
l-Lactic Acid Production Using Engineered Saccharomyces cerevisiae with Improved Organic Acid Tolerance

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Construction
2.2. ALE
2.3. Fermentation Studies
2.4. Genome Sequencing and Single-Nucleotide Polymorphism (SNP) Analysis
2.5. SNP Confirmation by Sanger Sequencing
2.6. Reverse Engineering
2.7. Cell growth, pH, and Extracellular Metabolite Analysis
2.8. Intracellular Metabolite Extraction and Derivatization
2.9. Intracellular Metabolite Analysis and Identification Using Gas Chromatography-Mass Spectroscopy (GC-MS)
2.10. Data and Statistical Analysis
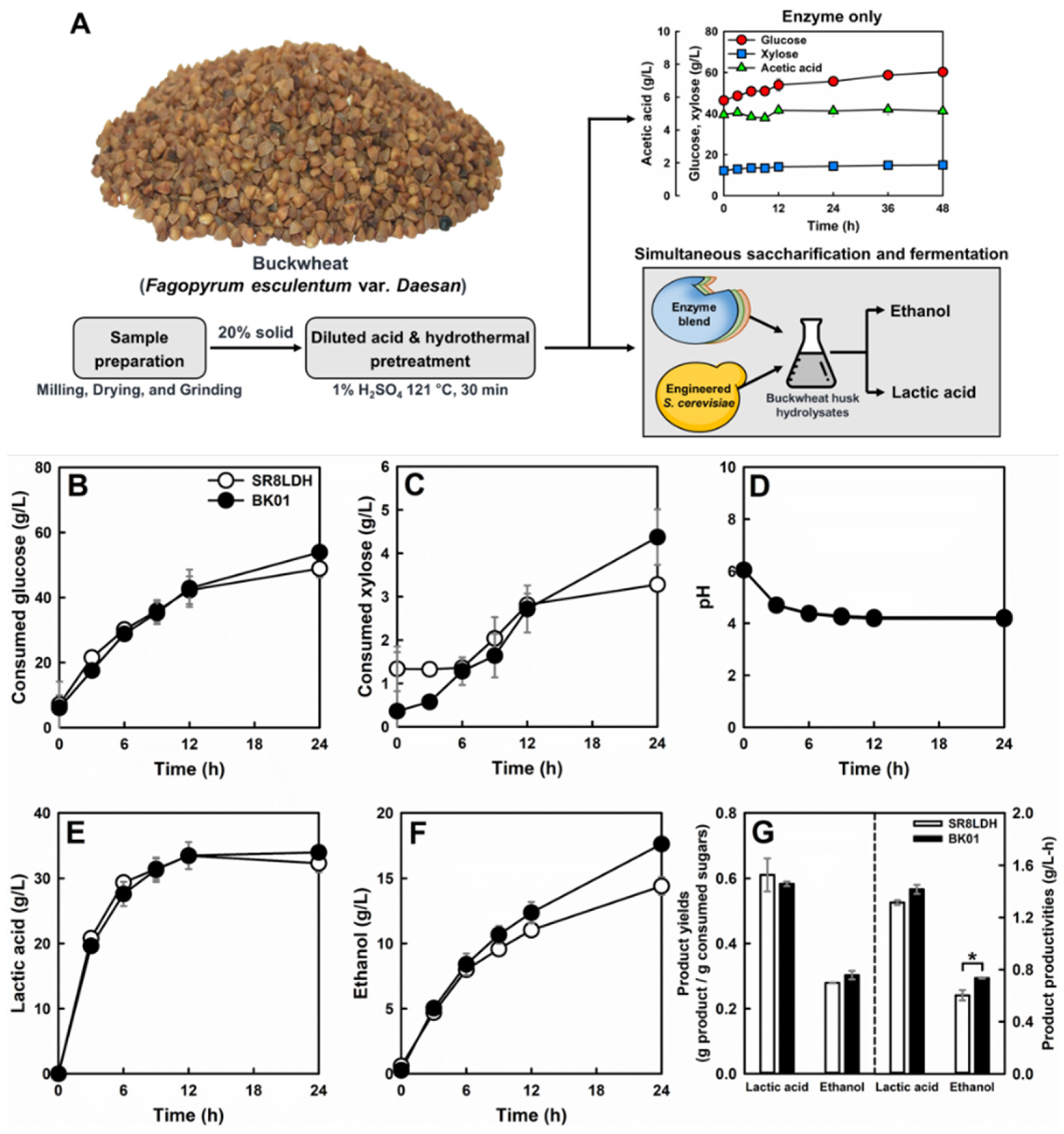
2.11. Lignocellulosic Hydrolysate Preparation and Simultaneous Saccharification and Fermentation (SSF) of the Engineered Yeast Strain
3. Results and Discussion
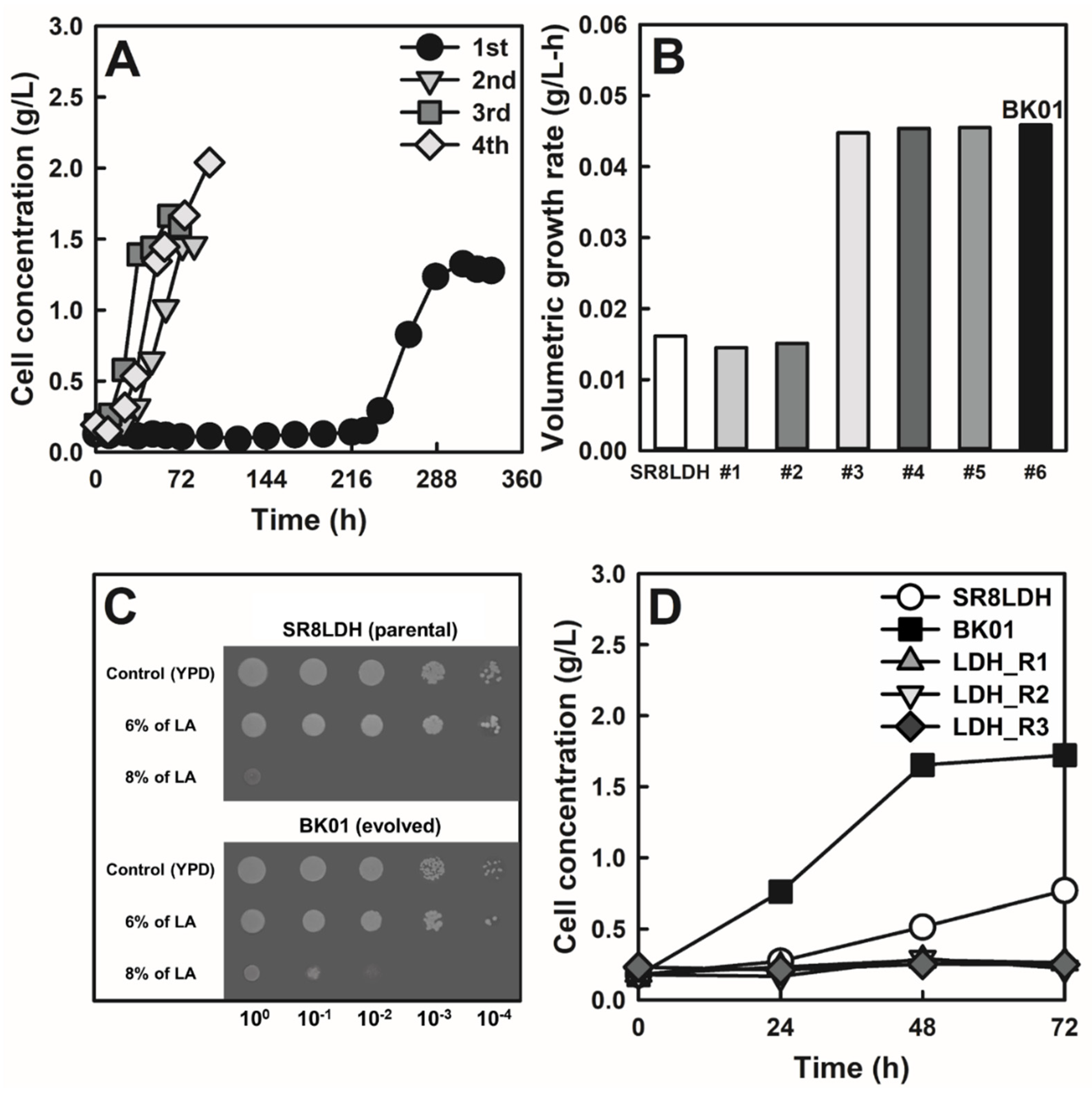
3.1. ALE of Lactic Acid-Producing S. cerevisiae in a High Concentration of Lactic Acid
3.2. Genome Sequencing of the Evolved Strain and Reverse Engineering
3.3. Metabolomic Analysis of the Evolved Strain
3.4. Lactic Acid Production by the Evolved Strain
3.5. Cellulosic Lactic Acid Production by the Evolved Strain
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Esteban, J.; Ladero, M. Food waste as a source of value-added chemicals and materials: A biorefinery perspective. Int. J. food Sci. Technol. 2018, 53, 1095–1108. [Google Scholar] [CrossRef]
- Bustamante, D.; Tortajada, M.; Ramón, D.; Rojas, A. Production of d-Lactic Acid by the Fermentation of Orange Peel Waste Hydrolysate by Lactic Acid Bacteria. Fermentation 2020, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Jamshidian, M.; Tehrany, E.A.; Imran, M.; Jacquot, M.; Desobry, S. Poly-lactic acid: Production, applications, nanocomposites, and release studies. Compr. Rev. food Sci. food Saf. 2010, 9, 552–571. [Google Scholar] [CrossRef] [PubMed]
- Singhvi, M.; Zinjarde, S.; Gokhale, D. Polylactic acid: Synthesis and biomedical applications. J. Appl. Microbiol. 2019, 127, 1612–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohn, Y.J.; Kim, H.T.; Jo, S.Y.; Song, H.M.; Baritugo, K.-A.; Pyo, J.; Choi, J.-i.; Joo, J.C.; Park, S.J. Recent Advances in Systems Metabolic Engineering Strategies for the Production of Biopolymers. Biotechnol. Bioprocess Eng. 2020, 25, 848–861. [Google Scholar] [CrossRef]
- Sauer, M.; Porro, D.; Mattanovich, D.; Branduardi, P. Microbial production of organic acids: Expanding the markets. Trends Biotechnol. 2008, 26, 100–108. [Google Scholar] [CrossRef]
- Ghaffar, T.; Irshad, M.; Anwar, Z.; Aqil, T.; Zulifqar, Z.; Tariq, A.; Kamran, M.; Ehsan, N.; Mehmood, S. Recent trends in lactic acid biotechnology: A brief review on production to purification. J. Radiat. Res. Appl. Sci. 2014, 7, 222–229. [Google Scholar] [CrossRef]
- Kim, J.-w.; Jang, J.H.; Yeo, H.J.; Seol, J.; Kim, S.R.; Jung, Y.H. Lactic acid production from a whole slurry of acid-pretreated spent coffee grounds by engineered Saccharomyces cerevisiae. Appl. Biochem. Biotechnol. 2019, 189, 206–216. [Google Scholar] [CrossRef]
- Miller, C.; Fosmer, A.; Rush, B.; McMullin, T.; Beacom, D.; Suominen, P. Industrial production of lactic acid. Compr. Biotechnol. 2011, 3, 179–188. [Google Scholar]
- Kong, X.; Zhang, B.; Hua, Y.; Zhu, Y.; Li, W.; Wang, D.; Hong, J. Efficient l-lactic acid production from corncob residue using metabolically engineered thermo-tolerant yeast. Bioresour. Technol. 2019, 273, 220–230. [Google Scholar] [CrossRef]
- Ishida, N.; Suzuki, T.; Tokuhiro, K.; Nagamori, E.; Onishi, T.; Saitoh, S.; Kitamoto, K.; Takahashi, H. d-Lactic acid production by metabolically engineered Saccharomyces cerevisiae. J. Biosci. Bioeng. 2006, 101, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Eş, I.; Mousavi Khaneghah, A.; Barba, F.J.; Saraiva, J.A.; Sant′Ana, A.S.; Hashemi, S.M.B. Recent advancements in lactic acid production—A review. Food Res. Int. 2018, 107, 763–770. [Google Scholar] [CrossRef]
- Kim, S.-K.; Auh, J.-H. Evaluating the Engineered Saccharomyces cerevisiae With High Spermidine Contents for Increased Tolerance to Lactic, Succinic, and Malic Acids and Increased Xylose Fermentation. Biotechnol. Bioprocess Eng. 2021, 26, 47–54. [Google Scholar] [CrossRef]
- Saitoh, S.; Ishida, N.; Onishi, T.; Tokuhiro, K.; Nagamori, E.; Kitamoto, K.; Takahashi, H. Genetically engineered wine yeast produces a high concentration of l-lactic acid of extremely high optical purity. Appl. Environ. Microbiol. 2005, 71, 2789–2792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Kang, C.D.; Lee, S.H.; Park, Y.K.; Cho, K.M. Engineering cellular redox balance in Saccharomyces cerevisiae for improved production of l-lactic acid. Biotechnol. Bioeng. 2015, 112, 751–758. [Google Scholar] [CrossRef]
- Yamada, R.; Wakita, K.; Mitsui, R.; Ogino, H. Enhanced d-lactic acid production by recombinant Saccharomyces cerevisiae following optimization of the global metabolic pathway. Biotechnol. Bioeng. 2017, 114, 2075–2084. [Google Scholar] [CrossRef]
- Baek, S.H.; Kwon, E.Y.; Bae, S.J.; Cho, B.R.; Kim, S.Y.; Hahn, J.S. Improvement of d-lactic acid production in Saccharomyces cerevisiae under acidic conditions by evolutionary and rational metabolic engineering. Biotechnol. J. 2017, 12, 1700015. [Google Scholar] [CrossRef]
- Baek, S.-H.; Kwon, E.Y.; Kim, Y.H.; Hahn, J.-S. Metabolic engineering and adaptive evolution for efficient production of d-lactic acid in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2016, 100, 2737–2748. [Google Scholar] [CrossRef] [PubMed]
- Mimitsuka, T.; Sawai, K.; Kobayashi, K.; Tsukada, T.; Takeuchi, N.; Yamada, K.; Ogino, H.; Yonehara, T. Production of d-lactic acid in a continuous membrane integrated fermentation reactor by genetically modified Saccharomyces cerevisiae: Enhancement in d-lactic acid carbon yield. J. Biosci. Bioeng. 2015, 119, 65–71. [Google Scholar] [CrossRef]
- Song, J.-Y.; Park, J.-S.; Kang, C.D.; Cho, H.-Y.; Yang, D.; Lee, S.; Cho, K.M. Introduction of a bacterial acetyl-CoA synthesis pathway improves lactic acid production in Saccharomyces cerevisiae. Metab. Eng. 2016, 35, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.L.; Zhang, G.-C.; Kim, S.R.; Subramaniam, V.; Steffen, D.; Skory, C.D.; Jang, J.Y.; Yu, B.J.; Jin, Y.-S. Lactic acid production from xylose by engineered Saccharomyces cerevisiae without PDC or ADH deletion. Appl. Microbiol. Biotechnol. 2015, 99, 8023–8033. [Google Scholar] [CrossRef]
- Park, H.J.; Bae, J.H.; Ko, H.J.; Lee, S.H.; Sung, B.H.; Han, J.I.; Sohn, J.H. Low-pH production of d-lactic acid using newly isolated acid tolerant yeast Pichia kudriavzevii NG7. Biotechnol. Bioeng. 2018, 115, 2232–2242. [Google Scholar] [CrossRef] [PubMed]
- Singhvi, M.; Zendo, T.; Sonomoto, K. Free lactic acid production under acidic conditions by lactic acid bacteria strains: Challenges and future prospects. Appl. Microbiol. Biotechnol. 2018, 102, 5911–5924. [Google Scholar] [CrossRef]
- Vaidya, A.; Pandey, R.; Mudliar, S.; Kumar, M.S.; Chakrabarti, T.; Devotta, S. Production and recovery of lactic acid for polylactide—an overview. Crit. Rev. Environ. Sci. Technol. 2005, 35, 429–467. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial production of lactic acid: The latest development. Crit. Rev. Biotechnol. 2016, 36, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Peetermans, A.; Foulquié-Moreno, M.R.; Thevelein, J.M. Mechanisms underlying lactic acid tolerance and its influence on lactic acid production in Saccharomyces cerevisiae. Microb Cell 2021, 8, 111–130. [Google Scholar] [CrossRef]
- Jang, B.-K.; Jeong, D.; Seol, J.; Lee, Y.-K.; Kim, S.R. Xylose Facilitates Lactic Acid Yield of Engineered Saccharomyces cerevisiae. KSBB J. 2020, 35, 129–134. [Google Scholar] [CrossRef]
- Jang, J.; Ju, Y.; Lee, Y.-K.; Seol, J.; Kim, S.R. Production of Lactic Acid by Simultaneous Saccharification and Fermentation of Sargassum horneri. KSBB J. 2021, 36, 118–122. [Google Scholar] [CrossRef]
- Kim, S.R.; Skerker, J.M.; Kang, W.; Lesmana, A.; Wei, N.; Arkin, A.P.; Jin, Y.-S. Rational and evolutionary engineering approaches uncover a small set of genetic changes efficient for rapid xylose fermentation in Saccharomyces cerevisiae. PLoS ONE 2013, 8, e57048. [Google Scholar] [CrossRef] [Green Version]
- Lane, S.; Xu, H.; Oh, E.J.; Kim, H.; Lesmana, A.; Jeong, D.; Zhang, G.; Tsai, C.-S.; Jin, Y.-S.; Kim, S.R. Glucose repression can be alleviated by reducing glucose phosphorylation rate in Saccharomyces cerevisiae. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Kim, S.; Sorek, H.; Lee, Y.; Jeong, D.; Kim, J.; Oh, E.J.; Yun, E.J.; Wemmer, D.E.; Kim, K.H. PHO13 deletion-induced transcriptional activation prevents sedoheptulose accumulation during xylose metabolism in engineered Saccharomyces cerevisiae. Metab. Eng. 2016, 34, 88–96. [Google Scholar] [CrossRef]
- Shin, M.; Kim, J.-w.; Ye, S.; Kim, S.; Jeong, D.; Lee, D.Y.; Kim, J.N.; Jin, Y.-S.; Kim, K.H.; Kim, S.R. Comparative global metabolite profiling of xylose-fermenting Saccharomyces cerevisiae SR8 and Scheffersomyces stipitis. Appl. Microbiol. Biotechnol. 2019, 103, 5435–5446. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Oh, E.J.; Ko, J.K.; Nam, J.-O.; Park, H.-S.; Jin, Y.-S.; Lee, E.J.; Kim, S.R. Metabolic engineering considerations for the heterologous expression of xylose-catabolic pathways in Saccharomyces cerevisiae. PLoS ONE 2020, 15, e0236294. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, D.Y.; Wohlgemuth, G.; Park, H.S.; Fiehn, O.; Kim, K.H. Evaluation and optimization of metabolome sample preparation methods for Saccharomyces cerevisiae. Anal. Chem. 2013, 85, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.K.; Kim, J.; Cheong, Y.E.; Kim, K.H.; Cha, H.S. Variation in the synovial fluid metabolome according to disease activity of rheumatoid arthritis. Clin. Exp. Rheumatol. 2020, 38, 500–507. [Google Scholar]
- Furukawa, K.; Toyomizu, M.; Kikusato, M. Possible role of corticosterone in proteolysis, glycolytic, and amino acid metabolism in primary cultured avian myotubes incubated at high-temperature conditions. Domest. Anim. Endocrinol. 2021, 76, 106608. [Google Scholar] [CrossRef]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Ringnér, M. What is principal component analysis? Nat. Biotechnol. 2008, 26, 303–304. [Google Scholar] [CrossRef]
- Park, H.; Park, S.U.; Jang, B.K.; Lee, J.J.; Chung, Y.S. Germplasm evaluation of Kenaf (Hibiscus cannabinus) for alternative biomass for cellulosic ethanol production. GCB Bioenergy 2021, 13, 201–210. [Google Scholar] [CrossRef]
- Abbott, D.A.; Suir, E.; van Maris, A.J.; Pronk, J.T. Physiological and transcriptional responses to high concentrations of lactic acid in anaerobic chemostat cultures of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2008, 74, 5759–5768. [Google Scholar] [CrossRef] [Green Version]
- Graves, T.; Narendranath, N.V.; Dawson, K.; Power, R. Effect of pH and lactic or acetic acid on ethanol productivity by Saccharomyces cerevisiae in corn mash. J. Ind. Microbiol. Biotechnol. 2006, 33, 469. [Google Scholar] [CrossRef]
- Mitsui, R.; Yamada, R.; Matsumoto, T.; Yoshihara, S.; Tokumoto, H.; Ogino, H. Construction of lactic acid-tolerant Saccharomyces cerevisiae by using CRISPR-Cas-mediated genome evolution for efficient d-lactic acid production. Appl. Microbiol. Biotechnol. 2020, 104, 9147–9158. [Google Scholar] [CrossRef]
- Pal, P.; Sikder, J.; Roy, S.; Giorno, L. Process intensification in lactic acid production: A review of membrane based processes. Chem. Eng. Process. Process. Intensif. 2009, 48, 1549–1559. [Google Scholar] [CrossRef]
- Kumar, A.; Thakur, A.; Panesar, P.S. Lactic acid and its separation and purification techniques: A review. Rev. Environ. Sci. Bio/Technol. 2019, 18, 823–853. [Google Scholar] [CrossRef]
- Dusselier, M.; Van Wouwe, P.; Dewaele, A.; Makshina, E.; Sels, B.F. Lactic acid as a platform chemical in the biobased economy: The role of chemocatalysis. Energy Environ. Sci. 2013, 6, 1415–1442. [Google Scholar] [CrossRef]
- Suzuki, T.; Sakamoto, T.; Sugiyama, M.; Ishida, N.; Kambe, H.; Obata, S.; Kaneko, Y.; Takahashi, H.; Harashima, S. Disruption of multiple genes whose deletion causes lactic-acid resistance improves lactic-acid resistance and productivity in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2013, 115, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Yang, M.; Hao, X.; Sharshar, M.M.; Wang, Q.; Xing, J. Improvement of d-lactic acid production at low pH through expressing acid-resistant gene IoGAS1 in engineered Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. I: Inhibition and detoxification. Bioresour. Technol. 2000, 74, 17–24. [Google Scholar] [CrossRef]
- Ko, J.K.; Enkh-Amgalan, T.; Gong, G.; Um, Y.; Lee, S.M. Improved bioconversion of lignocellulosic biomass by Saccharomyces cerevisiae engineered for tolerance to acetic acid. GCB Bioenergy 2020, 12, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Baptista, S.L.; Costa, C.E.; Cunha, J.T.; Soares, P.O.; Domingues, L. Metabolic engineering of Saccharomyces cerevisiae for the production of top value chemicals from biorefinery carbohydrates. Biotechnol. Adv. 2021, 107697. [Google Scholar] [CrossRef]
- Lam, F.H.; Turanlı-Yıldız, B.; Liu, D.; Resch, M.G.; Fink, G.R.; Stephanopoulos, G. Engineered yeast tolerance enables efficient production from toxified lignocellulosic feedstocks. Sci. Adv. 2021, 7, eabf7613. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Strain | Genotype | Reference |
|---|---|---|---|
| Background strain | SR8 | S. cerevisiae D452-2 expressing the xylose oxidoreductase pathway derived from Pichia stipitis (XYL1, XYL2, and XYL3), Δald6, adaptive laboratory evolution on xylose | [29] |
| Parental strain | SR8LDH | Xylose-metabolizing SR8 expressing the ldh gene from Lactobacillus acidophilus | [8] |
| Evolved strain | BK01 | SR8LDH strain evolved on complex medium containing 20 g/L glucose and 8% lactic acid | This study |
| BK_D1 | BK01 Δypt7 | This study | |
| BK_D2 | BK01 Δyol159c-a | This study | |
| BK_D3 | BK01 Δypt7/Δyol159c-a | This study | |
| Reverse-engineered strains | LDH_R1 | SR8LDH mYPT7 (533G>A) | This study |
| LDH_R2 | SR8LDH mYOL159C-A (172C>A) | This study | |
| LDH_R3 | SR8LDH mYPT7/mYOL159C-A | This study | |
| LDH_D1 | SR8LDH Δypt7 | This study | |
| LDH_D2 | SR8LDH Δyol159c-a | This study | |
| LDH_D3 | SR8LDH Δypt7/Δyol159c-a | This study |
| Strain | LDH Origin | ALE | pH Control | pH | Time (h) | Yield (g/g) | Titer (g/L) | Reference |
|---|---|---|---|---|---|---|---|---|
| δpHδLA2-51/dP36 | Lm | + | 20 g/L CaCO3 | 6.4 | 23 | 0.55 | 52 | [42] |
| YPH499/dPdA3-34/DLDH/1-18 | Lm | − | 40 g/L CaCO3 | ND | 216 | 0.65 | 60 | [16] |
| JHY5730 | Lm | + | 4 N NaOH | 3.5 | 55 | 0.83 | 83 | [17] |
| JHY5330 | Lm | − | 50 g/L CaCO3 | ND | 51 | 0.80 | 112 | [18] |
| SP7 | Ps | − | Continuous | 3.5–6.0 | 49 | 0.58 | 117 | [15] |
| T165R | Bt | − | Unknown | 5.2 | 48 | 0.61 | 122 | [14] |
| SP1130 | Ps, Bt | − | 5 N Ca(OH)2 | 4.7 | 40 | 0.89 | 142 | [20] |
| δpHδLA2-51/dP36 | Lm | + | None | ND | 52 | 0.30 | 34 | [42] |
| SH6779 | Bt | − | None | ND | 72 | ND | 48 | [46] |
| YIP-A15G12 | Ec | − | None | ND | 76 | 0.70 | 92 | [47] |
| SR8LDH | La | − | None | ND | 96 | 0.61 | 102 | This study |
| BK01 | La | + | None | ND | 96 | 0.72 | 119 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, B.-K.; Ju, Y.; Jeong, D.; Jung, S.-K.; Kim, C.-K.; Chung, Y.-S.; Kim, S.-R. l-Lactic Acid Production Using Engineered Saccharomyces cerevisiae with Improved Organic Acid Tolerance. J. Fungi 2021, 7, 928. https://doi.org/10.3390/jof7110928
Jang B-K, Ju Y, Jeong D, Jung S-K, Kim C-K, Chung Y-S, Kim S-R. l-Lactic Acid Production Using Engineered Saccharomyces cerevisiae with Improved Organic Acid Tolerance. Journal of Fungi. 2021; 7(11):928. https://doi.org/10.3390/jof7110928
Chicago/Turabian StyleJang, Byeong-Kwan, Yebin Ju, Deokyeol Jeong, Sung-Keun Jung, Chang-Kil Kim, Yong-Suk Chung, and Soo-Rin Kim. 2021. "l-Lactic Acid Production Using Engineered Saccharomyces cerevisiae with Improved Organic Acid Tolerance" Journal of Fungi 7, no. 11: 928. https://doi.org/10.3390/jof7110928