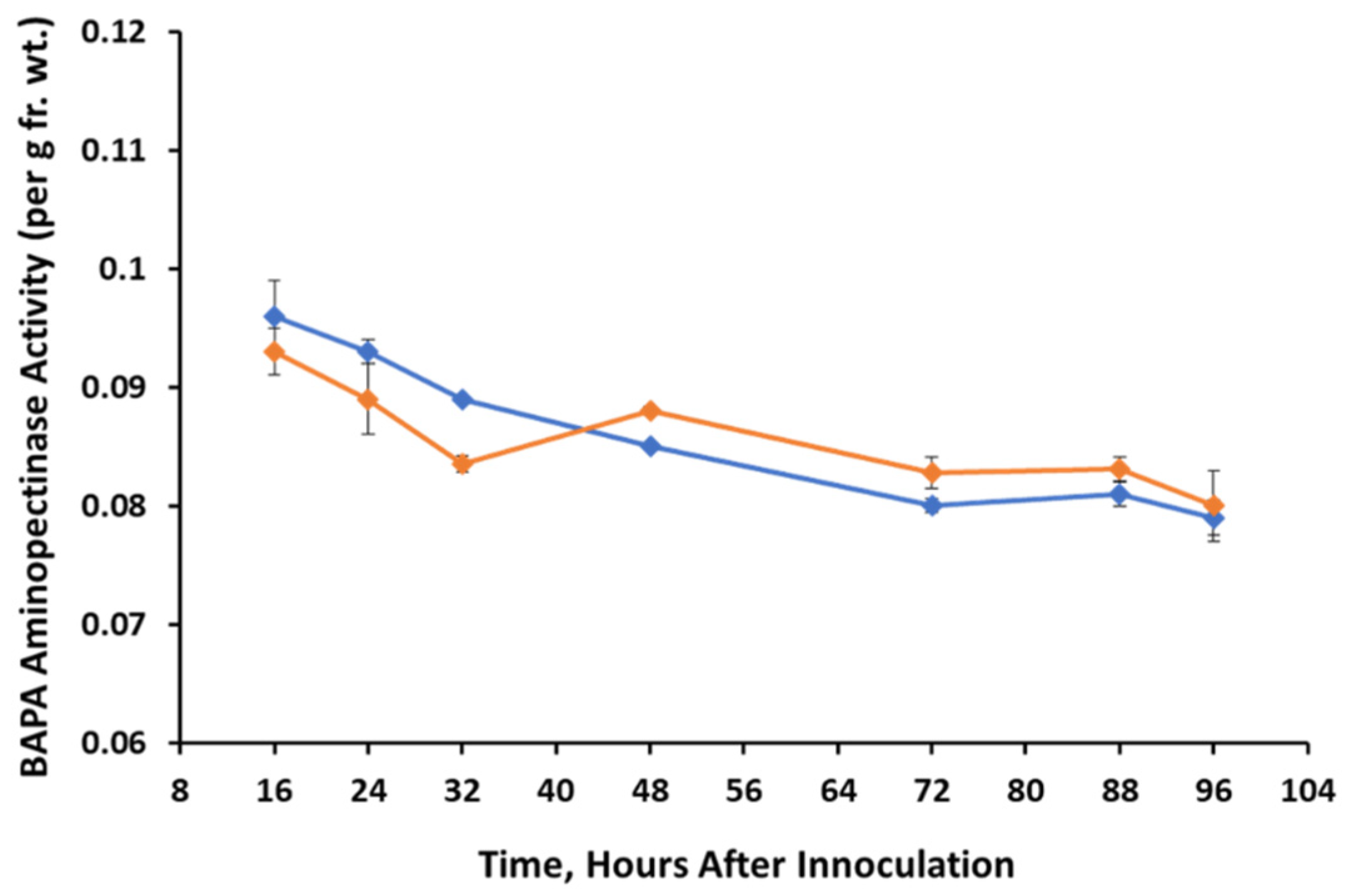
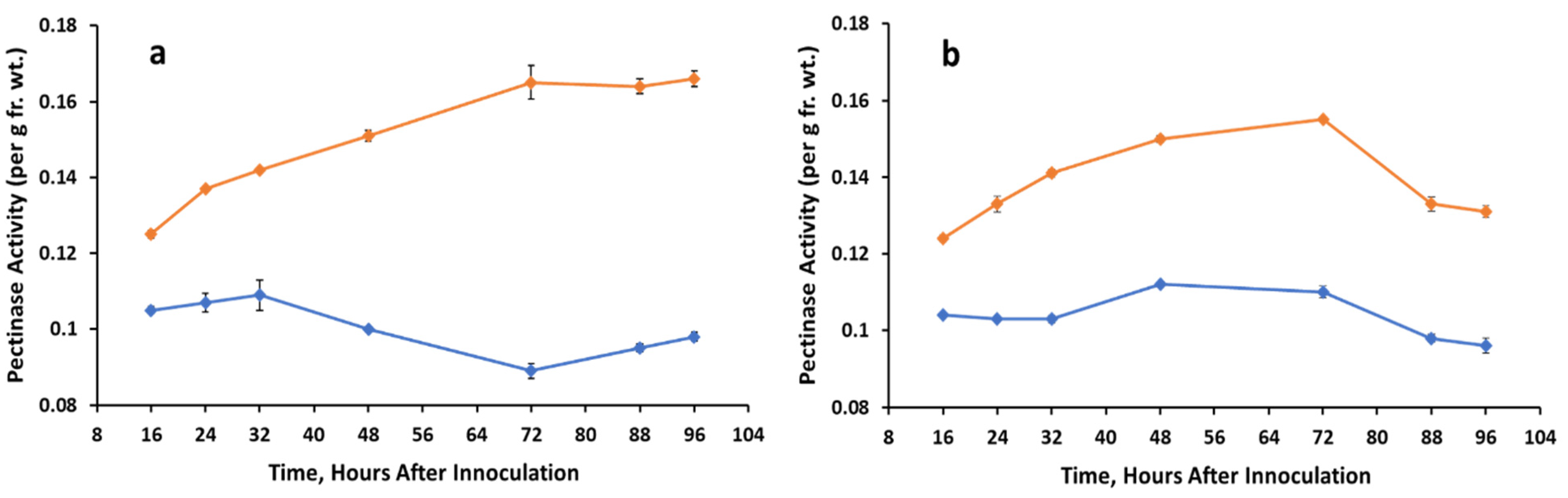
3.4. Pectinolytic Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoagland, R.E. Microbes and Microbial Products as Herbicides; American Chemical Society Symposium Series No. 439; ACS Books: Washington, DC, USA, 1990. [Google Scholar]
- TeBeest, D.O. Microbial Control of Weeds; Chapman and Hall: New York, NY, USA, 1991. [Google Scholar]
- Rosskopf, E. Use of plant pathogens in weed control. In Handbook of Weed Control; Academic Press: New York, NY, USA, 1999; pp. 891–911. [Google Scholar]
- Charudattan, R. Biological control of weeds by means of plant pathogens: Significance for integrated weed management in modern agro-ecology. BioControl 2001, 46, 229–260. [Google Scholar] [CrossRef]
- Weaver, M.A.; Lyn, M.E.; Boyette, C.D.; Hoagland, R.E. Bioherbicides for weed control. In Non-Chemical Weed Management; Upadhyaya, M.K., Blackshaw, R.E., Eds.; CAB International: New York, NY, USA, 2007; pp. 93–110. [Google Scholar]
- Boyetchko, S.M.; Rosskopf, E.N.; Caesar, A.J.; Charudattan, R. Biological weed control with pathogens: Search for candidates to applications. In Applied Mycology and Biotechnology; Khachatourians, G.G., Arora, D.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; Volume 2, pp. 239–274. [Google Scholar]
- Hoagland, R.E. Microbial allelochemicals and pathogens as bioherbicidal agents. Weed Technol. 2001, 15, 835–857. [Google Scholar] [CrossRef]
- Boyetchko, S.; Peng, G. Challenges and strategies for development of mycoherbicides. In Fungal Biotechnology in Agricultural, Food, and Environmental Applications; Arora, D.K., Bridge, P., Bhatnagar, D., Eds.; Marcel Dekker Inc.: New York, NY, USA, 2004; Volume 21, pp. 111–121. [Google Scholar]
- Duke, S.O.; Scheffler, B.E.; Boyette, C.D.; Dayan, F.E. Biotechnology in weed control. In Encyclopedia of Chemical Technology; Kirk-Othmer, Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2015; pp. 1–25. [Google Scholar]
- Hoagland, R.E.; Boyette, C.D. Controlling herbicide-susceptible, -tolerant and -resistant weeds with microbial bioherbicides. Outlooks Pest Manag. 2016, 27, 256–266. [Google Scholar] [CrossRef]
- Harding, D.P.; Raizada, M.N. Controlling weeds with fungi, bacteria and viruses: A review. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Morin, L. Progress in biological control of weeds with plant pathogens. Annu. Rev. Phytopathol. 2020, 58, 201–223. [Google Scholar] [CrossRef]
- Dalinova, A.A.; Salimova, D.R.; Berestetskiy, A.O. Fungi of the genera Alternaria as producers of biological compounds and mycoherbicides. Appl. Biochem. Microbiol. 2020, 56, 256–272. [Google Scholar] [CrossRef]
- Prasannath, K. Plant defense-related enzymes against pathogens: A review. Agrieast J. Agric. Sci. 2017, 11, 38–48. [Google Scholar] [CrossRef]
- Motlagh, M.R.S. Evaluation of Alternaria alternata causing leaf spot of barnyardgrass grown in rice fields. Afr. J. Microbiol. Res. 2012, 6, 4481–4488. [Google Scholar]
- Ghorbani, R.; Seel, W.; Litterick, A.; Leifert, C. Evaluation of Alternaria alternata for biological control of Amaranthus retroflexus. Weed Sci. 2000, 48, 474–480. [Google Scholar] [CrossRef]
- Sanyal, D.; Bhowmik, P.C.; Abbas, H.K. Effect of surfactants on bioherbicidal activity of Alternaria helianthi on multiple-seeded cocklebur. J. Plant Pathol. 2008, 1, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Masangkay, R.F.; Mabbayad, M.O.; Poulitz, T.C.; Watson, A.K. Host range of Alternaria alternata f.sp. sphenocleae causing leaf blight Sphenoclea zeylanica. Can. J. Bot. 1999, 77, 103–112. [Google Scholar]
- Shabanna, Y.M.; Charudattan, R.; Elwakil, M.A. Identification, pathogenicity and safety of Alternaria eichorniae from egypt as a bioherbicide agent for water hyacinth. Biol. Control 1995, 5, 123–135. [Google Scholar] [CrossRef]
- Walker, H.L. Control of Sicklepod, Showy Crotolaria and Coffee Senna with a Fungal Pathogen. U.S. Patent 4390360, 28 June 1983. [Google Scholar]
- Walker, H.L.; Riley, J.A. Evaluation of Alternaria cassia for the biocontrol of sicklepod (Cassia obtusifolia). Weed Sci. 1982, 30, 651–654. [Google Scholar] [CrossRef]
- Walker, H.L. A seedling blight of sicklepod caused by Alternaria cassiae. Plant Dis. 1982, 66, 426–428. [Google Scholar] [CrossRef]
- Walker, H.L.; Boyette, C.D. Biocontrol of sicklepod (Cassia obtusifolia) in soybeans (Glycine max) with Alternaria cassia. Weed Sci. 1985, 33, 212–215. [Google Scholar] [CrossRef]
- Hoagland, R.E. The mycoherbicide Alternaria cassiae infects and alters phenolic metabolism of Cassia alata Seedlings. Phytoprotection 1995, 76, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, R.E. Alternaria cassia alters phenylpropanoid metabolism in sicklepod (Cassia obtusifolia). J. Phytopathol. 1990, 130, 177–187. [Google Scholar] [CrossRef]
- Tervet, I.W.; Cherry, E. A simple device for collection of fungus spores. Plant Dis. Rep. 1950, 34, 238. [Google Scholar]
- Hoagland, R.E. Hydroponic seedling bioassay for the bioherbicides Colletotrichum truncatum and Alternaria cassiae. Biocontrol Sci. Technol. 1995, 5, 251–259. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidase. Meth. Enzymol. 1955, 2, 773–775. [Google Scholar]
- Erlanger, B.F.; Kokowsky, N.; Cohen, W. The preparation and properties of two new chromogenic substrates of trypsin. Arch. Biochem. Biophys. 1961, 96, 271–278. [Google Scholar] [CrossRef]
- Hagerman, A.E.; Austin, P.J. Continuous spectrophotometric assay for plant pectin methyl esterase. J. Agric. Food Chem. 1986, 34, 440–444. [Google Scholar] [CrossRef]
- Almagro, L.; Gomez Ros, L.V.; Belchi- Navarro, S.; Bru, R.; Ros Barcelo, A.; Pedreno, M.A. Class III peroxidases in plant defence reactions. J. Exper. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, C.; Montillet, J.L.; Bresson, E.; Agnel, J.P.; Dai, G.H.; Daniel, J.F.; Geiger, J.P.; Nicole, M. Apoplastic peroxidase generates superoxide anions in cells of cotton cotyledons undergoing the hypersensitive reaction to Xanthomonas campestris pv. malvacearum race. Mol. Plant Microbe Interact. 1998, 11, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Bindschedler, L.V.; Dewdney, J.; Blee, K.A.; Stone, J.M.; Asai, T.; Plotnikov, J.; Denoux, C.; Hayes, T.; Gerrish, C.; Davies, D.R.; et al. Peroxidase-dependent apoplastic oxidative burst in Arabidopsis required for pathogen resistance. Plant J. Cell Molec. Biol. 2006, 47, 851–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a swiss army knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Iwai, T.; Hiraga, S. Ten rice peroxidases redundantly respond to multiple stresses including infection with rice blast fungus. Plant Cell Physiol. 2004, 45, 1442–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tudzynski, B. Fungal phytohormones in pathogenic and mutualistic associations. In The Mycota; Caroll, G.C., Tudzynski, P., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1997; pp. 167–184. [Google Scholar]
- Barrett, A.J. Classification of peptidases. Methods Enzymol. 1994, 244, 1–15. [Google Scholar] [PubMed]
- Jorda, L.; Coego, A.; Conejero, V.; Vera, P. A genomic cluster containing four differentially regulated subtilisin-like processing protease genes is in tomato plants. J. Biol. Chem. 1999, 274, 2360–2365. [Google Scholar] [CrossRef] [Green Version]
- Joosten, M.H.A.J.; De Wit, P.J.G.M. The tomato–Cladosporium fulvum interaction: A versatile experimental system to study plant–pathogen interactions. Annu. Rev. Phytopathol. 1999, 37, 335–367. [Google Scholar] [CrossRef]
- Devoto, A.; Turner, J.G. Regulation of jasmonate-mediated plant responses in arabidopsis. Ann Bot. 2003, 92, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y. Proteases in pathogenesis and plant defence. Cellular Microbiol. 2004, 6, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Huitema, E.; Da Cunha, L.; Torto-Alalibo, T.; Kamoun, S. A kazal-like extracellular serine protease inhibitor from Phytophthora infestans targets the tomato pathogenesis-related protease P69B. J. Biol. Chem. 2004, 279, 26370–26377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mata, R.; Gamboa, A.; Macias, M.; Santillan, S.; Ulloa, M.; Gonzalez, C. Effects of selected phytotoxins from Guanomyces polythrix on the calmodulin-dependent activity of the enzymes cAMP phosphodiesterase and NAD-kinase. J. Agric. Food Chem. 2003, 51, 4559–4562. [Google Scholar] [CrossRef] [PubMed]
- Prade, R.A.; Zhan, D.; Ayoubi, P.; Mort, A.J. Pectins, pectinases and plant-microbe interactions. Biotechnol. Genet. Eng. Rev. 1999, 16, 361–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.; Dong, H.; Xu, C.; Liu, J.; Hu, B.; Ye, J.; Mai, G.; Li, H. Pectin methylesterases contribute the pathogenic differences between races 1 and 4 of Fusarium oxysporum f. sp. Cubense. Sci. Rep. 2017, 7, 13140. [Google Scholar] [CrossRef] [PubMed] [Green Version]


 Control,
Control,  A. cassiae.
Control, A. cassiae.
A. cassiae.
Control, A. cassiae.
 Control,
Control,  A. cassiae.
Control, A. cassiae.
A. cassiae.
Control, A. cassiae.
 Control,
Control,  A. cassiae.
Control, A. cassiae.
A. cassiae.
Control, A. cassiae.
 Control,
Control,  A. cassiae.
Control, A. cassiae.
A. cassiae.
Control, A. cassiae.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoagland, R.E.; Boyette, C.D. Effects of the Fungal Bioherbicide, Alternaria cassia on Peroxidase, Pectinolytic and Proteolytic Activities in Sicklepod Seedlings. J. Fungi 2021, 7, 1032. https://doi.org/10.3390/jof7121032
Hoagland RE, Boyette CD. Effects of the Fungal Bioherbicide, Alternaria cassia on Peroxidase, Pectinolytic and Proteolytic Activities in Sicklepod Seedlings. Journal of Fungi. 2021; 7(12):1032. https://doi.org/10.3390/jof7121032
Chicago/Turabian StyleHoagland, Robert E., and Clyde Douglas Boyette. 2021. "Effects of the Fungal Bioherbicide, Alternaria cassia on Peroxidase, Pectinolytic and Proteolytic Activities in Sicklepod Seedlings" Journal of Fungi 7, no. 12: 1032. https://doi.org/10.3390/jof7121032