
Fungal Biomarkers Stability in Mars Regolith Analogues after Simulated Space and Mars-like Conditions

 ,
,  , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ground-Based Simulations
2.1.1. Science Verification Tests (SVTs)
2.1.2. Fungal Melanin Extraction
2.1.3. Spectrophotometric Analysis
2.1.4. Confocal Raman Spectroscopy Analyses
2.2. Nucleic Acid Analysis
2.2.1. Nucleic Acid Extractions from Synthetic Mars and Terrestrial Soils
2.2.2. Acid Nucleic Detection through Quantitative Real-Time PCR (qPCR)
2.2.3. Statistical Analyses
2.2.4. Organic Compounds Detection by Gas Chromatography–Mass Spectrometry
3. Results
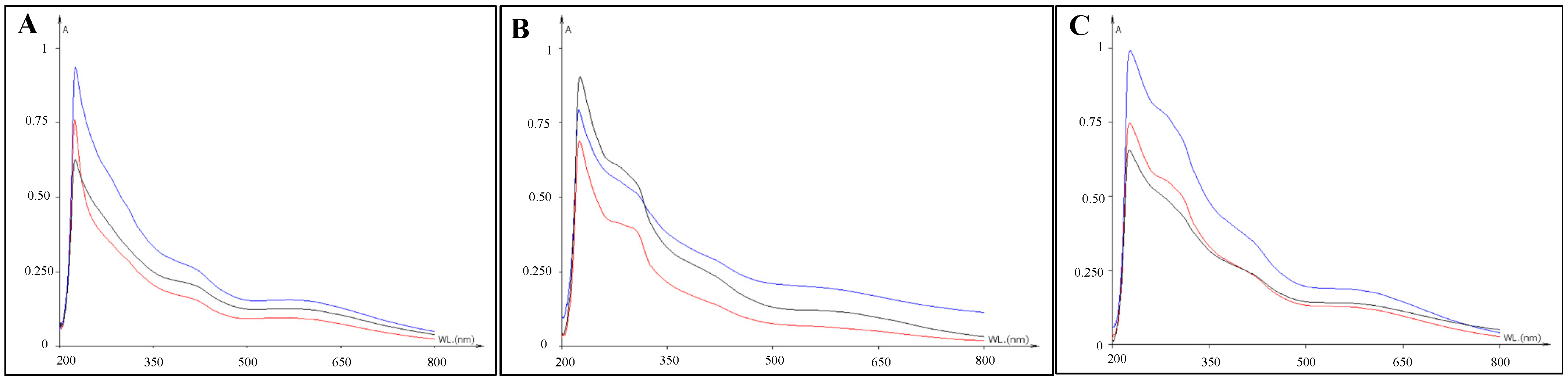
3.1. Detection of Pigments by Spectrophotometric Analyses
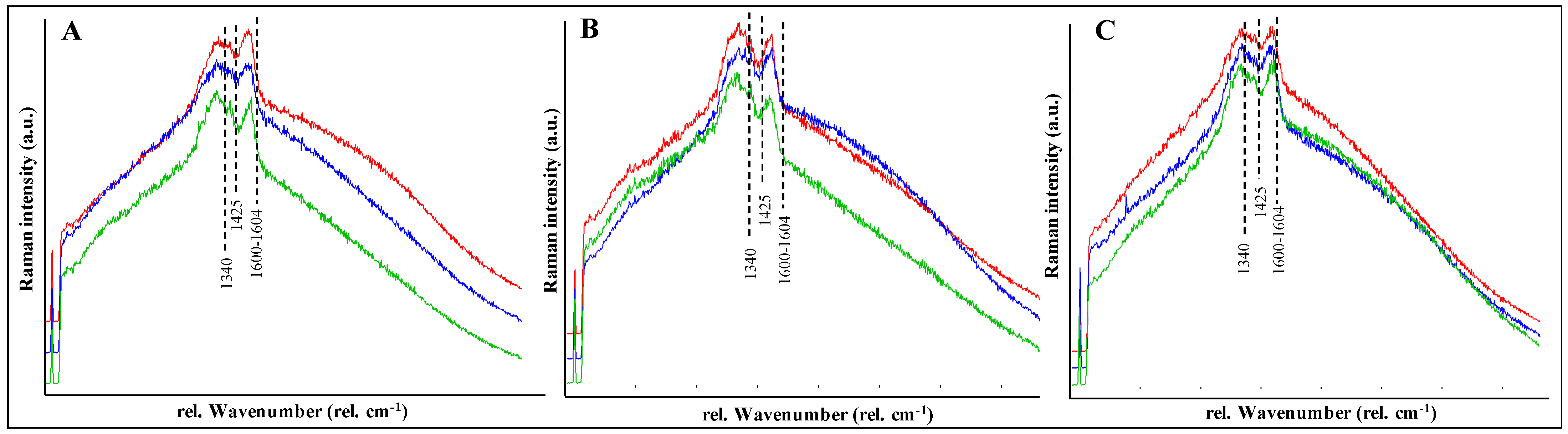
3.2. Detection of Pigments by Confocal Raman Spectroscopy
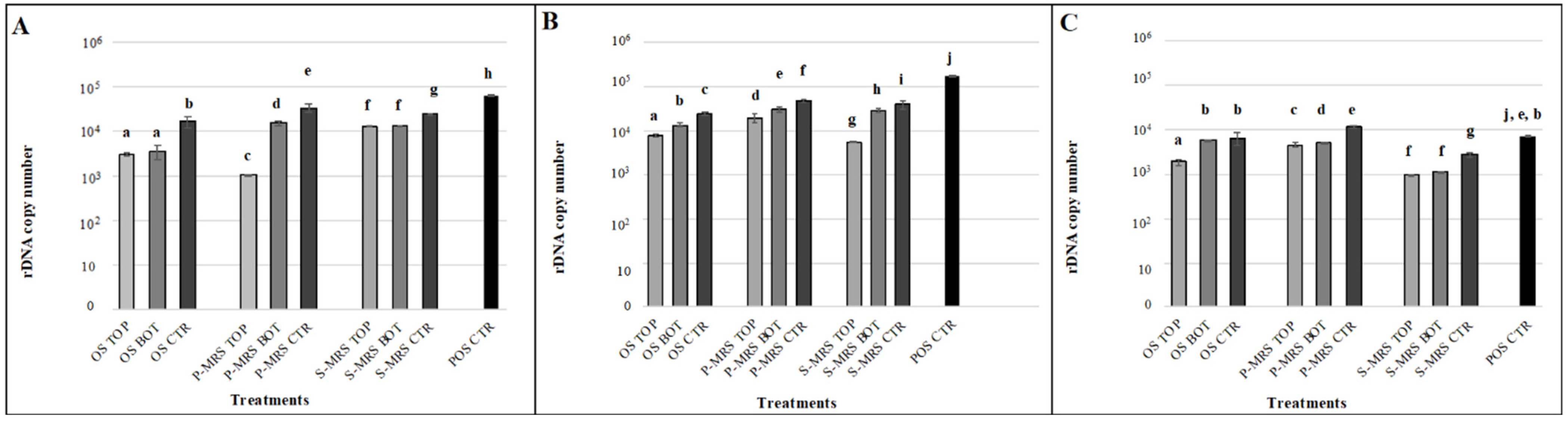
3.3. Detection of Nucleic Acids through Amplification Method
3.4. Organic Compounds Detection by Gas Chromatography Associated to Mass Spectrometry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grotzinger, J.P.; Sumner, D.Y.; Kah, L.C.; Stack, K.; Gupta, S.; Edgar, L.; Rubin, D.; Lewis, K.; Schieber, J.; Mangold, N.; et al. A Habitable Fluvio-Lacustrine Environment at Yellowknife Bay, Gale Crater, Mars. Science 2013, 343, 1242777. [Google Scholar] [CrossRef] [PubMed]
- Davila, A.F.; Duport, L.G.; Melchiorri, R.; Jaenchen, J.; Valea, S.; de Los Rios, A.; Wierzchos, J. Hygroscopic salts and the po-tential for life on Mars. Astrobiology 2010, 10, 617–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifford, S.M.; Lasue, J.; Heggy, E.; Boisson, J.; McGovern, P.; Max, M.D. Depth of the Martian cryosphere: Revised estimates and implications for the existence and detection of subpermafrost groundwater. J. Geophys. Res. Space Phys. 2010, 115, E07001. [Google Scholar] [CrossRef]
- Ehlmann, B.L.; Mustard, J.F.; Murchie, S.; Bibring, J.-P.; Meunier, A.; Fraeman, A.; Langevin, Y. Subsurface water and clay mineral formation during the early history of Mars. Nature 2011, 479, 53–60. [Google Scholar] [CrossRef]
- Thomas, R.J.; Hynek, B.M.; Osterloo, M.M.; Kierein-Young, K.S. Widespread exposure of Noachian phyllosilicates in the Margaritifer region of Mars: Implications for paleohydrology and astrobiological detection. J. Geophys. Res. Planets 2017, 122, 483–500. [Google Scholar] [CrossRef]
- Travis, B.J.; Rosenberg, N.D.; Cuzzi, J.N. On the role of widespread subsurface convection in bringing liquid water close to Mars’ surface. J. Geophys. Res. Space Phys. 2003, 108, 8040. [Google Scholar] [CrossRef]
- Dartnell, L.R.; Desorgher, L.; Ward, J.; Coates, A.J. Modelling the surface and subsurface Martian radiation environment: Implications for astrobiology. Geophys. Res. Lett. 2007, 34, L02207. [Google Scholar] [CrossRef] [Green Version]
- Dartnell, L.R.; Page, K.; Jorge-Villar, S.E.; Wright, G.; Munshi, T.; Scowen, I.J.; Edwards, H.G. Destruction of Raman biosig-natures by ionising radiation and the implications for life detection on Mars. Anal. Bioanal. Chem. 2012, 403, 131–144. [Google Scholar] [CrossRef]
- Hassler, D.M.; Zeitlin, C.; Wimmer-Schweingruber, R.F.; Ehresmann, B.; Rafkin, S.; Eigenbrode, J.L.; Brinza, D.E.; Weigle, G.; Böttcher, S.; Böhm, E.; et al. Mars’ Surface Radiation Environment Measured with the Mars Science Laboratory’s Curiosity Rover. Science 2013, 343, 1244797. [Google Scholar] [CrossRef] [Green Version]
- Vago, J.; Witasse, O.; Svedhem, H.; Baglioni, P.; Haldemann, A.; Gianfiglio, G.; Blancquaert, T.; McCoy, D.; De Groot, R. ESA ExoMars program: The next step in exploring Mars. Sol. Syst. Res. 2015, 49, 518–528. [Google Scholar] [CrossRef]
- Westall, F.; Foucher, F.; Bost, N.; Bertrand, M.; Loizeau, D.; Vago, J.L.; Kminek, G.; Gaboyer, F.; Campbell, K.; Bréhéret, J.-G.; et al. Biosignatures on Mars: What, Where, and How? Implications for the Search for Martian Life. Astrobiology 2015, 15, 998–1029. [Google Scholar] [CrossRef] [PubMed]
- Summons, R.E.; Amend, J.P.; Bish, D.L.; Buick, R.; Cody, G.D.; Marais, D.J.D.; Dromart, G.; Eigenbrode, J.L.; Knoll, A.H.; Sumner, D. Preservation of Martian Organic and Environmental Records: Final Report of the Mars Biosignature Working Group. Astrobiology 2011, 11, 157–181. [Google Scholar] [CrossRef]
- Vago, J.L.; Westall, F.; Teams, L.S.P.I.; Coates, A.; Jaumann, R.; Korablev, O.; Ciarletti, V.; Mitrofanov, I.; Josset, J.-L.; De Sanctis, M.C.; et al. Habitability on Early Mars and the Search for Biosignatures with the ExoMars Rover. Astrobiology 2017, 17, 471–510. [Google Scholar] [CrossRef]
- Aerts, J.W.; Röling, W.F.; Elsaesser, A.; Ehrenfreund, P. Biota and biomolecules in extreme environments on Earth: Implica-tions for life detection on Mars. Life 2014, 4, 535–565. [Google Scholar] [CrossRef] [PubMed]
- Eigenbrode, J.L.; Summons, R.E.; Steele, A.; Freissinet, C.; Millan, M.; Navarro-González, R.; Sutter, B.; McAdam, A.C.; Franz, H.B.; Glavin, D.P.; et al. Organic matter preserved in 3-billion-year-old mudstones at Gale crater, Mars. Science 2018, 360, 1096–1101. [Google Scholar] [CrossRef] [Green Version]
- Ellery, A.; Wynnwilliams, D.D. Why Raman Spectroscopy on Mars?—A Case of the Right Tool for the Right Job. Astrobiology 2003, 3, 565–579. [Google Scholar] [CrossRef]
- Wynn-Williams, D.; Edwards, H. Antarctic ecosystems as models for extraterrestrial surface habitats. Planet. Space Sci. 2000, 48, 1065–1075. [Google Scholar] [CrossRef]
- Onofri, S.; Selbmann, L.; Zucconi, L.; Pagano, S. Antarctic microfungi as models for exobiology. Planet. Space Sci. 2004, 52, 229–237. [Google Scholar] [CrossRef]
- Preston, L.J.; Barcenilla, R.; Dartnell, L.R.; Kucukkilic-Stephens, E.; Olsson-Francis, K. Infrared Spectroscopic Detection of Biosignatures at Lake Tírez, Spain: Implications for Mars. Astrobiology 2019, 20, 15–25. [Google Scholar] [CrossRef] [Green Version]
- de Vera, J.P.; Boettger, U.; de la Torre Noetzel, R.; Sánchez, F.J.; Grunow, D.; Schmitz, N.; Rettberg, P. Supporting Mars ex-ploration: BIOMEX in Low Earth Orbit and further astrobiological studies on the Moon using Raman and PanCam technology. Planet. Space Sci. 2012, 74, 103–110. [Google Scholar] [CrossRef]
- Onofri, S.; De La Torre, R.; De Vera, J.-P.; Ott, S.; Zucconi, L.; Selbmann, L.; Scalzi, G.; Venkateswaran, K.J.; Rabbow, E.; Iñigo, F.J.S.; et al. Survival of Rock-Colonizing Organisms After 1.5 Years in Outer Space. Astrobiology 2012, 12, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Onofri, S.; de Vera, J.P.; Zucconi, L.; Selbmann, L.; Scalzi, G.; Venkateswaran, K.J.; Horneck, G. Survival of Antarctic crypto-endolithic fungi in simulated Martian conditions on board the International Space Station. Astrobiology 2015, 15, 1052–1059. [Google Scholar] [CrossRef] [PubMed]
- Böttger, U.; de Vera, J.P.; Fritz, J.; Weber, I.; Hübers, H.W.; Schulze-Makuch, D. Optimizing the detection of carotene in cya-nobacteria in a martian regolith analogue with a Raman spectrometer for the ExoMars mission. Planet. Space Sci. 2012, 60, 356–362. [Google Scholar] [CrossRef]
- Rabbow, E.; Rettberg, P.; Barczyk, S.; Bohmeier, M.; Parpart, A.; Panitz, C.; Horneck, G.; Von Heise-Rotenburg, R.; Hoppenbrouwers, T.; Willnecker, R.; et al. EXPOSE-E: An ESA Astrobiology Mission 1.5 Years in Space. Astrobiology 2012, 12, 374–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbow, E.; Rettberg, P.; Barczyk, S.; Bohmeier, M.; Parpart, A.; Panitz, C.; Horneck, G.; Burfeindt, J.; Molter, F.; Jaramillo, E.; et al. The astrobiological mission EXPOSE-R on board of the International Space Station. Int. J. Astrobiol. 2014, 14, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Rosas, A.L.; Nosanchuk, J.D.; Feldmesser, M.; Cox, G.M.; McDade, H.C.; Casadevall, A. Synthesis of Polymerized Melanin by Cryptococcus neoformans in Infected Rodents. Infect. Immun. 2000, 68, 2845–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, N.M.; Ramasamy, S. Genetic validation and spectroscopic detailing of DHN-melanin extracted from an environmental fungus. Biochem. Biophys. Rep. 2017, 12, 98–107. [Google Scholar] [CrossRef]
- Baqué, M.; Hanke, F.; Böttger, U.; Leya, T.; Moeller, R.; De Vera, J.-P. Protection of cyanobacterial carotenoids’ Raman signatures by Martian mineral analogues after high-dose gamma irradiation. J. Raman Spectrosc. 2018, 49, 1617–1627. [Google Scholar] [CrossRef]
- Selbmann, L.; De Hoog, G.S.; Mazzaglia, A.; Friedmann, E.I.; Onofri, S. Fungi at the edge of life: Cryptoendolithic black fungi from Antarctic desert. Stud. Mycol. 2005, 51, 1–32. [Google Scholar]
- Cubeta, M.A.; Echandi, E.; Abernethy, T.; Vilgalys, R. Characterization of anastomosis groups of binucleate Rhizoctonia species using restriction analysis of an amplified ribosomal RNA gene. Phytopathology 1991, 81, 1395–1400. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Holland, P.M.; Chutjian, A.; Darrach, M.R.; Orient, O.J. Miniaturized GC/MS instrumentation for in situ measurements: Micro gas chromatography coupled with miniature quadrupole array and Paul ion trap mass spectrometers. Proc. SPIE 2003, 4878, 1–7. [Google Scholar] [CrossRef]
- Bell, A.A.; Wheeler, M.H. Biosynthesis and functions of fungal melanins. Annu. Rev. Phytopathol. 1986, 24, 411–451. [Google Scholar] [CrossRef]
- Cockell, C.S.; Knowland, J. Ultraviolet radiation screening compounds. Biol. Rev. 1999, 74, 311–345. [Google Scholar] [CrossRef]
- Pacelli, C.; Cassaro, A.; Baqué, M.; Selbmann, L.; Zucconi, L.; Maturilli, A.; Botta, L.; Saladino, R.; de Vera, J.-P.P.; Onofri, S.; et al. Fungal biomarkers are detectable in Martian rock-analogues after space exposure: Implications for the search of life on Mars. Int. J. Astrobiol. 2020, in press. [Google Scholar]
- Culka, A.; Jehlička, J.; Ascaso, C.; Artieda, O.; Casero, M.C.; Wierzchos, J. Raman microspectrometric study of pigments in melanized fungi from the hyperarid Atacama desert gypsum crust. J. Raman Spectrosc. 2017, 48, 1487–1493. [Google Scholar] [CrossRef]
- Samokhvalov, A.; Liu, Y.; Simon, J.D. Characterization of the Fe(III)-binding Site in Sepia Eumelanin by Resonance Raman Confocal Microspectroscopy. Photochem. Photobiol. 2007, 80, 84–88. [Google Scholar] [CrossRef]
- Galván, I.; Jorge, A.; Ito, K.; Tabuchi, K.; Solano, F.; Wakamatsu, K. Raman spectroscopy as a non-invasive technique for the quantification of melanins in feathers and hairs. Pigment. Cell Melanoma Res. 2013, 26, 917–923. [Google Scholar] [CrossRef]
- Thiel, C.S.; Ehrenfreund, P.; Foing, B.; Pletser, V.; Ullrich, O. PCR-based analysis of microbial communities during the Eu-roGeoMars campaign at Mars Desert Research Station, Utah. Int. J. Astrobiol. 2012, 10, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Goesmann, F.; Brinckerhoff, W.B.; Raulin, F.; Goetz, W.; Danell, R.; Getty, S.A.; Siljeström, S.; Mißbach, H.; Steininger, H.; Arevalo, R.D.; et al. The Mars Organic Molecule Analyzer (MOMA) Instrument: Characterization of Organic Material in Martian Sediments. Astrobiology 2017, 17, 655–685. [Google Scholar] [CrossRef]
- Mahaffy, P.R.; Webster, C.R.; Cabane, M.; Conrad, P.G.; Coll, P.; Atreya, S.K.; Arvey, R.; Barciniak, M.; Benna, M.; Bleacher, L.; et al. The Sample Analysis at Mars Investigation and Instrument Suite. Space Sci. Rev. 2012, 170, 401–478. [Google Scholar] [CrossRef]
- Melosh, H.J. Exchange of Meteorites (and Life?) Between Stellar Systems. Astrobiology 2003, 3, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Claudia, P.; Alessia, C.; Siong, L.M.; Lorenzo, A.; Ralf, M.; Akira, F.; Silvano, O. Insights into the Survival Capabilities of Cryomyces antarcticus Hydrated Colonies after Exposure to Fe Particle Radiation. J. Fungi 2021, 7, 495. [Google Scholar]
- Aureli, L.; Pacelli, C.; Cassaro, A.; Fujimori, A.; Moeller, R.; Onofri, S. Iron Ion Particle Radiation Resistance of Dried Colonies of Cryomyces antarcticus Embedded in Martian Regolith Analogues. Life 2020, 10, 306. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2011, 93, 931–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhartia, R.; Hug, W.F.; Reid, R.D.; Beegle, L.W. Explosives detection and analysis by fusing deep ultraviolet native fluorescence and resonance Raman spectroscopy. In Laser-Based Optical Detection of Explosives; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Samokhvalov, A.; Garguilo, J.; Yang, W.-C.; Edwards, G.S.; Nemanich, R.J.; Simon, J.D. Photoionization Threshold of Eumelanosomes Determined Using UV Free Electron Laser−Photoelectron Emission Microscopy. J. Phys. Chem. B 2004, 108, 16334–16338. [Google Scholar] [CrossRef]
- Pacelli, C.; Bryan, R.A.; Onofri, S.; Selbmann, L.; Zucconi, L.; Shuryak, I.; Dadachova, E. The effect of protracted X-ray exposure on cell survival and metabolic activity of fast and slow growing fungi capable of melanogenesis. Environ. Microbiol. Rep. 2018, 10, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Van de Meent, D.; Brown, S.C.; Philp, R.; Simoneit, B.R. Pyrolysis-high resolution gas chromatography and pyrolysis gas chromatography-mass spectrometry of kerogens and kerogen precursors. Geochim. Cosmochim. Acta 1980, 44, 999–1013. [Google Scholar] [CrossRef]
- Horsfield, B. Practical criteria for classifying kerogens: Some observations from pyrolysis-gas chromatography. Geochim. et Cosmochim. Acta 1989, 53, 891–901. [Google Scholar] [CrossRef]
- Eglinton, T.; Damste, J.S.; Pool, W.; de Leeuw, J.W.; Eijkel, G.; Boon, J.J. Organic sulphur in macromolecular sedimentary organic matter. II. Analysis of distributions of sulphur-containing pyrolysis products using multivariate techniques. Geochim. et Cosmochim. Acta 1992, 56, 1545–1560. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, M.; Goetz, W.; Thiel, V. Testing Flight-like Pyrolysis Gas Chromatography–Mass Spectrometry as Performed by the Mars Organic Molecule Analyzer Onboard the ExoMars 2020 Rover on Oxia Planum Analog Samples. Astrobiology 2020, 20, 415–428. [Google Scholar] [CrossRef]
- Faure, P.; Jeanneau, L.; Lannuzel, F. Analysis of organic matter by flash pyrolysis-gas chromatography–mass spectrometry in the presence of Na-smectite: When clay minerals lead to identical molecular signature. Org. Geochem. 2006, 37, 1900–1912. [Google Scholar] [CrossRef]
- Moldoveanu, S. Chapter 17 Pyrolysis of Carboxylic Acids. Tech. Instrum. Anal. Chem. 2010, 28, 471–526. [Google Scholar] [CrossRef]
- Dorn, E.D.; Nealson, K.H.; Adami, C. Monomer Abundance Distribution Patterns as a Universal Biosignature: Examples from Terrestrial and Digital Life. J. Mol. Evol. 2011, 72, 283–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKay, C.P. What is life—And how do we search for it in other worlds? PLoS Biol. 2004, 2, e302. [Google Scholar] [CrossRef] [PubMed]
- Mustard, J.F.; Adler, M.; Allwood, A.; Bass, D.S.; Beaty, D.W.; Bell, J.F.; Edgett, K.S. Report of the Mars 2020 science definition team. Mars Explor. Progr. Anal. Gr. 2013, 150, 155–205. [Google Scholar]
- Neveu, M.; Hays, L.E.; Voytek, M.A.; New, M.H.; Schulte, M.D. The Ladder of Life Detection. Astrobiology 2018, 18, 1375–1402. [Google Scholar] [CrossRef] [Green Version]
- Kminek, G.; Bada, J. The effect of ionizing radiation on the preservation of amino acids on Mars. Earth Planet. Sci. Lett. 2006, 245, 1–5. [Google Scholar] [CrossRef]
- Dabney, J.; Meyer, M.; Pääbo, S. Ancient DNA damage. Csh. Perspect. Biol. 2013, 5, a012567. [Google Scholar] [CrossRef]
- Millar, C.D.; Lambert, D.M. Ancient DNA: Towards a million-year-old genome. Nature 2013, 499, 34–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, M.; Zeelen, J.P.; Mainfroid, V.; Rentier-Delrue, F.; Martial, J.A.; Wyns, L.; Maes, D. Triose-phosphate Isomerase (TIM) of the Psychrophilic Bacterium Vibrio marinus Kinetic and Structural Properties. J. Biol. Chem. 1998, 273, 2199–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isenbarger, T.A.; Carr, C.E.; Johnson, S.S.; Finney, M.; Church, G.M.; Gilbert, W.; Zuber, M.T.; Ruvkun, G. The Most Conserved Genome Segments for Life Detection on Earth and Other Planets. Orig. Life Evol. Biosphere 2008, 38, 517–533. [Google Scholar] [CrossRef] [PubMed]
- Fajardo-Cavazos, P.; Schuerger, A.C.; Nicholson, W.L. Exposure of DNA and Bacillus subtilis Spores to Simulated Martian Environments: Use of Quantitative PCR (qPCR) to Measure Inactivation Rates of DNA to Function as a Template Molecule. Astrobiology 2010, 10, 403–411. [Google Scholar] [CrossRef]
- Haberle, R.M.; Juarez, M.D.L.T.; Kahre, M.A.; Kass, D.M.; Barnes, J.R.; Hollingsworth, J.L.; Harri, A.-M.; Kahanpää, H. Detection of Northern Hemisphere transient eddies at Gale Crater Mars. Icarus 2018, 307, 150–160. [Google Scholar] [CrossRef]
- John, K.K.; Botkin, D.S.; Burton, A.S.; Castro-Wallace, S.L.; Chaput, J.D.; Dworkin, J.P.; Stahl, S. The Biomolecule Sequencer Project: Nanopore Sequencing as a Dual-Use Tool for Crew Health and Astrobiology Investigations. 2016. Available online: https://pdxscholar.library.pdx.edu/chem_fac/123/ (accessed on 8 October 2021).
- Castro-Wallace, S.L.; Chiu, C.Y.; John, K.K.; Stahl, S.E.; Rubins, K.H.; McIntyre, A.B.; Stephenson, T.A. Nanopore DNA se-quencing and genome assembly on the International Space Station. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Maggiori, C.; Stromberg, J.; Blanco, Y.; Goordial, J.; Cloutis, E.; García-Villadangos, M.; Parro, V.; Whyte, L. The Limits, Capabilities, and Potential for Life Detection with MinION Sequencing in a Paleochannel Mars Analog. Astrobiology 2020, 20, 375–393. [Google Scholar] [CrossRef]
- Javaux, E.J. Extreme life on Earth—past, present and possibly beyond. Res. Microbiol. 2006, 157, 37–48. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Test Parameters | Duration |
|---|---|
| Vacuum (2 × 10−4) + polychromatic UV irradiation (200–400 nm), with SOL2000 at 1271.2 W/m2, attenuated with 0.1% neutral density filter. | 28 days |
| SOL2000 | |
| 125 h | |
| Simulated CO2 Mars atmosphere 103 Pa+ polychromatic UV irradiation (200–400 nm), with SOL2000 at 1271.2 W/m2, attenuated with 0.1% neutral density filter. | 28 days |
| SOL2000 | |
| 125 h | |
| Control experiment, 1 atm air, dark, room temperature | 28 days |
| Compounds [a] | OS Top | OS Control | P-MRS Top | P-MRS Control | S-MRS Top | S-MRS Control |
|---|---|---|---|---|---|---|
| Azelaic acid | 0.99 | 0.13 | 0.69 | 0.45 | 0.30 | 0.02 |
| Myristic acid | nd | nd | nd | nd | 0.48 | 0.42 |
| Pyruvic acid | nd | nd | nd | nd | nd | 0.17 |
| Glucose | 3.14 | 2.78 | nd | nd | 0.68 | 13.87 |
| Fructose | 0.64 | 7.53 | nd | nd | nd | 4.50 |
| Glucitol | nd | 3.58 | nd | nd | 0.88 | 2.50 |
| Glycerol | nd | nd | nd | nd | nd | nd |
| Ethylene glycol | nd | nd | 1.73 | nd | 0.41 | nd |
| Products [a] | m/z (%) |
|---|---|
| Azelaic Acid [c] | 317 (25) [M-CH3], 302 (3) [M-2xCH3], 243 (2) [M-OSi(CH3)3], 201 (15) [M-Si(CH3)3-CO2-CH3], 186 (3) [M-2xSi(CH3)3], 170 (4) [M-OSi(CH3)3- Si(CH3)3], 73 (100). |
| Myristic Acid [b] | 300 (10) [M], 285 (95) [M-CH3], 257 (3) [M-2xCH3], 73 (100). |
| Palmitic Acid [b] | 328 (20) [M], 313 (100) [M-CH3], 73 (100). |
| Stearic Acid [b] | 356 (20) [M], 341 (90) [M-CH3], 327 (2) [M-CH3- CH2], 313 (50) [M-CH3-2xCH2]. |
| Lactic Acid [b] | 219 (6) [M-CH3], 190 (14) [M-CO2], 147 (71) [M-Si(CH3)3-CH3], 133 (7), 117 (76) [M-Si(CH3)3-(CH3)3]. |
| Pyruvic acid [b] | 160 (10) [M], 145 (7) [M-CH3], 88 (14) [M-Si(CH3)3], 71 (12) [M-Si(CH3)3-OH], 43 (100) [M-HSi(CH3)3- CO2]. |
| Glucose [e] | 437 (5) [M-Si(CH3)3-2xCH3], 394 (4) [M-2xSi(CH3)3], 305 (5) [M-OSi(CH3)3-2xSi(CH3)3], 217 [g] (30), 204 [g] (100), 191 [g] (75). |
| Fructose [e] | 437 (5) [M-Si(CH3)3-2xCH3], 217 [g] (30), 204 [g] (100), 146 (75). |
| Glucitol [f] | 319 (60) [M], 297 (94) [M-CH3], 282 (30) [M-2xCH3], 267 (40) [M-3xCH3]; 217 [g] (90), 204 [g] (80); 147 (40). |
| Glycerol [d] | 293 (3) [M-CH3], 263 (2) [M-3xCH3], 218 (20) [M-OSi(CH3)3], 205 (60) [M-OSi(CH3)3-CH3], 191 (3) [M-OSi(CH3)3-2xCH3], 171 (4) [M-OSi(CH3)3-3xCH3]. |
| Ethylene Glycol [c] | 191 (25) [M-CH3], 147 (100) [M-4xCH3], 133 (5) [M-Si(CH3)3], 103 (20) [M-Si(CH3)3-2xCH3]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cassaro, A.; Pacelli, C.; Baqué, M.; de Vera, J.-P.P.; Böttger, U.; Botta, L.; Saladino, R.; Rabbow, E.; Onofri, S. Fungal Biomarkers Stability in Mars Regolith Analogues after Simulated Space and Mars-like Conditions. J. Fungi 2021, 7, 859. https://doi.org/10.3390/jof7100859
Cassaro A, Pacelli C, Baqué M, de Vera J-PP, Böttger U, Botta L, Saladino R, Rabbow E, Onofri S. Fungal Biomarkers Stability in Mars Regolith Analogues after Simulated Space and Mars-like Conditions. Journal of Fungi. 2021; 7(10):859. https://doi.org/10.3390/jof7100859
Chicago/Turabian StyleCassaro, Alessia, Claudia Pacelli, Mickael Baqué, Jean-Pierre Paul de Vera, Ute Böttger, Lorenzo Botta, Raffaele Saladino, Elke Rabbow, and Silvano Onofri. 2021. "Fungal Biomarkers Stability in Mars Regolith Analogues after Simulated Space and Mars-like Conditions" Journal of Fungi 7, no. 10: 859. https://doi.org/10.3390/jof7100859