
Species Diversity and Distribution Characteristics of Calonectria in Five Soil Layers in a Eucalyptus Plantation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site, Soil Sampling, and Calonectria Isolation
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Multi-Gene Phylogenetic Analyses, Morphology, and Species Identification
2.4. Calonectria Species Diversity in Different Soil Layers
2.5. Genotyping of Isolates within Each Calonectria Species
2.6. Genotype Diversity of Calonectria Species in Different Soil Layers
3. Results
3.1. Soil Sampling and Calonectria Isolation
3.2. Sequencing
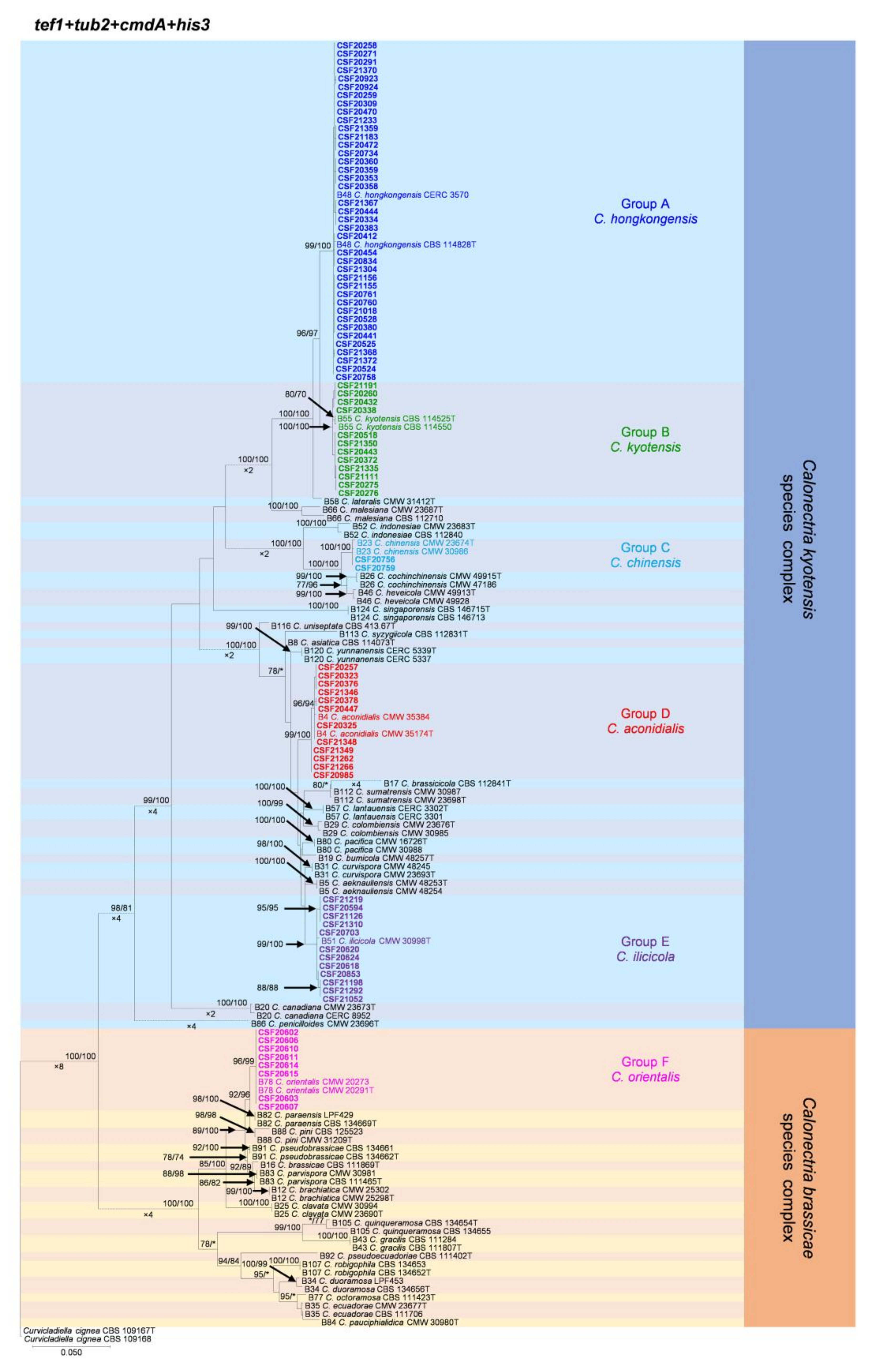
3.3. Multi-Gene Phylogenetic Analyses, Morphology, and Species Identification
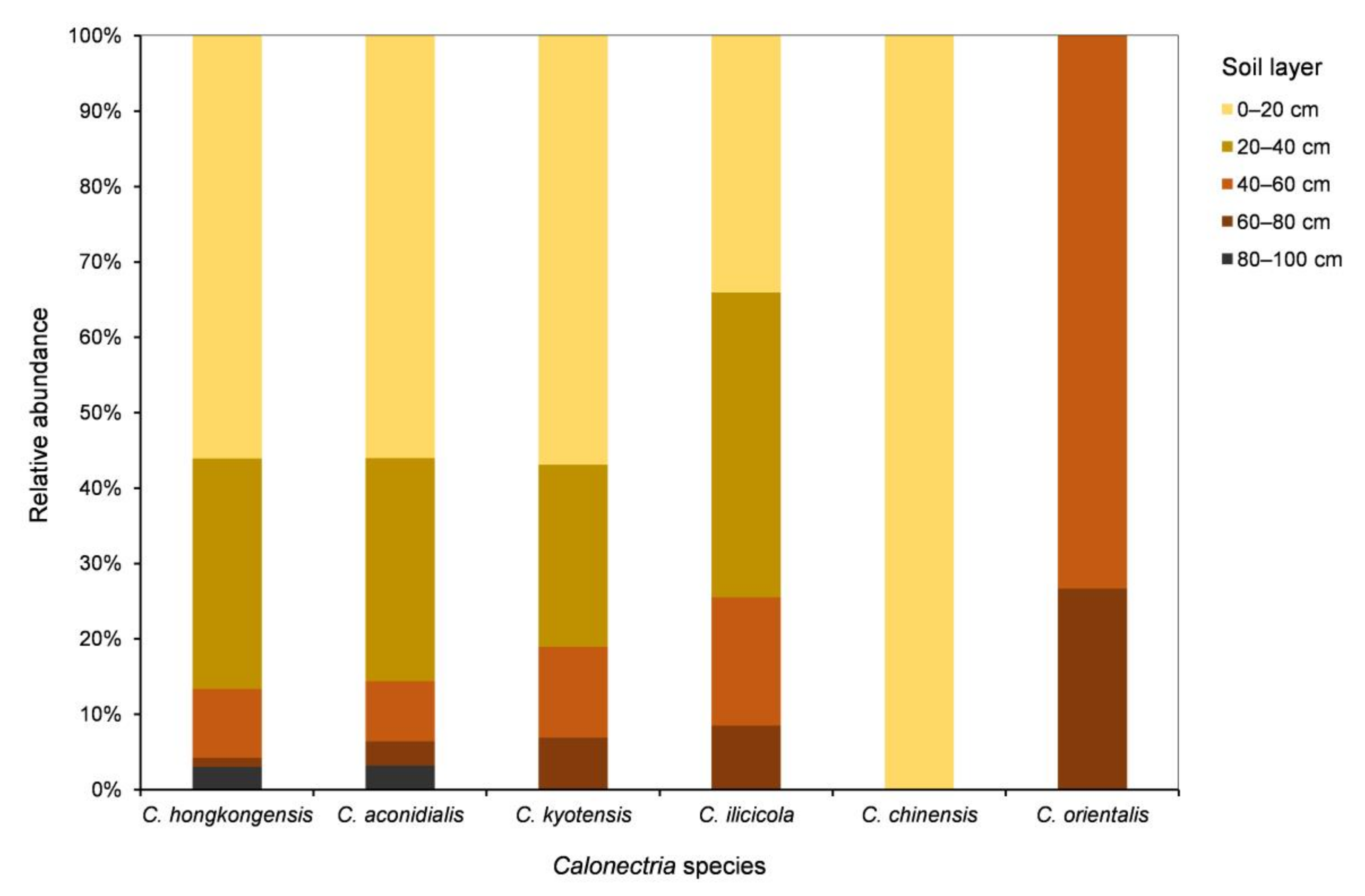
3.4. Calonectria Species Diversity in Different Soil Layers
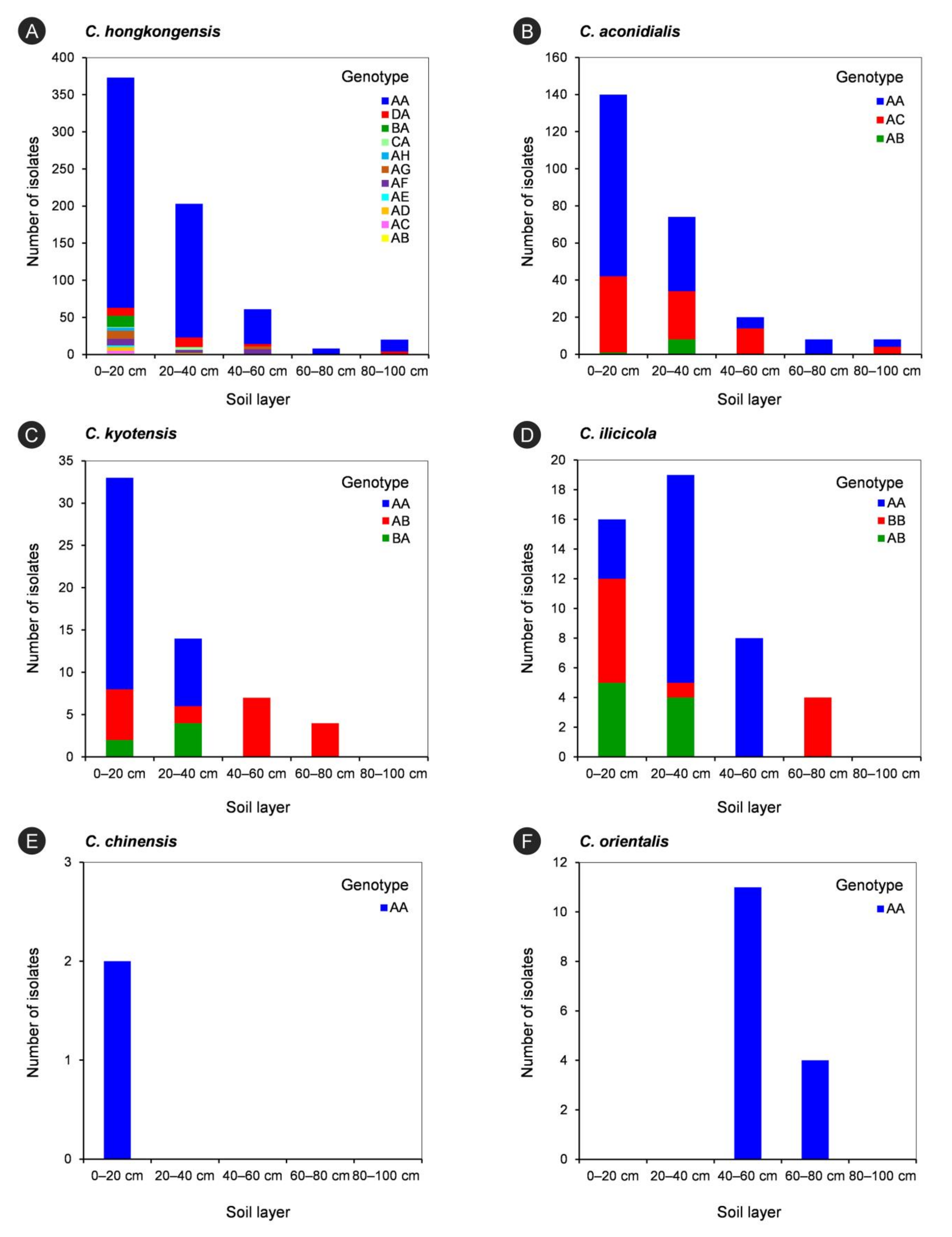
3.5. Genotyping of Isolates within Each Calonectria Species
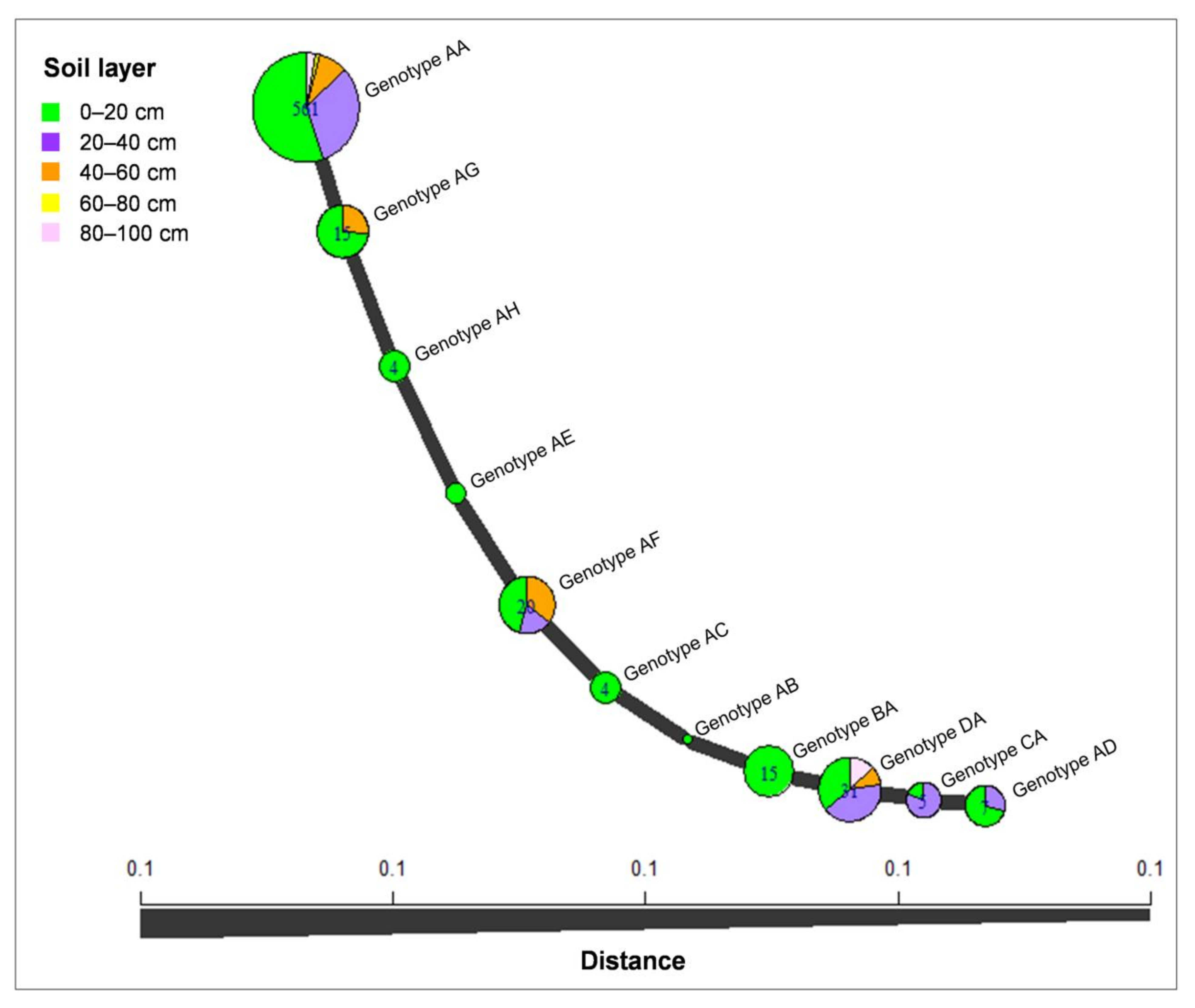
3.6. Genotype Diversity of Calonectria Species in Different Soil Layers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crous, P.W. Taxonomy and pathology of Cylindrocladium (Calonectria) and Allied Genera; APS Press: St. Paul, MN, USA, 2002. [Google Scholar]
- Lombard, L.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Species concepts in Calonectria (Cylindrocladium). Stud. Mycol. 2010, 66, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Vitale, A.; Crous, P.W.; Lombard, L.; Polizzi, G. Calonectria diseases on ornamental plants in Europe and the Mediterranean basin: An overview. J. Plant Pathol. 2013, 95, 463–476. [Google Scholar]
- Daughtrey, M.L. Boxwood blight: Threat to ornamentals. Annu. Rev. Phytopathol. 2019, 57, 189–209. [Google Scholar] [CrossRef]
- Liu, Q.L.; Li, J.Q.; Wingfield, M.J.; Duong, T.A.; Wingfield, B.D.; Crous, P.W.; Chen, S.F. Reconsideration of species boundaries and proposed DNA barcodes for Calonectria. Stud. Mycol. 2020, 97, 100106. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Q.; Barnes, I.; Liu, F.F.; Wingfield, M.J.; Chen, S.F. Global genetic diversity and mating type distribution of Calonectria pauciramosa: An important wide host-range plant pathogen. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Lombard, L.; Rodas, C.A.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Calonectria (Cylindrocladium) species associated with dying Pinus cuttings. Persoonia 2009, 23, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombard, L.; Zhou, X.D.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Calonectria species associated with cutting rot of Eucalyptus. Persoonia 2010, 24, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombard, L.; Chen, S.F.; Mou, X.; Zhou, X.D.; Crous, P.W.; Wingfield, M.J. New species, hyper-diversity and potential importance of Calonectria spp. from Eucalyptus in South China. Stud. Mycol. 2015, 80, 151–188. [Google Scholar] [CrossRef] [Green Version]
- Alfenas, R.F.; Lombard, L.; Pereira, O.L.; Alfenas, A.C.; Crous, P.W. Diversity and potential impact of Calonectria species in Eucalyptus plantations in Brazil. Stud. Mycol. 2015, 80, 89–130. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Q.; Wingfield, M.J.; Liu, Q.L.; Barnes, I.; Roux, J.; Lombard, L.; Crous, P.W.; Chen, S.F. Calonectria species isolated from Eucalyptus plantations and nurseries in South China. IMA Fungus 2017, 8, 259–286. [Google Scholar] [CrossRef] [Green Version]
- Pham, N.; Barnes, I.; Chen, S.F.; Liu, F.F.; Dang, Q.; Pham, T.; Lombard, L.; Crous, P.W.; Wingfield, M.J. Ten new species of Calonectria from Indonesia and Vietnam. Mycologia 2019, 111, 78–102. [Google Scholar] [CrossRef]
- Wang, Q.C.; Liu, Q.L.; Chen, S.F. Novel species of Calonectria isolated from soil near Eucalyptus plantations in southern China. Mycologia 2019, 111, 1028–1040. [Google Scholar] [CrossRef]
- Wu, W.X.; Chen, S.F. Species diversity, mating strategy, and pathogenicity of Calonectria species from diseased leaves and soils in the Eucalyptus plantation in southern China. J. Fungi 2021, 7, 73. [Google Scholar] [CrossRef]
- Old, K.M.; Wingfield, M.J.; Yuan, Z.Q. A Manual of Diseases of Eucalypts in South-East Asia; Center for International Forestry Research: Jakarta, Indonesia, 2003. [Google Scholar]
- Rodas, C.A.; Lombard, L.; Gryzenhout, M.; Slippers, B.; Wingfield, M.J. Cylindrocladium blight of Eucalyptus grandis in Colombia. Australas. Plant Pathol. 2005, 34, 143–149. [Google Scholar] [CrossRef]
- Lechat, C.; Crous, P.W.; Groenewald, J.Z. The enigma of Calonectria species occurring on leaves of Ilex aquifolium in Europe. IMA Fungus 2010, 1, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.F.; Lombard, L.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Novel species of Calonectria associated with Eucalyptus leaf blight in Southeast China. Persoonia 2011, 26, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gehesquière, B.; Crouch, J.A.; Marra, R.E.; van Poucke, K.; Rys, F.; Maes, M.; Gobin, B.; Höfte, M.; Heungens, K. Characterization and taxonomic reassessment of the box blight pathogen Calonectria pseudonaviculata, introducing Calonectria henricotiae sp. nov. Plant Pathol. 2016, 65, 37–52. [Google Scholar] [CrossRef]
- Lopes, U.P.; Alfenas, R.F.; Zambolim, L.; Crous, P.W.; Costa, H.; Pereira, O.L. A new species of Calonectria causing rot on ripe strawberry fruit in Brazil. Australas. Plant Pathol. 2017, 47, 1–11. [Google Scholar] [CrossRef]
- Gai, Y.; Deng, Q.; Chen, X.; Guan, M.; Xiao, X.; Xu, D.; Deng, M.; Pan, R. Phylogenetic diversity of Calonectria ilicicola causing Cylindrocladium black rot of peanut and red crown rot of soybean in southern China. J. Gen. Plant Pathol. 2017, 83, 273–282. [Google Scholar] [CrossRef]
- Freitas, R.G.; Alfenas, R.F.; Guimarães, L.M.S.; Badel, J.L.; Alfenas, A.C. Genetic diversity and aggressiveness of Calonectria pteridis in Eucalyptus spp. Plant Pathol. 2019, 68, 869–877. [Google Scholar] [CrossRef]
- Wang, Q.C.; Chen, S.F. Calonectria pentaseptata causes severe leaf disease on cultivated Eucalyptus in Leizhou Peninsula of southern China. Plant Dis. 2020, 104, 493–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, D.K.; Sobers, E.K. A peg, pod, and root necrosis of peanuts caused by a species of Calonectria. Phytopathology 1966, 56, 1361–1364. [Google Scholar]
- Johnston, S.A.; Beute, M.K. Histopathology of Cylindrocladium black rot of peanut. Phytopathology 1975, 64, 649–653. [Google Scholar] [CrossRef]
- Pataky, J.K.; Beute, M.K.; Wynne, J.C.; Carlson, G.A. Peanut yield, market quality and value reductions due to Cylindrocladium black rot. Peanut Sci. 1983, 10, 62–66. [Google Scholar] [CrossRef]
- Berner, D.K.; Berggren, G.T.; Snow, J.P.; White, E.P. Distribution and management of red crown rot of soybean in Louisiana. Appl. Agric. Res. 1988, 3, 160–166. [Google Scholar]
- Yamamoto, R.; Nakagawa, A.; Shimada, S.; Komatsu, S.; Kanematsu, S. Histopathology of red crown rot of soybean. J. Gen. Plant Pathol. 2016, 83, 23–32. [Google Scholar] [CrossRef]
- Lombard, L.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Phylogeny and systematics of the genus Calonectria. Stud. Mycol. 2010, 66, 31–69. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; Wingfield, M.J.; Alfenas, A.C.; Crous, P.W. The forgotten Calonectria collection: Pouring old wine into new bags. Stud. Mycol. 2016, 85, 159–198. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef]
- Crous, P.W.; Hernández-Restrepo, M.; Schumacher, R.K.; Cowan, D.A.; Maggs-Kölling, G.; Marais, E.; Wingfield, M.J.; Yilmaz, N.; Adan, O.C.G.; Akulov, A.; et al. New and Interesting Fungi. 4. Fungal Syst. Evol. 2021, 7, 255–343. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.J.; Noordeloos, M.E.; Dima, B.; Brandrud, T.E.; Jansen, G.M.; et al. Fungal Planet description sheets: 1182–1283. Persoonia 2021, 46, 313–528. [Google Scholar]
- Liu, Q.L.; Chen, S.F. Two novel species of Calonectria isolated from soil in a natural forest in China. MycoKeys 2017, 26, 25–60. [Google Scholar] [CrossRef] [Green Version]
- Phipps, P.M.; Beute, M.K.; Barker, K.R. An elutriation method for quantitative isolation of Cylindrocladium crotalariae microsclerotia from peanut field soil. Phytopathology 1976, 66, 1255–1259. [Google Scholar] [CrossRef]
- Thies, W.F.; Patton, R.F. The biology of Cylindrocladium scoparium in Wisconsin forest tree nurseries. Phytopathology 1970, 60, 1662–1668. [Google Scholar] [CrossRef]
- Sobers, E.K.; Littrell, R.H. Pathogenicity of three species of Cylindrocladium to select hosts. Plant Dis. Report. 1974, 58, 1017–1019. [Google Scholar]
- Anderson, P.J. Rose Canker and Its Control; Massachusetts Agricultural Experiment Station Bulletin: Boston, MA, USA, 1918; Volume 183, pp. 11–46. [Google Scholar]
- Dumas, M.T.; Greifenhagen, S.; Halicki-Hayden, G.; Meyer, T.R. Effect of seedbed steaming on Cylindrocladium floridanum, soil microbes and the development of white pine seedlings. Phytoprotection 1998, 79, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Du, A.; Wang, Z.; Zhu, W.; Li, C.; Wu, L. Effects of different rotation periods of Eucalyptus plantations on soil physiochemical properties, enzyme activities, microbial biomass and microbial community structure and diversity. For. Ecol. Manag. 2020, 456, 117683. [Google Scholar] [CrossRef]
- Cooperative Research Group on Chinese Soil Taxonomy. Chinese Soil Taxonomy; Science Press: Beijing, China, 2001. [Google Scholar]
- van Burik, J.A.; Schreckhise, R.W.; White, T.C.; Bowden, R.A.; Myerson, D. Comparison of six extraction techniques for isolation of DNA from filamentous fungi. Med. Mycol. 1998, 36, 299–303. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Graça, R.N.; Alfenas, A.C.; Maffia, L.A.; Titon, M.; Alfenas, R.F.; Lau, D.; Rocabado, J.M.A. Factors influencing infection of eucalypts by Cylindrocladium pteridis. Plant Pathol. 2009, 58, 971–981. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, P.W.; Groenewald, J.Z.; Risede, J.M.; Simoneau, P.; Hywel-Jones, N.L. Calonectria species and their Cylindrocladium anamorphs: Species with sphaeropedunculate vesicles. Stud. Mycol. 2004, 50, 415–430. [Google Scholar]
- Kang, J.C.; Crous, P.W.; Schoch, C.L. Species concepts in the Cylindrocladium floridanum and Cy. spathiphylli complexes (Hypocreaceae) based on multi-allelic sequence data, sexual compatibility and morphology. Syst. Appl. Microbiol. 2001, 24, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Victor, D.; Crous, P.W.; Janse, B.J.H.; Wingfield, M.J. Genetic variation in Cylindrocladium floridanum and other morphologically similar Cylindrocladium species. Syst. Appl. Microbiol. 1997, 20, 268–285. [Google Scholar] [CrossRef] [Green Version]
- Boedijn, K.B.; Reitsma, J. Notes on the genus Cylindrocladium (Fungi: Mucedineae). Reinwardtia 1950, 1, 51–60. [Google Scholar]
- Terashita, T. A new species of Calonectria and its conidial state. Trans. Mycol. Soc. Japan 1968, 8, 124–129. [Google Scholar]
- Tubaki, K. Studies on the Japanese Hyphomycetes. V. Leaf & stem group with a discussion of the classification of Hyphomycetes and their perfect stages. J. Hattori Bot. Lab. 1958, 20, 142–244. [Google Scholar]
- El-Gholl, N.E.; Alfieri, S.A., Jr.; Barnard, E.L. Description and pathogenicity of Calonectria clavata sp. nov. Mycotaxon 1993, 48, 201–216. [Google Scholar]
- Crous, P.W.; Groenewald, J.Z.; Risède, J.M.; Simoneau, P.; Hyde, K.D. Calonectria species and their Cylindrocladium anamorphs: Species with clavate vesicles. Stud. Mycol. 2006, 55, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Alfenas, A.C. Additions to Calonectria. Mycotaxon 1993, 46, 217–234. [Google Scholar]
- Decock, C.; Crous, P.W. Curvicladium gen. nov., a new hyphomycete genus from French Guiana. Mycologia 1998, 90, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, C.W. Can three incongruence tests predict when data should be combined? Mol. Biol. Evol. 1997, 14, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Pei, W.H.; Cao, J.F.; Yang, M.Y.; Zhao, Z.J.; Xue, S.M. First report of black rot of medicago sativa caused by Cylindrocladium parasiticum (teleomorph Calonectria ilicicola) in Yunnan province, China. Plant Dis. 2015, 99, 890. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Jumpponen, A.; Jones, K.L.; Blair, J. Vertical distribution of fungal communities in tallgrass prairie soil. Mycologia 2010, 102, 1027–1041. [Google Scholar] [CrossRef] [PubMed]
- de Araujo Pereira, A.P.; Santana, M.C.; Bonfim, J.A.; de Lourdes Mescolotti, D.; Cardoso, E.J.B.N. Digging deeper to study the distribution of mycorrhizal arbuscular fungi along the soil profile in pure and mixed Eucalyptus grandis and Acacia mangium plantations. Appl. Soil Ecol. 2018, 128, 1–11. [Google Scholar] [CrossRef]
- Grishkan, I.; Lázaro, R.; Kidron, G.J. Vertical divergence of cultivable microfungal communities through biocrusted and bare soil profiles at the Tabernas Desert, Spain. Geomicrobiol. J. 2020, 37, 534–549. [Google Scholar] [CrossRef]
- Upton, R.N.; Sielaff, A.C.; Hofmockel, K.S.; Xu, X.; Polley, H.W.; Wilsey, B.J. Soil depth and grassland origin cooperatively shape microbial community co-occurrence and function. Ecosphere 2020, 11, e02973. [Google Scholar] [CrossRef] [Green Version]
- Frey, B.; Walthert, L.; Perez-Mon, C.; Stierli, B.; Köchli, R.; Dharmarajah, A.; Brunner, I. Deep soil layers of drought-exposed forests harbor poorly known bacterial and fungal communities. Front. Microbiol. 2021, 12, 674160. [Google Scholar] [CrossRef]
- Na, X.; Ma, S.; Ma, C.; Liu, Z.; Xu, P.; Zhu, H.; Liang, W.; Kardol, P. Lycium barbarum L. (goji berry) monocropping causes microbial diversity loss and induces Fusarium spp. enrichment at distinct soil layers. Appl. Soil Ecol. 2021, 168, 104107. [Google Scholar] [CrossRef]
- Lennon, J.T. Microbial life deep underfoot. mBio 2020, 11, e03201-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Layer | C. hongkongensis | C. aconidialis | C. kyotensis | C. ilicicola | C. chinensis | C. orientalis | All six Calonectria species | Percentage |
|---|---|---|---|---|---|---|---|---|
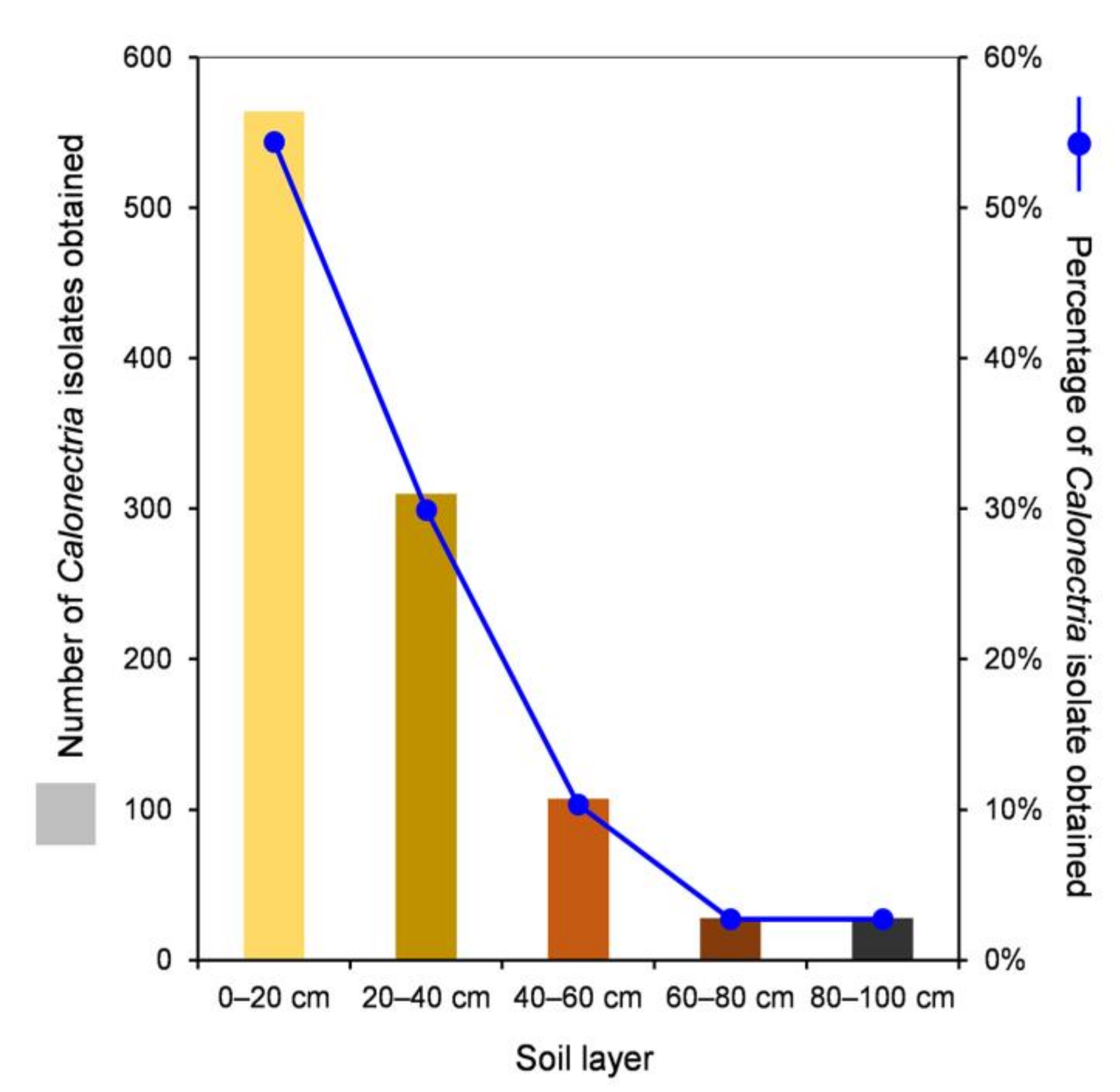
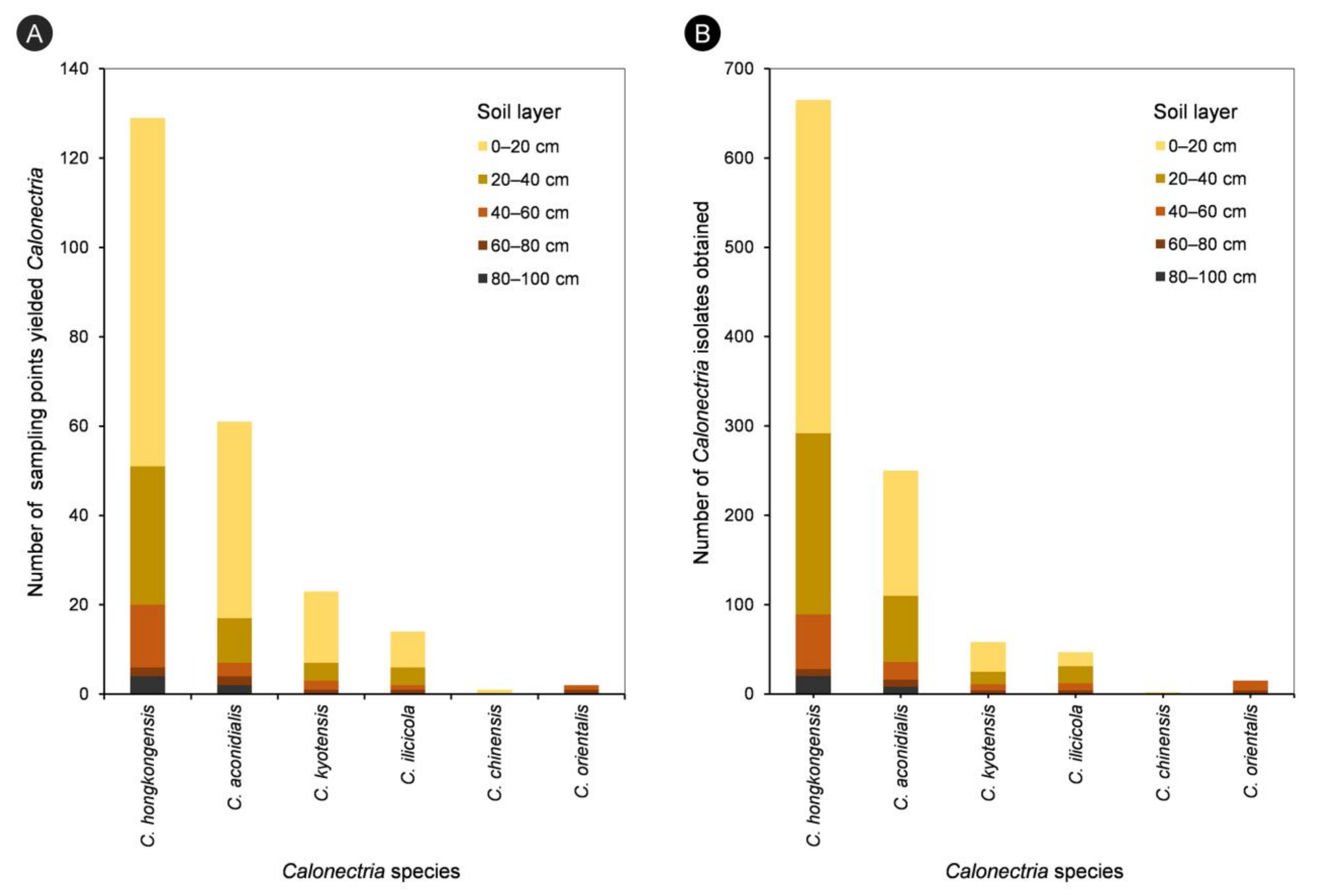
| 0–20 cm | 373 | 140 | 33 | 16 | 2 | 0 | 564 | 54.4% |
| 20–40 cm | 203 | 74 | 14 | 19 | 0 | 0 | 310 | 29.9% |
| 40–60 cm | 61 | 20 | 7 | 8 | 0 | 11 | 107 | 10.3% |
| 60–80 cm | 8 | 8 | 4 | 4 | 0 | 4 | 28 | 2.7% |
| 80–100 cm | 20 | 8 | 0 | 0 | 0 | 0 | 28 | 2.7% |
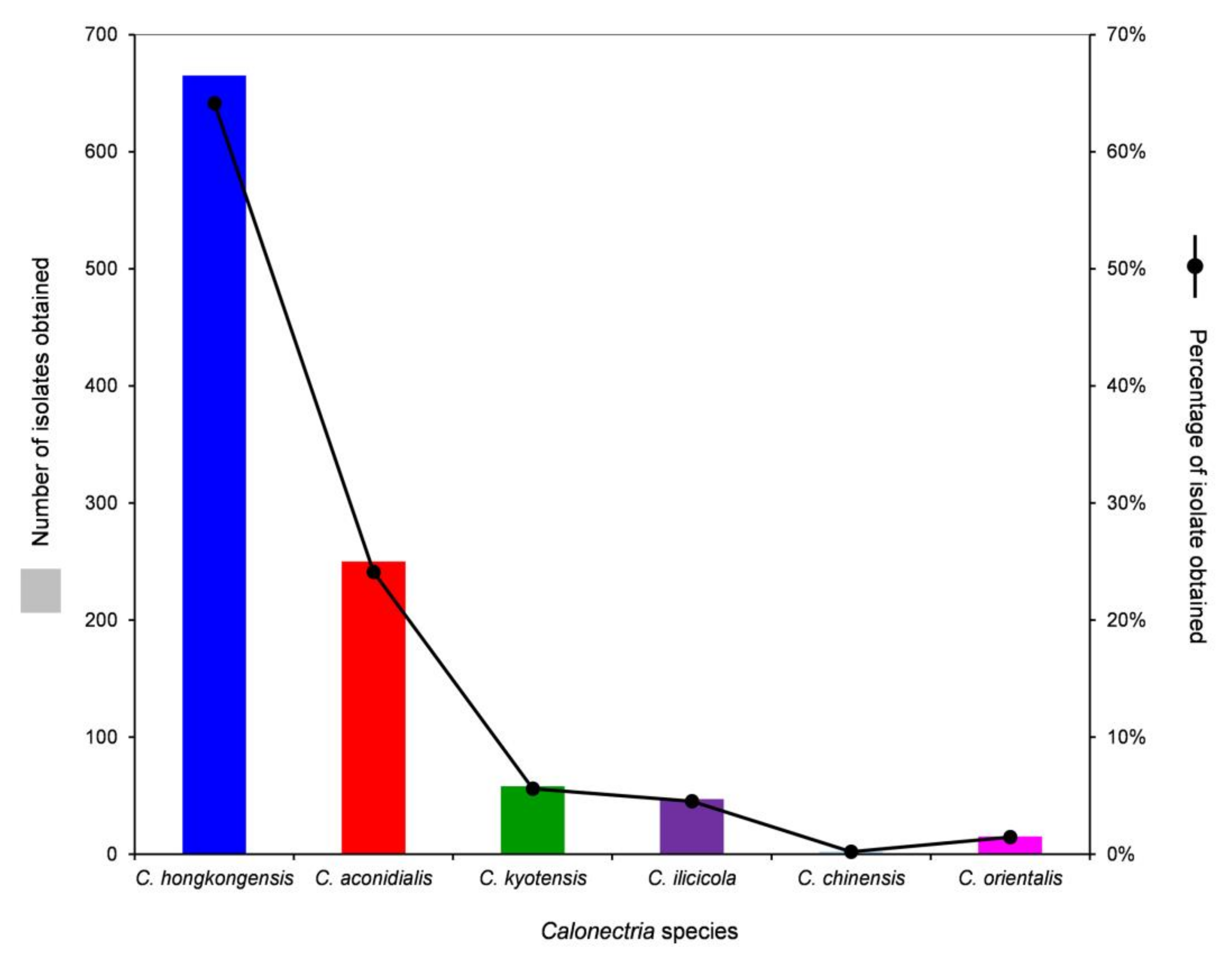
| All five soil layers | 665 | 250 | 58 | 47 | 2 | 15 | 1037 | |
| Percentage | 64.1% | 24.1% | 5.6% | 4.5% | 0.2% | 1.5% |
| Calonectria Species | Number of Genotypes Determined by tef1 and tub2 Gene Sequences | Genotype Determined by tef1 and tub2 Gene Sequences | Number of Isolates of Each Genotype | Number of isolates of Each Calonectria Species |
|---|---|---|---|---|
| C. hongkongensis | 11 | AA | 561 | 665 |
| AB | 1 | |||
| AC | 4 | |||
| AD | 7 | |||
| AE | 2 | |||
| AF | 20 | |||
| AG | 15 | |||
| AH | 4 | |||
| BA | 15 | |||
| CA | 5 | |||
| DA | 31 | |||
| C. aconidialis | 3 | AA | 156 | 250 |
| AB | 9 | |||
| AC | 85 | |||
| C. kyotensis | 3 | AA | 33 | 58 |
| AB | 19 | |||
| BA | 6 | |||
| C. ilicicola | 3 | AA | 26 | 47 |
| AB | 9 | |||
| BB | 12 | |||
| C. chinensis | 1 | AA | 2 | 2 |
| C. orientalis | 1 | AA | 15 | 15 |
| All six Calonectria species | 22 | 1037 | 1037 |
| Identity | Genotype 1 | Isolate No. 2 | Sampling Point No. 3 | Soil Layer | Sample and Isolate Information 4 | Collectors | GenBank Accession No. 5 | |||
|---|---|---|---|---|---|---|---|---|---|---|
| tef1 | tub2 | cmdA | his3 | |||||||
| C. aconidialis | AAAA | CSF20325 | 6 | 0–20 cm | 20200711-1-(3)_0–20 cm_A_R2_SC2 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167700 | OK168737 | OK169148 | OK169232 |
| C. aconidialis | AAAA | CSF21348 | 98 | 0–20 cm | 20200816-1-(6)_0–20 cm_A_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167855 | OK168892 | OK169151 | OK169235 |
| C. aconidialis | AACA | CSF20378 | 9 | 0–20 cm | 20200711-1-(6)_0–20 cm_A_R2_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167701 | OK168738 | OK169149 | OK169233 |
| C. aconidialis | AACA | CSF20447 | 11 | 0–20 cm | 20200715-1-(1)_0–20 cm_B_R2_SC2 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167704 | OK168741 | OK169150 | OK169234 |
| C. aconidialis | ABBA | CSF20985 6 | 68 | 20–40 cm | 20200811-1-(4)_0–40 cm_B_R1_SC3 | L.L. Liu, J.L. Han, and L.S. Sun | OK167856 | OK168893 | OK169152 | OK169236 |
| C. aconidialis | ABBA | CSF21262 | 93 | 20–40 cm | 20200816-1-(1)_0–40 cm_B_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167857 | OK168894 | OK169153 | OK169237 |
| C. aconidialis | ABBA | CSF21266 | 93 | 20–40 cm | 20200816-1-(1)_0–40 cm_B_R2_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK167861 | OK168898 | OK169154 | OK169238 |
| C. aconidialis | ABBA | CSF21349 | 98 | 0–20 cm | 20200816-1-(6)_0–20 cm_A_R2_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK167864 | OK168901 | OK169155 | OK169239 |
| C. aconidialis | ACAA | CSF20257 | 1 | 0–20 cm | 20200709-1-(1)_0–20 cm_A_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167865 | OK168902 | OK169156 | OK169240 |
| C. aconidialis | ACAA | CSF20323 6 | 6 | 0–20 cm | 20200711-1-(3)_0–20 cm_A_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167866 | OK168903 | OK169157 | OK169241 |
| C. aconidialis | ACAA | CSF20376 6 | 9 | 0–20 cm | 20200711-1-(6)_0–20 cm_A_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167868 | OK168905 | OK169158 | OK169242 |
| C. aconidialis | ACAA | CSF21346 | 98 | 0–20 cm | 20200816-1-(6)_0–20 cm_A_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167946 | OK168983 | OK169159 | OK169243 |
| C. chinensis | AAAA | CSF20756 6 | 52 | 0–20 cm | 20200809-1-(2)_0–20 cm_A_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK168055 | OK169092 | OK169184 | OK169268 |
| C. chinensis | AAAA | CSF20759 6 | 52 | 0–20 cm | 20200809-1-(2)_0–20 cm_A_R2_SC4 | L.L. Liu, J.L. Han, and L.S. Sun | OK168056 | OK169093 | OK169185 | OK169269 |
| C. hongkongensis | AAAA | CSF20258 | 1 | 0–20 cm | 20200709-1-(1)_0–20 cm_A_R1_SC2 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167035 | OK168072 | OK169109 | OK169194 |
| C. hongkongensis | AAAA | CSF20271 | 2 | 0–20 cm | 20200709-1-(2)_0–20 cm_A_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167044 | OK168081 | OK169110 | OK169195 |
| C. hongkongensis | AAAA | CSF20291 | 3 | 0–20 cm | 20200709-1-(3)_0–20 cm_A_R2_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167056 | OK168093 | OK169111 | OK169196 |
| C. hongkongensis | AAAA | CSF21370 | 100 | 0–20 cm | 20200816-1-(8)_0–20 cm_A_R2_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK167588 | OK168625 | OK169112 | OK169197 |
| C. hongkongensis | ABA- | CSF20758 | 52 | 0–20 cm | 20200809-1-(2)_0–20 cm_A_R2_SC3 | L.L. Liu, J.L. Han, and L.S. Sun | OK167596 | OK168633 | OK169113 | – 7 |
| C. hongkongensis | ACAA | CSF20524 | 17 | 0–20 cm | 20200715-1-(7)_0–20 cm_B_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167597 | OK168634 | OK169114 | OK169198 |
| C. hongkongensis | ACAA | CSF20525 | 17 | 0–20 cm | 20200715-1-(7)_0–20 cm_B_R1_SC2 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167598 | OK168635 | OK169115 | OK169199 |
| C. hongkongensis | ACAB | CSF21368 | 100 | 0–20 cm | 20200816-1-(8)_0–20 cm_A_R1_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK167599 | OK168636 | OK169116 | OK169200 |
| C. hongkongensis | ACAB | CSF21372 | 100 | 0–20 cm | 20200816-1-(8)_0–20 cm_B_R1_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK167600 | OK168637 | OK169117 | OK169201 |
| C. hongkongensis | ADAA | CSF20412 | 10 | 0–20 cm | 20200711-1-(7)_0–20 cm_B_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167601 | OK168638 | OK169118 | OK169202 |
| C. hongkongensis | ADAA | CSF20454 | 11 | 20–40 cm | 20200715-1-(1)_0–40 cm_A_R2_SC3 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167602 | OK168639 | OK169119 | OK169203 |
| C. hongkongensis | ADAA | CSF20834 | 60 | 0–20 cm | 20200810-1-(4)_0–20 cm_B_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167604 | OK168641 | OK169120 | OK169204 |
| C. hongkongensis | ADAA | CSF21304 | 96 | 0–20 cm | 20200816-1-(4)_0–20 cm_A_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167607 | OK168644 | OK169121 | OK169205 |
| C. hongkongensis | AEAA | CSF20923 | 65 | 0–20 cm | 20200811-1-(1)_0–20 cm_A_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167608 | OK168645 | OK169122 | OK169206 |
| C. hongkongensis | AEAA | CSF20924 6 | 65 | 0–20 cm | 20200811-1-(1)_0–20 cm_A_R1_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK167609 | OK168646 | OK169123 | OK169207 |
| C. hongkongensis | AFAA | CSF20259 | 1 | 0–20 cm | 20200709-1-(1)_0–20 cm_A_R2_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167610 | OK168647 | OK169124 | OK169208 |
| C. hongkongensis | AFAA | CSF20309 | 4 | 0–20 cm | 20200711-1-(1)_0–20 cm_A_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167611 | OK168648 | OK169125 | OK169209 |
| C. hongkongensis | AFAA | CSF20470 | 12 | 0–20 cm | 20200715-1-(2)_0–20 cm_A_R2_SC1 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167615 | OK168652 | OK169126 | OK169210 |
| C. hongkongensis | AFAA | CSF21233 | 90 | 0–20 cm | 20200815-1-(3)_0–20 cm_B_R2_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK167629 | OK168666 | OK169127 | OK169211 |
| C. hongkongensis | AGAA | CSF20380 | 9 | 0–20 cm | 20200711-1-(6)_0–20 cm_B_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167630 | OK168667 | OK169128 | OK169212 |
| C. hongkongensis | AGAA | CSF20441 | 11 | 0–20 cm | 20200715-1-(1)_0–20 cm_A_R1_SC2 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167631 | OK168668 | OK169129 | OK169213 |
| C. hongkongensis | AGAA | CSF20528 | 17 | 40–60 cm | 20200715-1-(7)_0–60 cm_A_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167632 | OK168669 | OK169130 | OK169214 |
| C. hongkongensis | AGAA | CSF21018 | 71 | 0–20 cm | 20200811-1-(7)_0–20 cm_B_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167644 | OK168681 | OK169131 | OK169215 |
| C. hongkongensis | AHAA | CSF20760 | 52 | 0–20 cm | 20200809-1-(2)_0–20 cm_B_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167645 | OK168682 | OK169132 | OK169216 |
| C. hongkongensis | AHAA | CSF20761 6 | 52 | 0–20 cm | 20200809-1-(2)_0–20 cm_B_R1_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK167646 | OK168683 | OK169133 | OK169217 |
| C. hongkongensis | AHAA | CSF21155 | 82 | 0–20 cm | 20200813-1-(2)_0–20 cm_B_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167647 | OK168684 | OK169134 | OK169218 |
| C. hongkongensis | AHAA | CSF21156 | 82 | 0–20 cm | 20200813-1-(2)_0–20 cm_B_R2_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK167648 | OK168685 | OK169135 | OK169219 |
| C. hongkongensis | BAAA | CSF20472 | 12 | 0–20 cm | 20200715-1-(2)_0–20 cm_B_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167649 | OK168686 | OK169136 | OK169220 |
| C. hongkongensis | BAAA | CSF20734 | 51 | 0–20 cm | 20200809-1-(1)_0–20 cm_A_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167652 | OK168689 | OK169137 | OK169221 |
| C. hongkongensis | BAAA | CSF21183 | 86 | 0–20 cm | 20200814-1-(2)_0–20 cm_B_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167657 | OK168694 | OK169138 | OK169222 |
| C. hongkongensis | BAAA | CSF21359 | 99 | 0–20 cm | 20200816-1-(7)_0–20 cm_A_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167660 | OK168697 | OK169139 | OK169223 |
| C. hongkongensis | CAAA | CSF20353 6 | 7 | 0–20 cm | 20200711-1-(4)_0–20 cm_A_R2_SC2 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167664 | OK168701 | OK169140 | OK169224 |
| C. hongkongensis | CAAA | CSF20358 | 7 | 20–40 cm | 20200711-1-(4)_0–40 cm_B_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167665 | OK168702 | OK169141 | OK169225 |
| C. hongkongensis | CAAA | CSF20359 | 7 | 20–40 cm | 20200711-1-(4)_0–40 cm_B_R1_SC2 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167666 | OK168703 | OK169142 | OK169226 |
| C. hongkongensis | CAAA | CSF20360 6 | 7 | 20–40 cm | 20200711-1-(4)_0–40 cm_B_R1_SC3 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167667 | OK168704 | OK169143 | OK169227 |
| C. hongkongensis | DAAA | CSF20334 | 6 | 20–40 cm | 20200711-1-(3)_0–40 cm_B_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167669 | OK168706 | OK169144 | OK169228 |
| C. hongkongensis | DAAA | CSF20383 6 | 9 | 0–20 cm | 20200711-1-(6)_0–20 cm_B_R2_SC2 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167673 | OK168710 | OK169145 | OK169229 |
| C. hongkongensis | DAAA | CSF20444 | 11 | 0–20 cm | 20200715-1-(1)_0–20 cm_B_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167678 | OK168715 | OK169146 | OK169230 |
| C. hongkongensis | DAAA | CSF21367 | 100 | 0–20 cm | 20200816-1-(8)_0–20 cm_A_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167699 | OK168736 | OK169147 | OK169231 |
| C. ilicicola | AAAB | CSF20594 | 29 | 0–20 cm | 20200727-1-(5)_0–20 cm_A_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK168008 | OK169045 | OK169172 | OK169256 |
| C. ilicicola | AAAB | CSF21126 | 80 | 20–40 cm | 20200812-1-(8)_0–40 cm_A_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK168010 | OK169047 | OK169173 | OK169257 |
| C. ilicicola | AAAB | CSF21219 | 89 | 0–20 cm | 20200815-1-(2)_0–20 cm_A_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK168014 | OK169051 | OK169174 | OK169258 |
| C. ilicicola | AAAB | CSF21310 6 | 96 | 20–40 cm | 20200816-1-(4)_0–40 cm_A_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK168016 | OK169053 | OK169175 | OK169259 |
| C. ilicicola | ABAA | CSF20618 6 | 32 | 0–20 cm | 20200729-1-(2)_0–20 cm_A_R1_SC1 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168034 | OK169071 | OK169176 | OK169260 |
| C. ilicicola | ABAA | CSF20620 | 32 | 0–20 cm | 20200729-1-(2)_0–20 cm_A_R2_SC1 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168036 | OK169073 | OK169177 | OK169261 |
| C. ilicicola | ABAA | CSF20624 | 32 | 20–40 cm | 20200729-1-(2)_0–40 cm_A_R1_SC1 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168038 | OK169075 | OK169178 | OK169262 |
| C. ilicicola | ABAA | CSF20703 | 45 | 0–20 cm | 20200731-1-(2)_0–20 cm_B_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK168042 | OK169079 | OK169179 | OK169263 |
| C. ilicicola | BBAA | CSF20853 | 61 | 20–40 cm | 20200810-1-(5)_0–40 cm_A_R1_SC8 | L.L. Liu, J.L. Han, and L.S. Sun | OK168043 | OK169080 | OK169180 | OK169264 |
| C. ilicicola | BBBA | CSF21052 6 | 74 | 0–20 cm | 20200812-1-(2)_0–20 cm_A_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK168044 | OK169081 | OK169181 | OK169265 |
| C. ilicicola | BBBA | CSF21198 | 87 | 0–20 cm | 20200814-1-(3)_0–20 cm_A_R2_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK168047 | OK169084 | OK169182 | OK169266 |
| C. ilicicola | BBBA | CSF21292 | 95 | 0–20 cm | 20200816-1-(3)_0–20 cm_A_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK168053 | OK169090 | OK169183 | OK169267 |
| C. kyotensis | AAAA | CSF20372 | 8 | 0–20 cm | 20200711-1-(5)_0–20 cm_B_R2_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167950 | OK168987 | OK169160 | OK169244 |
| C. kyotensis | AAAA | CSF20443 | 11 | 0–20 cm | 20200715-1-(1)_0–20 cm_A_R2_SC2 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167952 | OK168989 | OK169161 | OK169245 |
| C. kyotensis | AAAA | CSF21350 | 98 | 0–20 cm | 20200816-1-(6)_0–20 cm_B_R1_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167981 | OK169018 | OK169163 | OK169247 |
| C. kyotensis | AAAB | CSF20518 | 16 | 0–20 cm | 20200715-1-(6)_0–20 cm_B_R2_SC1 | S.F. Chen, L.L. Liu, J.L. Han, L.S. Sun, and W.W. Li | OK167953 | OK168990 | OK169162 | OK169246 |
| C. kyotensis | ABAA | CSF21191 6 | 86 | 40–60 cm | 20200814-1-(2)_0–60 cm_B_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK167998 | OK169035 | OK169167 | OK169251 |
| C. kyotensis | ABAB | CSF20260 | 1 | 0–20 cm | 20200709-1-(1)_0–20 cm_A_R2_SC2 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167983 | OK169020 | OK169164 | OK169248 |
| C. kyotensis | ABAB | CSF20432 | 10 | 40–60 cm | 20200711-1-(7)_0–60 cm_B_R2_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167988 | OK169025 | OK169166 | OK169250 |
| C. kyotensis | ABBA | CSF20338 | 6 | 20–40 cm | 20200711-1-(3)_0–40 cm_B_R2_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK167986 | OK169023 | OK169165 | OK169249 |
| C. kyotensis | BAAA | CSF20275 | 2 | 20–40 cm | 20200709-1-(2)_0–40 cm_A_R1_SC1 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK168002 | OK169039 | OK169168 | OK169252 |
| C. kyotensis | BAAA | CSF20276 6 | 2 | 20–40 cm | 20200709-1-(2)_0–40 cm_A_R1_SC2 | S.F. Chen, L.L. Liu, J.L. Han, Y Liu, and X.Y. Liang | OK168003 | OK169040 | OK169169 | OK169253 |
| C. kyotensis | BAAA | CSF21111 | 78 | 0–20 cm | 20200812-1-(6)_0–20 cm_B_R2_SC1 | L.L. Liu, J.L. Han, and L.S. Sun | OK168006 | OK169043 | OK169170 | OK169254 |
| C. kyotensis | BAAA | CSF21335 6 | 97 | 0–20 cm | 20200816-1-(5)_0–20 cm_A_R1_SC2 | L.L. Liu, J.L. Han, and L.S. Sun | OK168007 | OK169044 | OK169171 | OK169255 |
| C. orientalis | AAAA | CSF20602 | 31 | 40–60 cm | 20200729-1-(1)_0–60 cm_A_R1_SC1 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168057 | OK169094 | OK169186 | OK169270 |
| C. orientalis | AAAA | CSF20603 | 31 | 40–60 cm | 20200729-1-(1)_0–60 cm_A_R1_SC2 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168058 | OK169095 | OK169187 | OK169271 |
| C. orientalis | AAAA | CSF20606 | 31 | 40–60 cm | 20200729-1-(1)_0–60 cm_B_R1_SC1 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168061 | OK169098 | OK169188 | OK169272 |
| C. orientalis | AAAA | CSF20607 | 31 | 40–60 cm | 20200729-1-(1)_0–60 cm_B_R1_SC2 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168062 | OK169099 | OK169189 | OK169273 |
| C. orientalis | AAAA | CSF20610 | 31 | 40–60 cm | 20200729-1-(1)_0–60 cm_B_R2_SC1 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168064 | OK169101 | OK169190 | OK169274 |
| C. orientalis | AAAA | CSF20611 | 31 | 40–60 cm | 20200729-1-(1)_0–60 cm_B_R2_SC2 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168065 | OK169102 | OK169191 | OK169275 |
| C. orientalis | AAAA | CSF20614 6 | 31 | 60–80 cm | 20200729-1-(1)_0–80 cm_B_R1_SC1 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168068 | OK169105 | OK169192 | OK169276 |
| C. orientalis | AAAA | CSF20615 | 31 | 60–80 cm | 20200729-1-(1)_0–80 cm_B_R1_SC2 | L.L. Liu, J.L. Han, L.S. Sun, Y Liu, and X.Y. Liang | OK168069 | OK169106 | OK169193 | OK169277 |
| Species Code 1 | Species | Isolates No. 2,3 | Other Collection Number 3 | Hosts | Area of Occurrence | Collector | GenBank Accession No. 4 | References | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| tef1 | tub2 | cmdA | his3 | ||||||||
| Species in Calonectria kyotensis species complex | |||||||||||
| B4 | C. aconidialis | CMW 35174T | CBS 136086; CERC 1850 | Soil in Eucalyptus plantation | HaiNan, China | X. Mou and S.F. Chen | MT412695 | OK357463 | MT335165 | MT335404 | [5,9] |
| CMW 35384 | CBS 136091; CERC 1886 | Soil in Eucalyptus plantation | HaiNan, China | X. Mou and S.F. Chen | MT412696 | OK357464 | MT335166 | MT335405 | [5,9] | ||
| B5 | C. aeknauliensis | CMW 48253T | CBS 143559 | Soil in Eucalyptus plantation | Aek Nauli, North Sumatra, Indonesia | M.J. Wingfield | MT412710 | OK357465 | MT335180 | MT335419 | [5,12] |
| CMW 48254 | CBS 143560 | Soil in Eucalyptus plantation | Aek Nauli, North Sumatra, Indonesia | M.J. Wingfield | MT412711 | OK357466 | MT335181 | MT335420 | [5,12] | ||
| B8 | C. asiatica | CBS 114073T | CMW 23782; CPC 3900 | Debris leaf litter | Prathet Thai, Thailand | N.L. Hywel-Jones | AY725705 | AY725616 | AY725741 | AY725658 | [29,49] |
| B17 | C. brassicicola | CBS 112841T | CMW 51206; CPC 4552 | Soil at Brassica sp. | Indonesia | M.J. Wingfield | KX784689 | KX784619 | KX784561 | N/A 5 | [30] |
| B19 | C. bumicola | CMW 48257T | CBS 143575 | Soil in Eucalyptus plantation | Aek Nauli, North Sumatra, Indonesia | M.J. Wingfield | MT412736 | OK357467 | MT335205 | MT335445 | [5,12] |
| B20 | C. canadiana | CMW 23673T | CBS 110817; STE-U 499 | Picea sp. | Canada | S. Greifenhagen | MT412737 | MT412958 | MT335206 | MT335446 | [1,5,17,50] |
| CERC 8952 | – | Soil | HeNan, China | S.F. Chen | MT412821 | MT413035 | MT335290 | MT335530 | [5,36] | ||
| B23 | C. chinensis | CMW 23674T | CBS 114827; CPC 4101 | Soil | Hong Kong, China | E.C.Y. Liew | MT412751 | MT412972 | MT335220 | MT335460 | [5,29,49] |
| CMW 30986 | CBS 112744; CPC 4104 | Soil | Hong Kong, China | E.C.Y. Liew | MT412752 | MT412973 | MT335221 | MT335461 | [5,29,49] | ||
| B26 | C. cochinchinensis | CMW 49915T | CBS 143567 | Soil in Hevea brasiliensis plantation | Duong Minh Chau, Tay Ninh, Vietnam | N.Q. Pham, Q.N. Dang, and T.Q. Pham | MT412756 | MT412977 | MT335225 | MT335465 | [5,12] |
| CMW 47186 | CBS 143568 | Soil in Acacia auriculiformis plantation | Song May, Dong Nai, Vietnam | N.Q. Pham and T.Q. Pham | MT412757 | MT412978 | MT335226 | MT335466 | [5,12] | ||
| B29 | C. colombiensis | CMW 23676T | CBS 112220; CPC 723 | Soil in E. grandis trees | La Selva, Colombia | M.J. Wingfield | MT412759 | MT412980 | MT335228 | MT335468 | [5,49] |
| CMW 30985 | CBS 112221; CPC 724 | Soil in E. grandis trees | La Selva, Colombia | M.J. Wingfield | MT412760 | MT412981 | MT335229 | MT335469 | [5,49] | ||
| B31 | C. curvispora | CMW 23693T | CBS 116159; CPC 765 | Soil | Tamatave, Madagascar | P.W. Crous | MT412763 | OK357468 | MT335232 | MT335472 | [1,5,9,29,51] |
| CMW 48245 | CBS 143565 | Soil in Eucalyptus plantation | Aek Nauli, North Sumatra, Indonesia | M.J. Wingfield | MT412764 | N/A | MT335233 | MT335473 | [5,12] | ||
| B46 | C. heveicola | CMW 49913T | CBS 143570 | Soil in H. brasiliensis plantation | Bau Bang, Binh Duong, Vietnam | N.Q. Pham, Q.N. Dang, and T.Q. Pham | MT412786 | MT413004 | MT335255 | MT335495 | [5,12] |
| CMW 49928 | CBS 143571 | Soil | Bu Gia Map National Park, Binh Phuoc, Vietnam | N.Q. Pham, Q.N. Dang, and T.Q. Pham | MT412811 | MT413025 | MT335280 | MT335520 | [5,12] | ||
| B48 | C. hongkongensis | CBS 114828T | CMW 51217; CPC 4670 | Soil | Hong Kong, China | M.J. Wingfield | MT412789 | MT413007 | MT335258 | MT335498 | [5,49] |
| CERC 3570 | CMW 47271 | Soil in Eucalyptus plantation | BeiHai, Guangxi, China | S.F. Chen, J.Q. Li, and G.Q. Li | MT412791 | MT413009 | MT335260 | MT335500 | [5,11] | ||
| B51 | C. ilicicola | CMW 30998T | CBS 190.50; IMI 299389; STE-U 2482 | Solanum tuberosum | Bogor, Java, Indonesia | K.B. Boedijn and J. Reitsma | MT412797 | OK357469 | MT335266 | MT335506 | [1,5,29,52] |
| B52 | C. indonesiae | CMW 23683T | CBS 112823; CPC 4508 | Syzygium aromaticum | Warambunga, Indonesia | M.J. Wingfield | MT412798 | MT413015 | MT335267 | MT335507 | [5,49] |
| CBS 112840 | CMW 51205; CPC 4554 | S. aromaticum | Warambunga, Indonesia | M.J. Wingfield | MT412799 | MT413016 | MT335268 | MT335508 | [5,49] | ||
| B55 | C. kyotensis | CBS 114525T | ATCC 18834; CMW 51824; CPC 2367 | Robinia pseudoacacia | Japan | T. Terashita | MT412802 | MT413019 | MT335271 | MT335511 | [1,5,30,53] |
| CBS 114550 | CMW 51825; CPC 2351 | Soil | China | M.J. Wingfield | MT412777 | MT412995 | MT335246 | MT335486 | [5,30] | ||
| B57 | C. lantauensis | CERC 3302T | CBS 142888; CMW 47252 | Soil | LiDao, Hong Kong, China | M.J. Wingfield and S.F. Chen | MT412803 | OK357470 | MT335272 | MT335512 | [5,11] |
| CERC 3301 | CBS 142887; CMW 47251 | Soil | LiDao, Hong Kong, China | M.J. Wingfield and S.F. Chen | MT412804 | OK357471 | MT335273 | MT335513 | [5,11] | ||
| B58 | C. lateralis | CMW 31412T | CBS 136629 | Soil in Eucalyptus plantation | GuangXi, China | X. Zhou, G. Zhao, and F. Han | MT412805 | MT413020 | MT335274 | MT335514 | [5,9] |
| B66 | C. malesiana | CMW 23687T | CBS 112752; CPC 4223 | Soil | Northern Sumatra, Indonesia | M.J. Wingfield | MT412817 | MT413031 | MT335286 | MT335526 | [5,49] |
| CBS 112710 | CMW 51199; CPC 3899 | Leaf litter | Prathet, Thailand | N.L. Hywel-Jones | MT412818 | MT413032 | MT335287 | MT335527 | [5,49] | ||
| B80 | C. pacifica | CMW 16726T | A1568; CBS 109063; IMI 354528; STE-U 2534 | Araucaria heterophylla | Hawaii, USA | M. Aragaki | MT412842 | OK357472 | MT335311 | MT335551 | [1,5,49,50] |
| CMW 30988 | CBS 114038 | Ipomoea aquatica | Auckland, New Zealand | C.F. Hill | MT412843 | OK357473 | MT335312 | MT335552 | [1,5,29,49] | ||
| B86 | C. penicilloides | CMW 23696T | CBS 174.55; STE-U 2388 | Prunus sp. | Hatizyo Island, Japan | M. Ookubu | MT412869 | MT413081 | MT335338 | MT335578 | [1,5,54] |
| B112 | C. sumatrensis | CMW 23698T | CBS 112829; CPC 4518 | Soil | Northern Sumatra, Indonesia | M.J. Wingfield | MT412913 | OK357474 | MT335382 | MT335622 | [5,49] |
| CMW 30987 | CBS 112934; CPC 4516 | Soil | Northern Sumatra, Indonesia | M.J. Wingfield | MT412914 | OK357475 | MT335383 | MT335623 | [5,49] | ||
| B113 | C. syzygiicola | CBS 112831T | CMW 51204; CPC 4511 | Syzygium aromaticum | Sumatra, Indonesia | M.J. Wingfield | KX784736 | KX784663 | N/A | N/A | [30] |
| B116 | C. uniseptata | CBS 413.67T | CMW 23678; CPC 2391; IMI 299577 | Paphiopedilum callosum | Celle, Germany | W. Gerlach | GQ267307 | GQ267208 | GQ267379 | GQ267248 | [30] |
| B120 | C. yunnanensis | CERC 5339T | CBS 142897; CMW 47644 | Soil in Eucalyptus plantation | YunNan, China | S.F. Chen and J.Q. Li | MT412927 | MT413134 | MT335396 | MT335636 | [5,11] |
| CERC 5337 | CBS 142895; CMW 47642 | Soil in Eucalyptus plantation | YunNan, China | S.F. Chen and J.Q. Li | MT412928 | MT413135 | MT335397 | MT335637 | [5,11] | ||
| B124 | C. singaporensis | CBS 146715T | MUCL 048320 | leaf litter submerged in a small stream | Mac Ritchie Reservoir, Singapore | C. Decock | MW890086 | MW890124 | MW890042 | MW890055 | [34] |
| CBS 146713 | MUCL 048171 | leaf litter submerged in a small stream | Mac Ritchie Reservoir, Singapore | C. Decock | MW890084 | MW890123 | MW890040 | MW890053 | [34] | ||
| Species in Calonectria brassicae species complex | |||||||||||
| B12 | C. brachiatica | CMW 25298T | CBS 123700 | Pinus maximinoi | Buga, Colombia | M.J. Wingfield | MT412726 | MT412948 | MT335195 | MT335435 | [5,7] |
| CMW 25302 | – | Pinus ecunumanii | Buga, Colombia | M.J. Wingfield | MT412727 | MT412949 | MT335196 | MT335436 | [5,7] | ||
| B16 | C. brassicae | CBS 111869T | CPC 2409 | Argyreia splendens | Indonesia | F. Bugnicourt | MT412733 | MT412955 | MT335202 | MT335442 | [1,5,29,30] |
| B25 | C. clavata | CMW 23690T | ATCC 66389; CBS 114557; CPC 2536; P078-1543 | Callistemon viminalis | Lake Placid, Florida, USA | C.P. Seymour and E.L. Barnard | MT412754 | MT412975 | MT335223 | MT335463 | [1,5,29,55] |
| CMW 30994 | CBS 114666; CPC 2537; P078-1261 | Root debris in peat | Lee County, Florida, USA | D. Ferrin | MT412755 | MT412976 | MT335224 | MT335464 | [1,5,29,55] | ||
| B34 | C. duoramosa | CBS 134656T | – | Soil in tropical rainforest | Monte Dourado, Pará, Brazil | R.F. Alfenas | KM395853 | KM395940 | KM396027 | KM396110 | [10] |
| LPF453 | – | Soil in Eucalyptus plantation | Monte Dourado, Pará, Brazil | R.F. Alfenas | KM395854 | KM395941 | KM396028 | KM396111 | [10] | ||
| B35 | C. ecuadorae | CMW 23677T | CBS 111406; CPC 1635 | Soil | Ecuador | M.J. Wingfield | MT412773 | MT412991 | MT335242 | MT335482 | [5,29,56] |
| CBS 111706 | CMW 51821; CPC 1636 | Soil | Ecuador | M.J. Wingfield | MT412771 | MT412989 | MT335240 | MT335480 | [5,31] | ||
| B43 | C. gracilis | CBS 111807T | AR2677; CMW 51189; STE-U 2634 | Manilkara zapota | Pará, Brazil | F. Carneiro de Albuquerque | GQ267323 | AF232858 | GQ267407 | DQ190646 | [1,30,31,56,57] |
| CBS 111284 | CMW 51175; CPC 1483 | Soil | Imbrapa, Brazil | P.W. Crous | GQ267324 | DQ190567 | GQ267408 | DQ190647 | [1,30,31,56,57] | ||
| B77 | C. octoramosa | CBS 111423T | CMW 51819; CPC 1650 | Soil | Ecuador | M.J. Wingfield | MT412834 | MT413048 | MT335303 | MT335543 | [5,31] |
| B78 | C. orientalis | CMW 20291T | CBS 125260 | Soil | Langam, Indonesia | M.J. Wingfield | MT412835 | MT413049 | MT335304 | MT335544 | [5,29] |
| CMW 20273 | CBS 125259 | Soil | Teso East, Indonesia | M.J. Wingfield | MT412836 | MT413050 | MT335305 | MT335545 | [5,29] | ||
| B82 | C. paraensis | CBS 134669T | LPF430 | Soil in Eucalyptus plantation | Monte Dourado, Pará, Brazil | R.F. Alfenas | KM395837 | KM395924 | KM396011 | KM396094 | [10] |
| LPF429 | – | Soil in tropical rainforest | Monte Dourado, Pará, Brazil | R.F. Alfenas | KM395841 | KM395928 | KM396015 | KM396098 | [10] | ||
| B83 | C. parvispora | CBS 111465T | CPC 1902 | Soil | Brazil | A.C. Alfenas | MT412845 | MT413057 | MT335314 | MT335554 | [5,31] |
| CMW 30981 | CBS 111478; CPC 1921 | Soil | Brazil | A.C. Alfenas | MT412844 | MT413056 | MT335313 | MT335553 | [5,29,30] | ||
| B84 | C. pauciphialidica | CMW 30980T | CBS 111394; CPC 1628 | Soil | Ecuador | M.J. Wingfield | MT412846 | MT413058 | MT335315 | MT335555 | [5,29,56] |
| B88 | C. pini | CMW 31209T | CBS 123698 | Pinus patula | Buga, Valle del Cauca, Colombia | C.A. Rodas | MT412870 | MT413082 | MT335339 | MT335579 | [5,29] |
| CBS 125523 | CMW 31210 | Pinus patula | Buga, Valle del Cauca, Colombia | C.A. Rodas | GQ267345 | GQ267225 | GQ267437 | GQ267274 | [29] | ||
| B91 | C. pseudobrassicae | CBS 134662T | LPF280 | Soil in Eucalyptus plantation | Santana, Pará, Brazil | A.C. Alfenas | KM395849 | KM395936 | KM396023 | KM396106 | [10] |
| CBS 134661 | LPF260 | Soil in Eucalyptus plantation | Santana, Pará, Brazil | A.C. Alfenas | KM395848 | KM395935 | KM396022 | KM396105 | [10] | ||
| B92 | C. pseudoecuadoriae | CBS 111402T | CMW 51179; CPC 1639 | Soil | Ecuador | M.J. Wingfield | KX784723 | KX784652 | KX784589 | N/A | [30,31] |
| B105 | C. quinqueramosa | CBS 134654T | LPF065 | Soil in Eucalyptus plantation | Monte Dourado, Pará, Brazil | R.F. Alfenas | KM395855 | KM395942 | KM396029 | KM396112 | [10] |
| CBS 134655 | LPF281 | Soil in Eucalyptus plantation | Santana, Pará, Brazil | A.C. Alfenas | KM395856 | KM395943 | KM396030 | KM396113 | [10] | ||
| B107 | C. robigophila | CBS 134652T | LPF192 | Eucalyptus sp. leaf | Açailandia, Maranhao, Brazil | R.F. Alfenas | KM395850 | KM395937 | KM396024 | KM396107 | [10] |
| CBS 134653 | LPF193 | Eucalyptus sp. leaf | Açailandia, Maranhao, Brazil | R.F. Alfenas | KM395851 | KM395938 | KM396025 | KM396108 | [10] | ||
| Outgroups | |||||||||||
| Curvicladiella cignea | CBS 109167T | CPC 1595; MUCL 40269 | Decaying leaf | French Guiana | C. Decock | KM231867 | KM232002 | KM231287 | KM231461 | [56,58,59] | |
| CBS 109168 | CPC 1594; MUCL 40268 | Decaying seed | French Guiana | C. Decock | KM231868 | KM232003 | KM231286 | KM231460 | [56,58,59] | ||
| Dataset | No. of Taxa | No. of bp 1 | Maximum Parsimony | ||||||
|---|---|---|---|---|---|---|---|---|---|
| PIC 2 | No. of Trees | Tree Length | CI 3 | RI 4 | RC 5 | HI 6 | |||
| tef1 | 157 | 522 | 241 | 110 | 588 | 0.697 | 0.973 | 0.678 | 0.303 |
| tub2 | 156 | 597 | 256 | 1000 | 694 | 0.635 | 0.967 | 0.614 | 0.365 |
| cmdA | 156 | 697 | 277 | 1000 | 617 | 0.676 | 0.969 | 0.655 | 0.324 |
| his3 | 153 | 478 | 166 | 973 | 602 | 0.570 | 0.960 | 0.547 | 0.430 |
| tef1/tub2/cmdA/his3 | 157 | 2303 | 944 | 150 | 2671 | 0.609 | 0.962 | 0.586 | 0.391 |
| Dataset | Maximum likelihood | ||||||||
| Subst. mode 7 | NST 8 | Rate matrix | Rates | ||||||
| tef1 | TIM2+G | 6 | 1.8670 | 3.4436 | 1.8670 | 1.0000 | 5.0336 | Gamma | |
| tub2 | TPM3uf+I+G | 6 | 1.4137 | 4.7965 | 1.0000 | 1.4137 | 4.7965 | Gamma | |
| cmdA | TrN+G | 6 | 1.0000 | 3.5934 | 1.0000 | 1.0000 | 7.2024 | Gamma | |
| his3 | GTR+I+G | 6 | 2.5191 | 8.8466 | 5.6820 | 2.1055 | 15.5239 | Gamma | |
| tef1/tub2/cmdA/his3 | GTR+I+G | 6 | 1.5966 | 4.2868 | 1.3927 | 0.9904 | 5.5003 | Gamma | |
| Species | Isolate/Species | Macroconidia (L × W) 1,2,3 | Macroconidia Average (L × W) 1,2 | Macroconidia Septation | Vesicle Width 1,2,3 | Vesicle Width Average 1 |
|---|---|---|---|---|---|---|
| C. aconidialis | Isolate CSF20323 (this study) | (35–)39.5–45.5(–48) × (4–)4–4.5(–5) | 42.5 × 4.5 | 1 | (3.5–)4.5–6(–6.5) | 5 |
| Isolate CSF20376 (this study) | (34.5–)38.5–45(–47.5) × (4–)4.5–5(–5.5) | 41.5 × 4.5 | 1 | (4–)4.5–11(–13) | 8 | |
| Isolate CSF20985 (this study) | (41–)46.5–51.5(–54) × (4–)4.5–5(–5.5) | 49 × 5 | 1 | (4.5–)5–6.5(–9.5) | 6 | |
| Species (this study) | (34.5–)40–48.5(–54) × (4–)4.5–5(–5.5) | 44.5 × 4.5 | 1 | (3.5–)4–8.5(–13) | 6 | |
| Species [9] | N/A 4 | N/A | N/A | N/A | N/A | |
| C. chinensis | Isolate CSF20756 (this study) | (35.5–)40–45(–49) × (3.5–)4–4.5(–4.5) | 42.5 × 4 | 1 | (3.5–)3.5–9(–11.5) | 6.5 |
| Isolate CSF20759 (this study) | (34.5–)37.5–43(–46) × (3.5–)4–4.5(–5) | 40.5 × 4 | 1 | (3–)5–10.5(–12) | 8 | |
| Species (this study) | (34.5–)38.5–44(–49) × (3.5–)4–4.5(–5) | 41.5 × 4 | 1 | (3–)4–10(–12) | 7 | |
| Species [49] | (38–)41–48(–56) × (3.5–)4(–4.5) | 45 × 4 | 1 | 6–9 | N/A | |
| C. hongkongensis | Isolate CSF20353 (this study) | (33.5–)36–42(–48) × (3.5–)4–4.5(–4.5) | 39 × 4 | 1 | (4–)5–8.5(–10.5) | 6.5 |
| Isolate CSF20360 (this study) | (34–)35.5–40(–43.5) × (3.5–)4–4.5(–5) | 37.5 × 4 | 1 | (4.5–)5.5–9(–11) | 7.5 | |
| Isolate CSF20383 (this study) | (37.5–)42.5–48(–50.5) × (4–)4–4.5(–5) | 45.5 × 4.5 | 1 | (4–)6–10.5(–11) | 8.5 | |
| Isolate CSF20761 (this study) | (32–)34.5–39.5(–43) × (3.5–)3.5–4(–4.5) | 37 × 4 | 1 | (4–)5.5–8(–9.5) | 6.5 | |
| Isolate CSF20924 (this study) | (35–)37.5–44(–45.5) × (3.5–)4–4.5(–5) | 40.5 × 4 | 1 | (6–)9–13(–14.5) | 11 | |
| Species (this study) | (32–)36–44(–50.5) × (3.5–)4–4.5(–5) | 40 × 4 | 1 | (4–)5.5–10.5(–14.5) | 8 | |
| Species [49] | (38–)45–48(–53) × 4(–4.5) | 46.5 × 4 | 1 | 8–14 | N/A | |
| C. ilicicola | Isolate CSF20618 (this study) | (52.5–)56.5–66(–71.5) × (6–)6.5–7.5(–8) | 61.5 × 7 | 1(–3) | (8–)9–11(–11.5) | 10 |
| Isolate CSF21052 (this study) | (31–)50.5–69(–78) × (3–)5–7(–7.5) | 59.5 × 6 | 1(–3) | (3.5–)5–8(–11) | 6.5 | |
| Isolate CSF21310 (this study) | (50–)55–62.5(–67) × (5.5–)6–7(–7.5) | 58.5 × 6.5 | (1–)3 | (4–)6.5–10(–11.5) | 8.5 | |
| Species (this study) | (31–)53.5–66(–78) × (3–)6–7(–8) | 60 × 6.5 | 1(–3) | (3.5–)6–10(–11.5) | 8 | |
| Species [1] | (45–)70–82(–90) × (4–)5–6.5(–7) | 62 × 6 | (1–)3 | (6–)7–10(–12) | N/A | |
| C. kyotensis | Isolate CSF20276 (this study) | (33.5–)36.5–44(–51) × (3.5–)4–4.5(–4.5) | 40.5 × 4 | 1 | (6.5–)8.5–11.5(–12.5) | 10 |
| Isolate CSF21191(this study) | (29.5–)32.5–38.5(–42.5) × (3.5–)4–4.5(–5) | 35.5 × 4 | 1 | (5–)7.5–10.5(–11.5) | 9 | |
| Isolate CSF21335 (this study) | (32–)35.5–40(–43) × (3.5–)4–4.5(–5) | 38 × 4 | 1 | (5–)8–10(–11) | 9 | |
| Species (this study) | (29.5–)34.5–41.5(–51) × (3.5–)4–4.5(–5) | 38 × 4 | 1 | (5–)7.5–10.5(–12.5) | 9 | |
| Species [1] | (35–)45–50(–55) × 3–4(–5) | 40 × 3.5 | 1 | 6–12 | N/A | |
| C. orientalis | Isolate CSF20614 (this study) | (30.5–)35–40(–43.5) × (4.5–)5–5.5(–5.5) | 37.5 × 5 | 1 | (3–)4–6.5(–7.5) | 5 |
| Species [29] | (43–)46–50(–53) × 4(–5) | 48 × 4 | 1 | 5–10 | N/A |
| Calonectria Species | Soil Layer | Genotype Determined by tef1 Gene Sequences | Number of Isolates Based on tef1 Genotype | Genotype Determined by tub2 Gene Sequence | Number of Isolates Based on tub2 Genotype | Genotype Determined by tef1 and tub2 Gene Sequences | Number of Isolates Based on tef1 and tub2 Genotype | Number of Isolates in Each Soil Layer for Each Species |
|---|---|---|---|---|---|---|---|---|
| C. hongkongensis | 0–20 cm | A | 346 | A | 337 | AA | 310 | 373 |
| B | 15 | B | 1 | AB | 1 | |||
| C | 1 | C | 4 | AC | 4 | |||
| D | 11 | D | 5 | AD | 5 | |||
| E | 2 | AE | 2 | |||||
| F | 9 | AF | 9 | |||||
| G | 11 | AG | 11 | |||||
| H | 4 | AH | 4 | |||||
| BA | 15 | |||||||
| CA | 1 | |||||||
| DA | 11 | |||||||
| 20–40 cm | A | 186 | A | 197 | AA | 180 | 203 | |
| C | 4 | D | 2 | AD | 2 | |||
| D | 13 | F | 4 | AF | 4 | |||
| CA | 4 | |||||||
| DA | 13 | |||||||
| 40–60 cm | A | 58 | A | 50 | AA | 47 | 61 | |
| D | 3 | F | 7 | AF | 7 | |||
| G | 4 | AG | 4 | |||||
| DA | 3 | |||||||
| 60–80 cm | A | 8 | A | 8 | AA | 8 | 8 | |
| 80–100 cm | A | 16 | A | 20 | AA | 16 | 20 | |
| D | 4 | DA | 4 | |||||
| C. aconidialis | 0–20 cm | A | 140 | A | 98 | AA | 98 | 140 |
| B | 1 | AB | 1 | |||||
| C | 41 | AC | 41 | |||||
| 20–40 cm | A | 74 | A | 40 | AA | 40 | 74 | |
| B | 8 | AB | 8 | |||||
| C | 26 | AC | 26 | |||||
| 40–60 cm | A | 20 | A | 6 | AA | 6 | 20 | |
| C | 14 | AC | 14 | |||||
| 60–80 cm | A | 8 | A | 8 | AA | 8 | 8 | |
| 80–100 cm | A | 8 | A | 4 | AA | 4 | 8 | |
| C | 4 | AC | 4 | |||||
| C. kyotensis | 0–20 cm | A | 31 | A | 27 | AA | 25 | 33 |
| B | 2 | B | 6 | AB | 6 | |||
| BA | 2 | |||||||
| 20–40 cm | A | 10 | A | 12 | AA | 8 | 14 | |
| B | 4 | B | 2 | AB | 2 | |||
| BA | 4 | |||||||
| 40–60 cm | A | 7 | B | 7 | AB | 7 | 7 | |
| 60–80 cm | A | 4 | B | 4 | AB | 4 | 4 | |
| 80–100 cm | – | – | – | – | – | – | 0 | |
| C. ilicicola | 0–20 cm | A | 9 | A | 4 | AA | 4 | 16 |
| B | 7 | B | 12 | AB | 5 | |||
| BB | 7 | |||||||
| 20–40 cm | A | 18 | A | 14 | AA | 14 | 19 | |
| B | 1 | B | 5 | AB | 4 | |||
| BB | 1 | |||||||
| 40–60 cm | A | 8 | A | 8 | AA | 8 | 8 | |
| 60–80 cm | B | 4 | B | 4 | BB | 4 | 4 | |
| 80–100 cm | – | – | – | – | – | – | 0 | |
| C. chinensis | 0–20 cm | A | 2 | A | 2 | AA | 2 | 2 |
| 20–40 cm | – | – | – | – | – | – | 0 | |
| 40–60 cm | – | – | – | – | – | – | 0 | |
| 60–80 cm | – | – | – | – | – | – | 0 | |
| 80–100 cm | – | – | – | – | – | – | 0 | |
| C. orientalis | 0–20 cm | – | – | – | – | – | – | 0 |
| 20–40 cm | – | – | – | – | – | – | 0 | |
| 40–60 cm | A | 11 | A | 11 | AA | 11 | 11 | |
| 60–80 cm | A | 4 | A | 4 | AA | 4 | 4 | |
| 80–100 cm | – | – | – | – | – | – | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Wu, W.; Chen, S. Species Diversity and Distribution Characteristics of Calonectria in Five Soil Layers in a Eucalyptus Plantation. J. Fungi 2021, 7, 857. https://doi.org/10.3390/jof7100857
Liu L, Wu W, Chen S. Species Diversity and Distribution Characteristics of Calonectria in Five Soil Layers in a Eucalyptus Plantation. Journal of Fungi. 2021; 7(10):857. https://doi.org/10.3390/jof7100857
Chicago/Turabian StyleLiu, LingLing, WenXia Wu, and ShuaiFei Chen. 2021. "Species Diversity and Distribution Characteristics of Calonectria in Five Soil Layers in a Eucalyptus Plantation" Journal of Fungi 7, no. 10: 857. https://doi.org/10.3390/jof7100857