
Mycorrhizal Fungal Partners Remain Constant during a Root Lifecycle of Pleione bulbocodioides (Orchidaceae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Sequencings
2.2. Data Analyses
3. Results
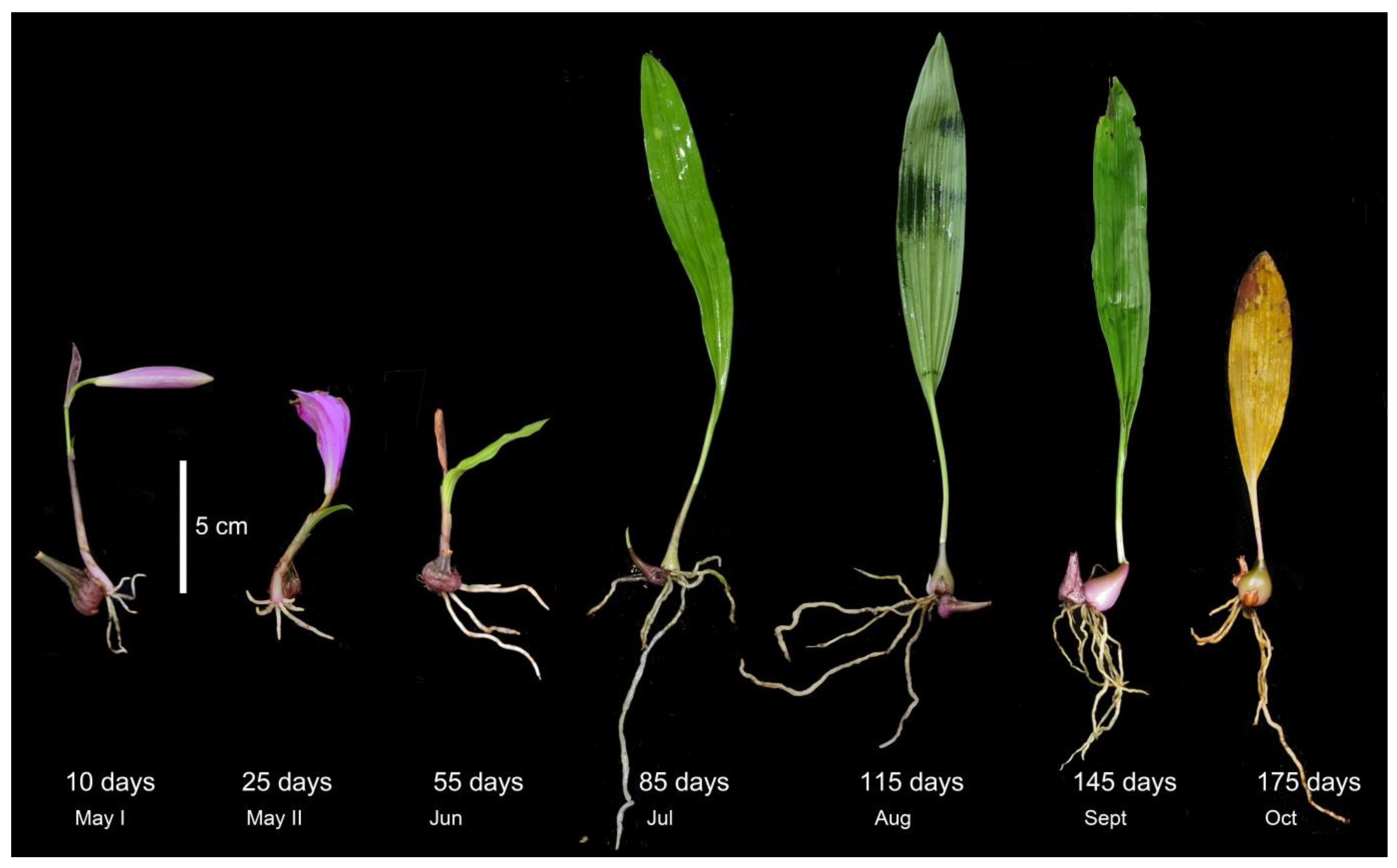
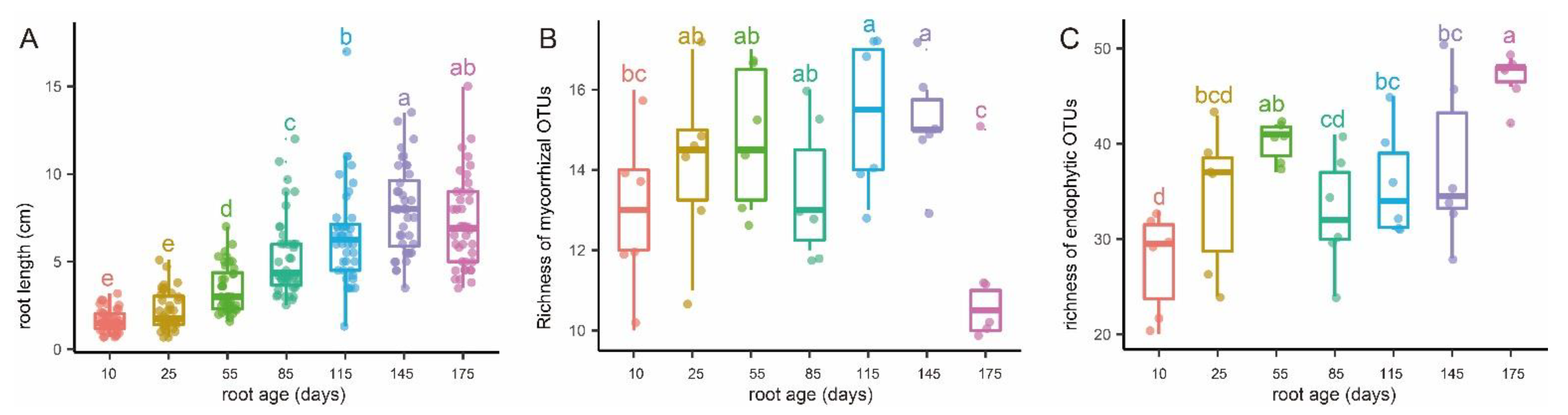
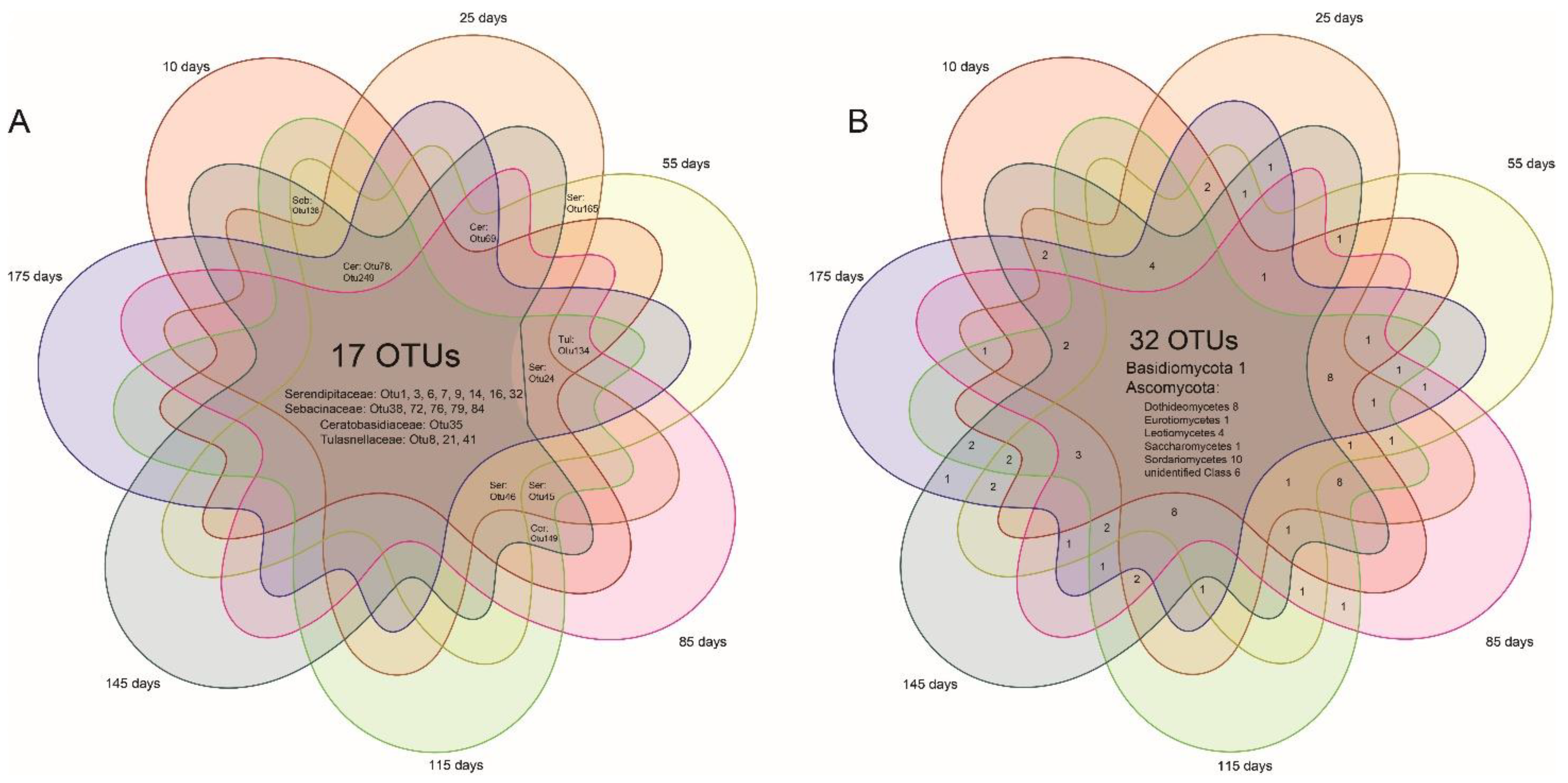
3.1. Root Phenology and the Variation of Mycorrhizal and Endophytic Fungal Richness with Root Age
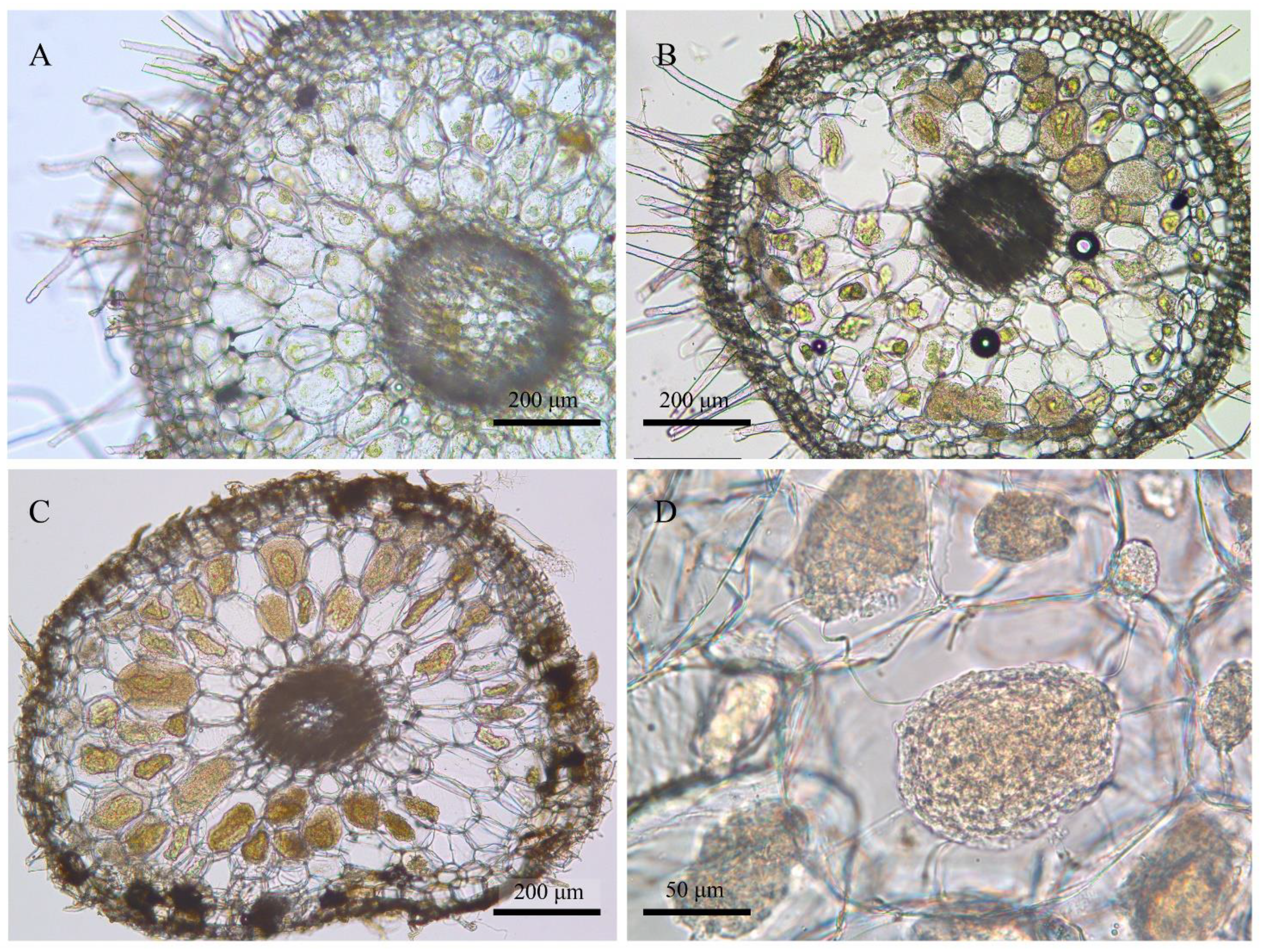
3.2. Community Compositions and the Phylogenetic Positions of Mycorrhizal Fungi
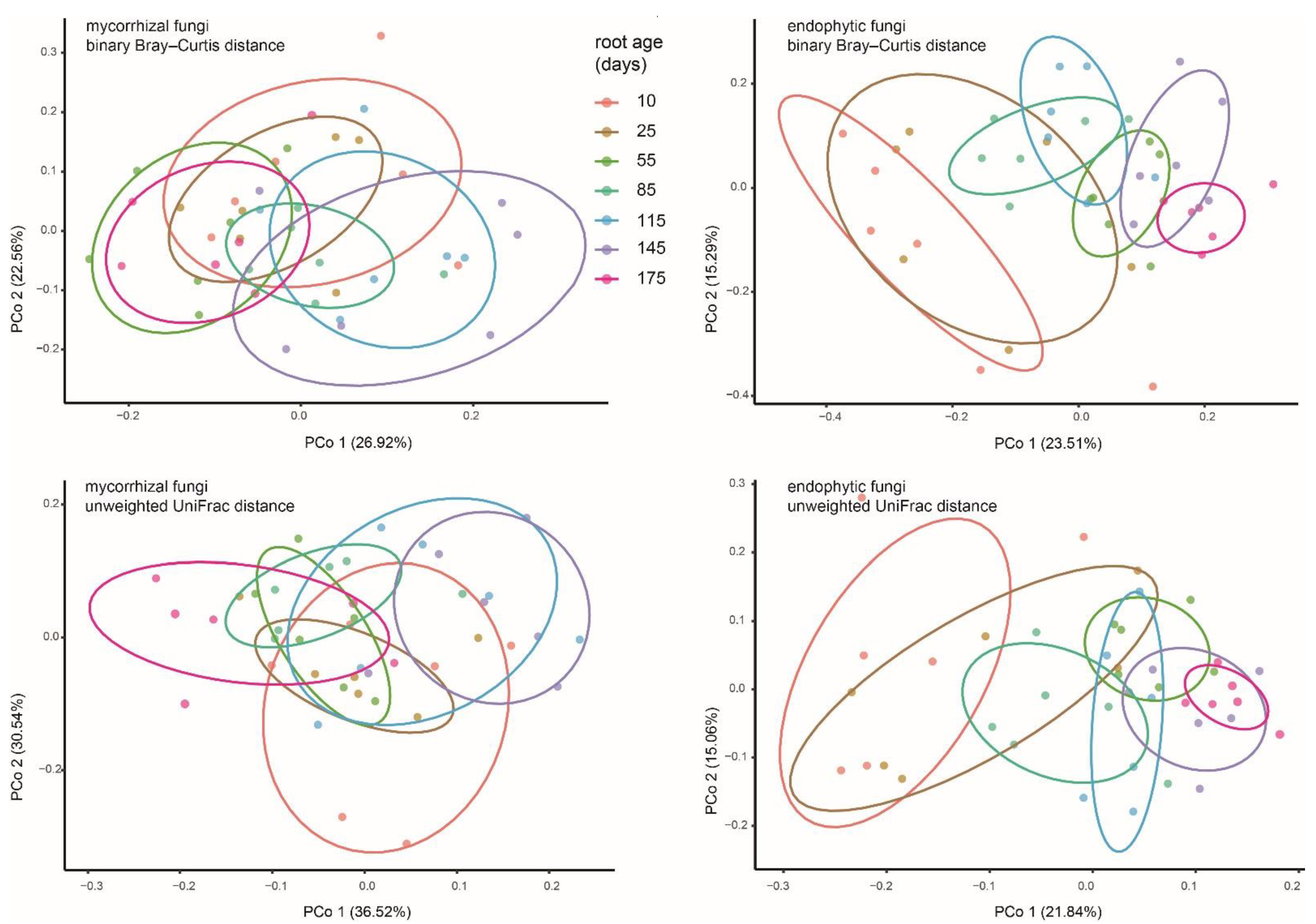
3.3. The Scenarios of Mycorrhizal and Endophytic Fungal Temporal Turnover of P. bulbocodioides
4. Discussion
4.1. Different Variation Trends in Mycorrhizal and Endophytic Fungal Richness
4.2. The Relative Constant OMF Assemblages in P. bulbocodioides
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tedersoo, L.; Bahram, M.; Zobel, M. How mycorrhizal associations drive plant population and community biology. Science 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.N. The Geographic Mosaic of Coevolution; University of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]
- Lespiaucq, A.V.; Jacquemyn, H.; Rasmussen, H.N.; Méndez, M. Temporal turnover in mycorrhizal interactions: A proof of concept with orchids. New Phytol. 2021, 230, 1690–1699. [Google Scholar] [CrossRef] [PubMed]
- McCormick, M.K.; Whigham, D.F.; Sloan, D.; O’Malley, K.; Hodkinson, B. Orchid-fungus fidelity: A marriage meant to last? Ecology 2006, 87, 903–911. [Google Scholar] [CrossRef]
- Mandyam, K.; Jumpponen, A. Seasonal and temporal dynamics of arbuscular mycorrhizal and dark septate endophytic fungi in a tallgrass prairie ecosystem are minimally affected by nitrogen enrichment. Mycorrhiza 2008, 18, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Shimizu, S.; Mori, M.; Ito, S.; Selosse, M. Seasonal and environmental changes of mycorrhizal associations and heterotrophy levels in mixotrophic Pyrola japonica (Ericaceae) growing under different light environments. Am. J. Bot. 2012, 99, 1177–1188. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Finlay, R.; Dahlberg, A.; Stenlid, J.; Wardle, D.; Lindahl, B.D. Carbon sequestration is related to mycorrhizal fungal community shifts during long-term succession in boreal forests. New Phytol. 2015, 205, 1525–1536. [Google Scholar] [CrossRef]
- Ercole, E.; Adamo, M.; Rodda, M.; Gebauer, G.; Girlanda, M.; Perotto, S. Temporal variation in mycorrhizal diversity and carbon and nitrogen stable isotope abundance in the wintergreen meadow orchid Anacamptis morio. New Phytol. 2015, 205, 1308–1319. [Google Scholar] [CrossRef]
- Rasmussen, H.N. Terrestrial orchids: From Seed to Mycotrophic Plant; Cambridge University Press: New York, NY, USA, 1995. [Google Scholar]
- Rasmussen, H.N.; Rasmussen, F.N. Seedling mycorrhiza: A discussion of origin and evolution in Orchidaceae. Bot. J. Linn. Soc. 2014, 175, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, UK, 2008. [Google Scholar]
- Gebauer, G.; Meyer, M. 15N and 13C natural abundance of autotrophic and myco-heterotrophic orchids provides insight into nitrogen and carbon gain from fungal association. New Phytol. 2003, 160, 209–223. [Google Scholar] [CrossRef]
- Stöckel, M.; Těšitelová, T.; Jersakova, J.; Bidartondo, M.I.; Gebauer, G. Carbon and nitrogen gain during the growth of orchid seedlings in nature. New Phytol. 2014, 202, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, H.N.; Dixon, K.; Jersakova, J.; Těšitelová, T. Germination and seedling establishment in orchids: A complex of requirements. Ann. Bot. 2015, 116, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, Y.-C.; Qin, L.-Y.; He, H.-Y.; Yu, X.-L.; Yang, M.-Z.; Zhang, H.-B. Dynamics of fungal communities during Gastrodia elata growth. BMC Microbiol. 2019, 19, 158. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.-Y.; Fan, X.-L.; Zhou, L.-R.; Shao, S.-C.; Liu, Q.; Selosse, M.-A.; Gao, J.-Y. Symbiotic fungi undergo a taxonomic and functional bottleneck during orchid seeds germination: A case study on Dendrobium moniliforme. Symbiosis 2019, 79, 205–212. [Google Scholar] [CrossRef]
- Těšitelová, T.; Těšitel, J.; Jersáková, J.; Ríhová, G.; Selosse, M.A. Symbiotic germination capability of four Epipactis species (Orchidaceae) is broader than expected from adult ecology. Am. J. Bot. 2012, 99, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Bidartondo, M.I.; Read, D.J. Fungal specificity bottlenecks during orchid germination and development. Mol. Ecol. 2008, 17, 3707–3716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, M.; Burnett, R.; Whigham, D. Protocorm-supporting fungi are retained in roots of mature Tipularia discolor orchids as mycorrhizal fungal diversity increases. Plants 2021, 10, 1251. [Google Scholar] [CrossRef]
- Deveautour, C.; Power, S.A.; Barnett, K.L.; Ochoa-Hueso, R.; Donn, S.; Bennett, A.E.; Powell, J. Temporal dynamics of mycorrhizal fungal communities and co-associations with grassland plant communities following experimental manipulation of rainfall. J. Ecol. 2020, 108, 515–527. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Voříšková, J.; Brabcova, V.; Cajthaml, T.; Baldrian, P. Seasonal dynamics of fungal communities in a temperate oak forest soil. New Phytol. 2014, 201, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y.; Xiao, H.; Gao, J. Seasonal dynamics of mycorrhizal fungi in Paphiopedilum spicerianum (Rchb. f) Pfitzer—A critically endangered orchid from China. Glob. Ecol. Conserv. 2016, 6, 327–338. [Google Scholar] [CrossRef]
- Calevo, J.; Voyron, S.; Adamo, M.; Alibrandi, P.; Perotto, S.; Girlanda, M. Can orchid mycorrhizal fungi be persistently harbored by the plant host? Fungal Ecol. 2021, 53, 101071. [Google Scholar] [CrossRef]
- Oja, J.; Kohout, P.; Tedersoo, L.; Kull, T.; Kõljalg, U. Temporal patterns of orchid mycorrhizal fungi in meadows and forests as revealed by 454 pyrosequencing. New Phytol. 2015, 205, 1608–1618. [Google Scholar] [CrossRef] [PubMed]
- Zotz, G. The systematic distribution of vascular epiphytes—A critical update. Bot. J. Linn. Soc. 2013, 171, 453–481. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, V.; Cruz-Blasi, J.; Damon, A.A.; Mora, J.V. Seasonality and mycorrhizal colonization in three species of epiphytic orchids in southeast Mexico. Acta Bot. Bras. 2014, 28, 512–518. [Google Scholar] [CrossRef]
- Illyes, Z.; Halasz, K.; Rudnoy, S.; Ouanphanivanh, N.; Garay, T.; Bratek, Z. Changes in the diversity of the mycorrhizal fungi of orchids as a function of the water supply of the habitat. J. Appl. Bot. Food Qual. 2009, 83, 28–36. [Google Scholar]
- Pecoraro, L.; Rasmussen, H.N.; Gomes, S.I.F.; Wang, X.; Merckx, V.S.F.T.; Cai, L.; Rasmussen, F.N. Fungal diversity driven by bark features affects phorophyte preference in epiphytic orchids from southern China. Sci. Rep. 2021, 11, 11287. [Google Scholar] [CrossRef]
- Zarate-García, A.M.; Noguera-Savelli, E.; Andrade-Canto, S.B.; Zavaleta-Mancera, H.A.; Gauthier, A.; Alatorre-Cobos, F. Bark water storage capacity influences epiphytic orchid preference for host trees. Am. J. Bot. 2020, 107, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-Q.; Li, W.; Chen, J.-X.; Zhai, J.-W.; Xu, H.-Y.; Ni, L.; Wu, S.-S. Chemical constituents and biological activity profiles on Pleione (Orchidaceae). Molecules 2019, 24, 3195. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhang, L.; Deng, Y.; Qin, J.; Zhang, S.; Hu, J. Chemical constituents of species in the genus Pleione (Orchidaceae) and the implications from molecular phylogeny. Chem. Biodivers. 2021, 18. [Google Scholar] [CrossRef] [PubMed]
- Turenne, C.Y.; Sanche, S.E.; Hoban, D.J.; Karlowsky, J.A.; Kabani, A.M. Rapid identification of fungi by using the ITS2 genetic region and an automated fluorescent capillary electrophoresis system. J. Clin. Microbiol. 1999, 37, 1846–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Waud, M.; Busschaert, P.; Lievens, B.; Jacquemyn, H. Specificity and localised distribution of mycorrhizal fungi in the soil may contribute to co-existence of orchid species. Fungal Ecol. 2016, 20, 155–165. [Google Scholar] [CrossRef]
- Cevallos, S.; Sanchez-Rodriguez, A.; Decock, C.; Declerck, S.; Suarez, J.P. Are there keystone mycorrhizal fungi associated to tropical epiphytic orchids? Mycorrhiza 2017, 27, 225–232. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Brys, R.; Waud, M.; Evans, A.; Figura, T.; Selosse, M.-A. Mycorrhizal communities and isotope signatures in two partially mycoheterotrophic orchids. Front. Plant Sci. 2021, 12, 618140. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17. [Google Scholar] [CrossRef]
- Edgar, R.C.; Flyvbjerg, H. Error filtering, pair assembly and error correction for next-generation sequencing reads. Bioinformatics 2015, 31, 3476–3482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Larsson, K.; Abarenkov, K.; Nilsson, R.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; et al. UNITE: A database providing web-based methods for the molecular identification of ectomycorrhizal fungi. New Phytol. 2005, 166, 1063–1068. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; I Gordon, J.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dearnaley, J.D.W.; Martos, F.; Selosse, M.-A. Orchid mycorrhizas: Molecular ecology, physiology, evolution and conservation aspects. In The Mycota Volume IX—Fungal Associations, 2nd ed.; Esser, K., Ed.; Springer: Berlin, Germany, 2012; pp. 207–230. [Google Scholar]
- Qin, J.; Zhang, W.; Ge, Z.-W.; Zhang, S.-B. Molecular identifications uncover diverse fungal symbionts of Pleione (Orchidaceae). Fungal Ecol. 2019, 37, 19–29. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.; Simpson, G.L.; Oksanen, M.J.; Suggests, M. Vegan: Community Ecology Package. R Package Version 2.5-6. 2013. Available online: http://CRAN.R-project.org/package=vegan (accessed on 1 January 2019).
- Mamakani, K.; Myrvold, W.; Ruskey, F. Generating all simple convexly-drawable polar symmetric 6-venn diagrams. In International Workshop on Combinatorial Algorithms; Iliopoulos, C.S., Smyth, W.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Busby, R.; Paschke, M.; Stromberger, M.E.; Gebhart, D.L. Seasonal variation in arbuscular mycorrhizal fungi root colonization of downy brome (Bromus tectorum), an invasive winter annual. J. Ecosys. Ecograph. 2021, S8. [Google Scholar] [CrossRef]
- Zeng, X.; Diao, H.; Ni, Z.; Shao, L.; Jiang, K.; Hu, C.; Huang, Q.; Huang, A.W. Temporal variation in community composition of root associated endophytic fungi and carbon and nitrogen stable isotope abundance in two Bletilla species (Orchidaceae). Plants 2021, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, H.; Whigham, D. Phenology of roots and mycorrhiza in orchid species differing in phototrophic strategy. New Phytol. 2002, 154, 797–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, T.T.; Lawrie, A.C.; Coates, F.; McLean, C.B. Effect of developmental stage and peloton morphology on success in isolation of mycorrhizal fungi in Caladenia formosa (Orchidaceae). Aust. J. Bot. 2004, 52, 231–241. [Google Scholar] [CrossRef]
- Roy, M.; Gonneau, C.; Rocheteau, A.; Berveiller, D.; Thomas, J.-C.; Damesin, C.; Selosse, M.-A. Why do mixotrophic plants stay green? A comparison between green and achlorophyllous orchid individuals in situ. Ecol. Monogr. 2013, 83, 95–117. [Google Scholar] [CrossRef] [Green Version]
- Taylor, D.L.; Bruns, T.D. Population, habitat and genetic correlates of mycorrhizal specialization in the ‘cheating’ orchids Corallorhiza maculata and C. mertensiana. Mol. Ecol. 1999, 8, 1719–1732. [Google Scholar] [CrossRef] [PubMed]
- Huynh, T.T.; Thomson, R.; Mclean, C.B.; Lawrie, A.C. Functional and genetic diversity of mycorrhizal fungi from single plants of Caladenia formosa (Orchidaceae). Ann. Bot. 2009, 104, 757–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohout, P.; Těšitelová, T.; Roy, M.; Vohník, M.; Jersáková, J. A diverse fungal community associated with Pseudorchis albida (Orchidaceae) roots. Fungal Ecol. 2013, 6, 50–64. [Google Scholar] [CrossRef]
- Zhang, W.; Hu, H.; Zhang, S.B. Divergent adaptive strategies by two co-occurring epiphytic orchids to water stress: Escape or avoidance? Front. Plant Sci. 2016, 7, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, C.K.Y.; Hew, C.S. Orchid pseudobulbs—‘false’ bulbs with a genuine importance in orchid growth and survival! Sci. Hortic. 2000, 83, 165–172. [Google Scholar] [CrossRef]
- Suetsugu, K.; Ohta, T.; Tayasu, I. Partial mycoheterotrophy in the leafless orchid Cymbidium macrorhizon. Am. J. Bot. 2018, 105, 1595–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera-Rus, I.; Pastor, J.E.; Juan, R. Fungal colonization associated with phenological stages of a photosynthetic terrestrial temperate orchid from the Southern Iberian Peninsula. J. Plant Res. 2020, 133, 807–825. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Toots, M.; Diédhiou, A.G.; Henkel, T.W.; Kjøller, R.; Morris, M.H.; Nara, K.; Nouhra, E.; Peay, K.G.; et al. Towards global patterns in the diversity and community structure of ectomycorrhizal fungi. Mol. Ecol. 2012, 21, 4160–4170. [Google Scholar] [CrossRef] [PubMed]
- Shefferson, R.P.; Bunch, W.; Cowden, C.C.; Lee, Y.I.; Kartzinel, T.R.; Yukawa, T.; Downing, J.; Jiang, H. Does evolutionary history determine specificity in broad ecological interactions? J. Ecol. 2019, 107, 1582–1593. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Waud, M.; Merckx, V.S.F.T.; Brys, R.; Tyteca, D.; Hedrén, M.; Lievens, B. Habitat driven variation in mycorrhizal communities in the terrestrial orchid genus Dactylorhiza. Sci. Rep. 2016, 6, 37182. [Google Scholar] [CrossRef]
- Voyron, S.; Ercole, E.; Ghignone, S.; Perotto, S.; Girlanda, M. Fine-scale spatial distribution of orchid mycorrhizal fungi in the soil of host-rich grasslands. New Phytol. 2017, 213, 1428–1439. [Google Scholar] [CrossRef]
- Wang, D.; Jacquemyn, H.; Gomes, S.I.F.; Vos, R.A.; Merckx, V.S.F.T. Symbiont switching and trophic mode shifts in Orchidaceae. New Phytol. 2021, 231, 791–800. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Weiss, M.; Kull, T.; Taylor, D.L. High specificity generally characterizes mycorrhizal association in rare lady’s slipper orchids, genus Cypripedium. Mol. Ecol. 2005, 14, 613–626. [Google Scholar] [CrossRef]
- Fochi, V.; Chitarra, W.; Kohler, A.; Voyron, S.; Singan, V.; Lindquist, E.A.; Barry, K.; Girlanda, M.; Grigoriev, I.; Martin, F.; et al. Fungal and plant gene expression in the Tulasnella calospora—Serapias vomeracea symbiosis provides clues about nitrogen pathways in orchid mycorrhizas. New Phytol. 2017, 213, 365–379. [Google Scholar] [CrossRef] [Green Version]
- Kohler, A.; Kuo, A.; Nagy, L.G.; Morin, E.; Barry, K.W.; Buscot, F.; Canbäck, B.; Choi, C.; Cichoki, N.; Clum, A.; et al. Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists. Nat. Genet. 2015, 47, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Mehra, S.; Morrison, P.D.; Coates, F.; Lawrie, A.C. Differences in carbon source utilisation by orchid mycorrhizal fungi from common and endangered species of Caladenia (Orchidaceae). Mycorrhiza 2017, 27, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Nurfadilah, S.; Swarts, N.D.; Dixon, K.; Lambers, H.; Merritt, D. Variation in nutrient-acquisition patterns by mycorrhizal fungi of rare and common orchids explains diversification in a global biodiversity hotspot. Ann. Bot. 2013, 111, 1233–1241. [Google Scholar] [CrossRef] [Green Version]
- Shao, S.-C.; Luo, Y.; Jacquemyn, H. Co-Cultures of Mycorrhizal fungi do not increase germination and seedling development in the epiphytic orchid Dendrobium nobile. Front. Plant Sci. 2020, 11, 571426. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycorrhizal Fungi | Endophytic Fungi | |||
|---|---|---|---|---|
| Explained Variations (R2) | p Value | Explained Variations (R2) | p Value | |
| 10 days/25 days | 0.24 | 0.009 ** | 0.2 | 0.002 ** |
| 25 days/55 days | 0.24 | 0.026 * | 0.26 | 0.004 ** |
| 55 days/85 days | 0.11 | 0.341 | 0.12 | 0.132 |
| 85 days/115 days | 0.07 | 0.634 | 0.06 | 0.880 |
| 115 days/145 days | 0.12 | 0.177 | 0.08 | 0.520 |
| 145 days/175 days | 0.15 | 0.117 | 0.18 | 0.014 * |
| 10 days/175 days | 0.19 | 0.057 | 0.17 | 0.003 ** |
| 25 days/175 days | 0.08 | 0.487 | 0.28 | 0.002 ** |
| Mycorrhizal Fungi | Endophytic Fungi | |
|---|---|---|
| Fidelity to one OMF | no | no |
| Fidelity to many OMF | 10 days–115 days | no |
| Partial replacement with constant richness | 10 days–55 days; 25 days–145 days; 55 days–115 days | 25 days–145 days |
| Partial replacement with loss | 10 days–25 days, –85 days, –145 days, –175 days; 25 days–85 days, –175 days; 55 days–85 days, –145 days; 85 days–175 days; 115 days–145 days, –175 days; 145 days–175 days | 25 days–55 days; 85 days–145 days; 115 days–145 days, –175 days |
| Partial replacement with gain | 25 days–55 days, –115 days; 85 days–115 days, –145 days | 10 days–25 days, –55 days, –85 days, –115 days, –145 days, –175 days; 25 days–85 days, –115 days, –175 days; 55 days–85 days, –115 days, –145 days, –175 days; 85 days–115 days, –175 days; 145 days–175 days |
| Nested OMF loss | 55 days–175 days | no |
| Nested OMF gain | no | no |
| Narrow total replacement | no | no |
| Total replacement with gain, less or constant richness | no | no |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, J.; Feng, J.-Q.; Zhang, W.; Zhang, S.-B. Mycorrhizal Fungal Partners Remain Constant during a Root Lifecycle of Pleione bulbocodioides (Orchidaceae). J. Fungi 2021, 7, 994. https://doi.org/10.3390/jof7110994
Qin J, Feng J-Q, Zhang W, Zhang S-B. Mycorrhizal Fungal Partners Remain Constant during a Root Lifecycle of Pleione bulbocodioides (Orchidaceae). Journal of Fungi. 2021; 7(11):994. https://doi.org/10.3390/jof7110994
Chicago/Turabian StyleQin, Jiao, Jing-Qiu Feng, Wei Zhang, and Shi-Bao Zhang. 2021. "Mycorrhizal Fungal Partners Remain Constant during a Root Lifecycle of Pleione bulbocodioides (Orchidaceae)" Journal of Fungi 7, no. 11: 994. https://doi.org/10.3390/jof7110994