
Chronic Diseases Associated with Malassezia Yeast

1
Aix Marseille Université, Institut de Recherche pour le Développement (IRD), Assistance Publique-Hôpitaux de Marseille (AP-HM), Service de Santé des Armées (SSA), VITROME: Vecteurs—Infections Tropicales et Méditerranéennes, 19-21 Boulevard Jean Moulin, 13005 Marseille, France
2
IHU Méditerranée Infection, 19-21 Boulevard Jean Moulin, 13005 Marseille, France
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(10), 855; https://doi.org/10.3390/jof7100855
Submission received: 21 September 2021
/
Revised: 4 October 2021
/
Accepted: 6 October 2021
/
Published: 12 October 2021
(This article belongs to the Topic Novel Concepts and Approaches in Neglected, Emerging and Re-Emerging Pathogens: From Environment to Patient)
Abstract
:Malassezia are a lipid-dependent basidiomycetous yeast of the normal skin microbiome, although Malassezia DNA has been recently detected in other body sites and has been associated with certain chronic human diseases. This new perspective raises many questions. Are these yeasts truly present in the investigated body site or were they contaminated by other body sites, adjacent or not? Does this DNA contamination come from living or dead yeast? If these yeasts are alive, do they belong to the resident mycobiota or are they transient colonizers which are not permanently established within these niches? Finally, are these yeasts associated with certain chronic diseases or not? In an attempt to shed light on this knowledge gap, we critically reviewed the 31 published studies focusing on the association of Malassezia spp. with chronic human diseases, including psoriasis, atopic dermatitis (AD), chronic rhinosinusitis (CRS), asthma, cystic fibrosis (CF), HIV infection, inflammatory bowel disease (IBD), colorectal cancer (CRC), and neurodegenerative diseases.
1. Introduction
Malassezia is a genus of fungi which consists of lipid-dependent basidiomycetous yeasts and forms part of the normal cutaneous microbiota of humans and other animals [1,2,3,4]. Although they are a human skin commensal, Malassezia species are involved, under certain circumstances, in various skin diseases including pityriasis versicolor, seborrheic dermatitis, folliculitis and dandruff [5]. Infrequently, Malassezia yeasts are involved in other dermatological diseases, such as confluent and reticulated papillomatosis and onychomycosis [6]. The association of Malassezia with psoriasis and atopic dermatitis has been mentioned and remains a matter of discussion. They have been also involved in fungemia, in immunocompromised and/or neonatal patients, especially those receiving lipidic parenteral nutrition [7,8,9]. In the last decade, the development of culture-independent tools such as high-throughput sequencing has expanded the knowledge of the different niches of Malassezia. For example, it is currently known that Malassezia spp. distribution is not limited to the skin because DNA-based assays have demonstrated that they are detected at relatively high frequencies in different parts of the body, such as the oral cavity (38% of sequences) [10], the gastrointestinal tract (88% of samples) [11], the respiratory tract (86% of sequences) [12] and the brain (89% of samples) [13]. The detection of Malassezia DNA in these different body sites, especially in patients with chronic pathologies, raises many questions. Are these yeasts truly present in the investigated body site or did they contaminate this niche from other body sites? The contamination might be due to the entire yeast (living or dead) or only to DNA. Does this DNA come from living or dead yeast? If these yeasts are alive, do they belong to the resident mycobiota or are they transient colonizers which are not permanently established within these niches? Finally, are these yeasts associated with certain chronic diseases or not? In an attempt to shed light on this knowledge gap, we critically reviewed published studies that analyzed the possible association of Malassezia spp. with chronic human diseases.
2. Materials and Methods
We conducted literature searches in PubMed (https://pubmed.ncbi.nlm.nih.gov/) and Web of Science (Clarivate Analytics) on 17 May 2021 using the following query: ((Malassezia and Diseases) OR (Malassezia and Associated) OR (Malassezia and Chronic)). No limits were set on publication date or language. The Zotero ver.5 (Corporation for Digital Scholarship; www.zotero.org, accessed on 17 May 2021) software was used to identify duplicates. We screened the articles by analyzing their titles and/or abstracts.
We only included articles assessing the association or involvement of Malassezia in chronic human diseases.
3. Results
The search results identified 2538 studies. After removing duplicates, we screened 1790 study titles and/or abstracts and excluded 1728 which were unrelated to the present study. Ultimately, 62 full-length articles were assessed for eligibility, and 31 were selected for qualitative analysis and included in this review. This selection flow is presented in Figure 1. The 31 studies detailed below address the association of Malassezia yeast with chronic diseases, including psoriasis, atopic dermatitis (AD), chronic rhinosinusitis (CRS), asthma, cystic fibrosis (CF), HIV infection, inflammatory bowel disease (IBD), colorectal cancer (CRC), and neurodegenerative diseases. We will present the association of Malassezia with psoriasis.
3.1. Association between Malassezia and Psoriasis
Psoriasis is a chronic, inflammatory and multifactorial skin disease with an estimated prevalence of 2% [14]. The association of Malassezia yeast with psoriasis is widely hypothesized, although the pathophysiological mechanisms are not completely known. The studies reporting the association of Malassezia with psoriasis are detailed in Table 1. In some studies, Malassezia appears to be increased in terms of the diversity and prevalence of skin culture in psoriatic patients compared to healthy subjects [15,16,17]. Some authors infer a causality from these findings, despite the fact the studies are not always in agreement with one another. This is the case of one study using a nested polymerase chain reaction (PCR) assay in patients in Japan where the average number of Malassezia species detected in the skin was found to be high in 22 psoriatic patients (3.7 ± 1.6) compared to 30 healthy individuals (2.8 ± 0.8), suggesting a greater diversity of the skin microbiome in psoriatic patients than in healthy subjects [15]. However, there was no correlation between the species richness found and the severity of the disease, although high detection rates were found in psoriatic patients with 96%, 82% and 64% for M. restricta, M. globosa and M. sympodialis, respectively [15]. A high diversity of Malassezia species was also found in 50 psoriatic patients compared to 50 healthy subjects with a positive culture rate of 68% and 60%, respectively [16]. The authors found a higher abundance of colonies isolated from skin lesions (53 colonies) than from normal skin (19 colonies) in psoriatic patients, especially on their scalp (p = 0.03), and no difference was found between patients and healthy subjects. In another Spanish study comparing 40 psoriatic patients and 40 healthy subjects, a difference was found regarding the positivity of direct microscopic examinations of scalp samples between patients (75%) and healthy subjects (30%) [17]. These results were confirmed by culture with a positive culture rate of 85% and 50% in patients and healthy subjects, respectively. In addition, the authors found that patients who had suffered an exacerbation of psoriasis had more Malassezia than non-exacerbated patients [17]. At the species level, M. restricta and M. globosa were most frequently isolated, with 40% and 45% in psoriatic patients compared to 25% and 15% in healthy subjects, respectively [17], suggesting an association between the psoriasis and Malassezia species. Furthermore, in an ITS1 metabarcoding study of 34 patients with chronic plaque psoriasis compared to 25 healthy subjects, M. sympodialis and M. restricta were found with a significantly higher abundance and associated with psoriatic lesions on the elbow and the back, respectively [18].
Despite the data from all these studies regarding the distribution of Malassezia, the mechanisms by which the Malassezia species may initiate or exacerbate psoriasis are not fully elucidated. However, elevated humoral immune responses directed against Malassezia have been observed in patients with psoriasis. It has been reported that antibodies against Malassezia proteins, such as N-acetyl glucosamine, are present in the serum from patients with psoriasis [19,20]. In addition, T cells which are reactive against Malassezia have been isolated from psoriatic skin lesions [21]. It has also been shown that Malassezia interacts with keratinocytes and induces an overproduction of molecules involved in cell migration and hyper-proliferation, such as transforming growth factor-beta, integrin chains and heat shock protein 70 [22]. The overproduction of these molecules is likely to promote the exacerbation of the disease in psoriatic patients. Finally, a highly increased Malassezia-specific Th1 response was observed in psoriatic patients compared to controls [23]. Interestingly, psoriasis-like lesions were observed when a killed suspension of Malassezia was applied to the intact skin of psoriatic patients compared to controls [24], which fulfills one of Koch’s postulates. When taking all these data into account, evidence for the involvement of Malassezia in psoriasis flares, particularly in lesions of the scalp, appears to be convincing. However, further evidence is needed because of the discrepancies between the studies regarding the distribution of Malassezia species on psoriatic patients’ skin, which are probably due to methodological heterogeneity.
3.2. Allergic Diseases Associated with Malassezia
In this section, we will discuss Malassezia spp. in allergic diseases, as these yeast species produce allergens and are also found to be associated with some of these diseases, including chronic rhinosinusitis, atopic dermatitis and asthma.
3.2.1. Chronic Rhinosinusitis
Chronic rhinosinusitis (CRS) is a complex inflammatory disease located in the nose and paranasal sinuses with distinctive symptoms such as nasal obstruction, nasal discharge, facial pain and/or a reduced sense of smell [25]. There are three defined phenotypic groups, including CRS with nasal polyps, CRS without nasal polyps, and allergic fungal rhinosinusitis. Although the interactions between the host immune system and microorganisms have not been fully determined, the pathogenic role of the microbiota in CRS has been suggested due to the presence of intramucosal bacteria, biofilms, microbial dysbiosis and super antigens [26]. DNA-based molecular studies have reported a high abundance of Malassezia in the sinuses of both CRS patients and controls [12,27,28], generating many questions, as these yeast species were previously only known to colonize the skin. In a 18S ribosomal DNA (rDNA) pyrosequencing study of the sinonasal mycobiome, Malassezia was found to be the most abundant fungal genus in 100% of all sinus samples, with a relative abundance of 50.09% and 57.5% in 23 patients with CRS and 11 non-CRS controls, respectively [27]. The presence of Malassezia was of great interest as it was the first time it had been described to be associated with the sinuses. The authors hypothesized that it could represent contamination from the nasal vestibule rather than a permanent colonization. Later, Gelber et al. [28] used quantitative polymerase chain reaction (qPCR) to assess whether Malassezia was a component of the normal sinus microbiome, to identify Malassezia in the sinuses and to compare seven non-CRS controls with CRS subtypes, including 15 CRS patients with polyps, three cases of fungal balls and three cases of allergic fungal rhinosinusitis. In their study, Malassezia was detected in 68% of all samples, with an even prevalence across the four groups (p > 0.99), suggesting a commensal role for this genus of yeast in the nasal cavity [28]. M. restricta was the most commonly detected species with a prevalence of 46% in all subjects, compared to M. globosa (14%, p= 0.029), which was detected only in CRS patients with polyps [28]. Similar results on the predominance of Malassezia were also reported on CRS clinical categories including 50 CRS without nasal polyps, 49 CRS with polyps, seven CRS with cystic fibrosis and 38 control subjects, where Malassezia was detected in 100% of subjects with an abundance of 86% of the sequences [12]. These authors found that the predominant Malassezia species were distinct in patients with cystic fibrosis compared to the other patients. It is not clear whether particular Malassezia species are associated with specific clinical features of CRS. In the case of allergic rhinitis, Malassezia was detected by 28 D1/D2 region pyrosequencing on the nasal vestibule with a relative abundance of 93.1% and 97.5% in four patients and four controls, respectively [29]. M. restricta was predominant and followed by M. globosa, suggesting that these two species are common in the nasal cavity of all clinical categories of CRS. It should be noted that despite the high prevalence of Malassezia in these studies, no significant difference was observed between patients and control subjects, reinforcing the idea of the commensal nature of Malassezia presence in the upper airways. Specific culture media are not routinely used to isolate Malassezia species. It is, therefore, difficult to assess the proportion of viable sinus-associated Malassezia. To further investigate this hypothesis, we conducted a study in our laboratory on 14 sinus biopsies from 11 meatus surgery patients. A Malassezia-specific real-time PCR was performed [30]. In parallel, the culture method was performed on the sinus samples using the FastFung culture medium that we developed in our laboratory [31,32]. In the 11 samples tested, the PCR was positive in 90.9% of samples, while no sample was positive in culture (presented in Table 2). The negative culture of these samples can be explained by the fact that the samples were inoculated two to three days after surgery, as the post-collection survival time of Malassezia in biological samples is not known. Another explanation could be that the DNA detected in the sinuses came from dead yeast.
Little is known about the role and mechanisms in which Malassezia might be involved in the sinonasal pathogenesis of patients with CRS. Recently, Lee et al. [33] showed, in a murine model of acute sinusitis, that bacterial-Malassezia interactions in the sinonasal mucosa influence the host immune response. The authors showed that co-infection of mice with M. sympodialis and S. aureus or p. aeruginosa increased allergic and inflammatory responses by significantly increasing the level of IL-5, IL-13 and IL-17, respectively [33]. These responses depended on the co-infecting bacterial species and were particularly marked in mice co-infected with S. aureus. In addition, the level of expression of the Dectin-1 receptor involved in the fungal sensing was significantly increased when mice were infected with S. aureus or P. aeruginosa in the presence or absence of M. sympodialis compared to mice infected with M. sympodialis only [33]. One of the authors’ hypotheses was that the interaction of these bacteria with Malassezia present in the sinonasal mucosa could alter the behavior of Malassezia by increasing its virulence, thus increasing the expression of Dectin-1 [33]. Since the Dectin-1 receptor recognizes Malassezia allergens [34], its activation could be one of the causes for the increase in IL-5 and IL-13 levels via an increased response to Th2-type CD4+ T cell [33]. These data support the hypothesis that Malassezia species are natural commensals of the upper airways and that they are probably not capable of exacerbating the disease independently. In other words, the contribution that Malassezia makes to the pathogenesis of CRS may be indirect, via interactions with bacteria. Much remains to be known about the impact of Malassezia spp. in the sinonasal mucosa of CRS patients. In particular, further assessment is required of the level of expression of Malassezia virulence genes when co-cultured with bacteria in the context of the disease.
3.2.2. Atopic Dermatitis and Asthma
Atopic dermatitis (AD) is a chronic inflammatory and relapsing skin disease. It is mainly characterized by intense itching and can be associated with allergic rhinitis and allergic asthma. It has been estimated that in the last three decades, the prevalence of AD has tripled in industrialized countries, affecting 30% of children and 10% of adults [35]. The pathogenesis of AD is complex and not yet fully understood. It has been hypothesized that genetic, immunological and environmental factors and impaired skin barrier function contribute to the disease. Many studies have also highlighted the role of the cutaneous microbiota, including Malassezia, in the disease [36]. A few studies have reported high differences in both diversity and abundance of Malassezia species colonization in AD patients compared to controls [15,37]. Most studies did not find significant differences between AD and control patients [38,39,40]. This has led some authors to believe that the implication of Malassezia in AD may be allergic by nature [38]. Indeed, Malassezia displays about 14 known allergens, and each triggers IgE-mediated sensitization in AD patients [5,40]. Many studies, presented in Table 3, which have measured serum-IgE levels, have found significant correlations between the level of Malassezia-specific IgE antibodies and the severity of AD [41,42,43,44]. This positive correlation was especially true for AD patients, with localized lesions on the head and neck and very high IgE levels [42,45]. In addition, AD patients treated with ketoconazole experienced a clinical improvement, with a decreased severity score and number of Malassezia positive cultures, compared to the placebo group [46]. It was therefore concluded that saprophytic yeasts could be a source of allergens in AD. Currently, there are no solid data showing a direct involvement of allergens in the initiation of AD. However, it is accepted that allergens are factors which exacerbate the disease, as there is an established link between AD severity, IgE levels, and proinflammatory cytokine levels, such as Interleukin (IL)-18 [36].
The pathophysiological mechanisms that Malassezia may use to trigger AD are not fully understood. However, two possible mechanisms have been hypothesized to explain Malassezia-host interactions. The first involves a direct interaction via recognition of Malassezia by pattern-recognizing receptors, such as Mincle and Toll-like receptors [47,48]. The second involves an indirect interaction through recognition of the components released by Malassezia. In recent years, many studies have focused on the components released by Malassezia and their role in skin inflammation. They have shown that, in addition to the direct release of allergens, Malassezia produces extracellular vesicles enriched with allergens, small RNAs and various proteins capable of interacting with skin immune cells [49,50,51]. These extracellular vesicles and allergens have been shown to stimulate keratinocytes and dendritic cells to induce high levels of cytokines including tumor necrosis factor-α (TNF-α), IL-6 and IL-10 [50,52]. Increased levels of these cytokines may be involved in skin inflammation in AD patients.
AD is also considered a risk factor for the development of atopic respiratory diseases, such as asthma, particularly in children [53]. Since IgE-mediated sensitization to Malassezia is found to be high in AD patients, special attention should be given to patients with asthma. Studies on the serology of Malassezia IgE in patients with asthma are scarce. However, in one study conducted on 73 AD patients, 156 asthmatic patients and 212 control patients, the prevalence of Malassezia IgE was found to be 53%, 1% and 0.5%, respectively [41]. These results did not support a significant association between Malassezia and asthma. However, in a 18S rRNA pyrosequencing study of sputum from 30 asthma patients and 13 controls, Malassezia was abundantly detected only in asthma patients with a percentage of reads from 0.01% to 21.65% [54]. Another study based on ITS2 metabarcoding recently reported that Malassezia was very abundant in sputum specimens from 40 asthmatic patients, especially children, with a mean abundance of 67.69%, 27.04%, 2.02% and 2.94% in pediatric asthma, adult asthma, healthy adult and healthy pediatric participants, respectively [55]. In addition, asthmatic patients receiving a combination of steroid and leukotriene receptor antagonists therapy have a significantly increased abundance of Malassezia in their airways. These data are striking in that these molecules are lipid-based while the Malassezia species are lipid-dependent. The question of whether the treatment favors the expansion of Malassezia in the airways of patients with asthma remains to be clarified. Further studies are needed, including culture and in vivo and in vitro experiments, to understand the pathophysiology of Malassezia in asthma patients.
3.3. Cystic Fibrosis and Malassezia
Similarly to the gastrointestinal tract, the respiratory tract is a complex polymicrobial niche which includes various bacteria, fungi and viruses [56]. These microorganisms can exacerbate chronic pulmonary diseases including cystic fibrosis (CF). A growing number of DNA-based studies reported high detection frequencies of Malassezia in the respiratory tract of patients with CF. The first molecular study of Malassezia in CF patients resulted from a study comparing culture and DNA-based methods for fungal detection in 77 adult patients with CF [57]. Malassezia was detected in 2/77 patients with CF by sequencing targeting the ITS2 region. An increasing number of subsequent study reported a relatively high prevalence rates of Malassezia. In a study of four French CF patients, Delhaes et al. [58] studied the fungal community of eight sputum samples. They found a high number of Malassezia reads (≈0.015% of fungal reads) in all patients by ITS2 pyrosequencing. At the species level, M. restricta was detected in all patients while M. globosa and M. sympodialis were detected in only one patient. In their study, they also used mycological culture media, but none were appropriate for Malassezia spp. which require specific media. Similar results were found on four CF patients by analyzing the ITS1 region by denaturing high-performance liquid chromatography combined with cloning/sequencing, where M. restricta and M. globosa were the two species identified on the sputum samples from three patients [59]. Another study carried out on six CF patients receiving antibacterial treatment also found an enrichment of a mixture of Candida species and Malassezia in all sputum samples, representing 74–99% fungal reads using a ITS1 pyrosequencing-based analysis [60]. These authors found that the abundance of both fungal genera persisted before and after treatment suggesting stability.
Although the sample sizes and participants in these studies were small, the results converge, suggesting that these common identified fungal species play a potential role in the respiratory tract of patients with CF. For C. albicans, it has already been shown by culture that this yeast species colonizes the airways of between 49.4% and 74.3% of patients in large cohorts of CF patients [61,62]. In addition, this colonization is associated with the exacerbation of the disease and impaired lung function [61], although the pathogenic mechanisms are not yet elucidated. For Malassezia spp., many questions remain unanswered about the Malassezia species detected in respiratory samples. It is unclear whether the DNA detected comes from dead cells or whether Malassezia survives as a commensal in the lungs. In the literature, isolation of Malassezia spp. from the respiratory tract has been reported in immunocompromised patients and was associated with clinical deterioration [63,64], suggesting that the high detection of Malassezia spp. in CF patients is deleterious and deserves further investigation. Recently, an ITS2 metagenomic sequencing study analyzed 33 sputum samples from patients with and without CF pulmonary exacerbation [65]. Interestingly, Malassezia was abundant and associated with CF pulmonary exacerbation. Further studies using in vivo and in vitro models will be necessary to understand the potential pathophysiological mechanisms.
3.4. Malassezia and HIV Infection
Patients living with HIV are subject to several microbial infections secondary to their immunosuppression. These infections can be caused by commensal opportunistic pathogens in different anatomical sites, such as the skin, a localization where Malassezia is the dominant genus of the human skin mycobiota [3]. With incidence of up to 80%, seborrheic dermatitis (SD) due to Malassezia is one of the major diseases affecting HIV patients [66,67]. In some quantitative culture studies, the density of Malassezia was found to be higher in the skin of HIV patients compared to non-HIV controls [68]. One recent study in particular reported that the positive culture rate and density of Malassezia were very high in 30 HIV patients with seborrheic dermatitis (76.7%) compared to 30 HIV patients without seborrheic dermatitis (50%) [69]. This high rate could be explained by the number of CD4 cells present in the patients, as in the seborrheic dermatitis patients, 24 had <500 CD4 cells including nine with <200 CD4 cells. Moreover, it has been reported that the CD4 count is a primary factor associated with the severity of seborrheic dermatitis in HIV patients, as it has been found to be associated with the abundance of yeast in skin lesions, the severity of seborrheic dermatitis and the CD4-positive T lymphocyte count [70]. This shows that CD4 cell-mediated immunity plays a crucial role in controlling the switch of these yeasts from commensal to pathogen. Whether other non-immune factors related to HIV promote the overgrowth of Malassezia remains to be determined. Regarding the difference in the prevalence of Malassezia between HIV and non-HIV patients, a recent study reported a positive culture rate of 69% and 79% in 48 HIV and 48 non-HIV patients, respectively [71]. They found that patients with high CD4 cells had relatively high colonization rates, but the difference with the other groups was not statistically significant. To date, there are no data showing any other consequence of overgrowth of Malassezia spp. in HIV patients outside the skin. However, by studying the gut eukaryotic microbiota using ITS metabarcoding in our laboratory, we found that HIV patients have a higher abundance of Malassezia spp. in their gut compared to healthy subjects [72]. It is not clear whether this high abundance is a common phenomenon in all patients living with HIV or whether it is a coincidence. Remarkably, in the 22/31 HIV patients in whom Malassezia spp. was detected, 15 patients had CD4 counts ranging from 2–436 cells, with an average of 149.73 cells. This suggests that the outcome of HIV-associated immune deficiency on Malassezia spp. is not limited to the skin and that Malassezia spp. probably colonizes other internal organs in immunocompromised HIV patients. Future studies using complementary techniques, including culture, are needed to further investigate the potential impact of Malassezia spp. on HIV patients, particularly within the gut and internal organs.
3.5. Malassezia in Enteric Diseases
Malassezia is increasingly detected at a relatively high prevalence in the gastrointestinal tract both in healthy individuals at a rate of 81–88.3% [11,73] and in diseased individuals at a rate of 68.4–100% [72,74,75]. In this section, we will discuss the potential role of Malassezia in the gastrointestinal tract in several enteric diseases, including inflammatory bowel disease and colorectal cancer.
3.5.1. Inflammatory Bowel Disease
Inflammatory bowel disease, which includes Crohn’s disease and ulcerative colitis, is a chronic inflammatory disease of the gastrointestinal tract. The gut microbiota and environmental and genetic factors influence inflammatory bowel disease although their exact role has not yet been completely determined. Little is known about the role of the gut mycobiota compared to bacterial gut microbiota in these diseases. The role of fungi has begun to generate great interest in research since high levels of anti-Saccharomyces cerevisiae antibodies (ASCA) have been found in the sera of patients with Crohn’s disease [76]. In addition, genome-wide association studies have identified variants in the genes coding for molecules involved in the defense against fungal infections, such as the caspase recruitment domain 9 (CARD9) [77]. By using sequencing methods, alterations of the gut mycobiota in patients with inflammatory bowel disease were observed with a high relative prevalence of yeasts such as Candida, Saccharomyces and Malassezia genera [74,78]. The role of these yeasts in inflammatory bowel disease pathogenesis is not fully understood. However, two studies have shown that C. albicans and S. cerevisiae exacerbate inflammation and colitis in mouse models with chemically induced colitis, suggesting that these members of the gut mycobiota may be associated with the severity of inflammatory bowel disease [79,80].
It has long been thought that Malassezia species were limited to the skin microbiota. For this reason, there was no interest in investigating whether they were involved in other diseases besides the skin, particularly in internal organs. Recently, Limon et al. [81] sought to understand the role of Malassezia in Crohn’s disease after the presence of M. restricta in the gut mucosa had been found to be associated with Crohn’s disease. In their study, they found that the presence of Malassezia was strongly associated with the Crohn’s disease risk allele CARD9S12N, and this association was correlated with an increased abundance of Malassezia in sigmoid colon samples of Crohn’s disease patients. In addition, depending on CARD9 signaling, they found that M. restricta triggers a stronger inflammatory response from innate immune cells (human or mouse dendritic cells) harboring the IBD-associated polymorphism, compared to C. albicans and S. cerevisiae. By giving pathogen-free mice an oral gavage with M. restricta, the authors found that this yeast species causes a shortening of the colon, a worsening of disease activity, and more severe intestinal inflammation characterized by increased mucosal erosion, crypt destruction, and inflammatory cell infiltration. In addition, in pathogen-free mice colonized with eight specific bacteria, M. restricta exacerbated colitis without altering the bacterial microbiota, showing that M. restricta is sufficient to exacerbate the disease by activating the intestinal immune response. One of the most striking results was the correlation of ASCA levels with M. restricta-recognizing IgA and IgG from the sera of patients with Crohn’s disease. The findings of this study showed that in patients with Crohn’s disease, M. restricta may increase disease severity, principally in patients carrying the CARD9S12N allele.
The high detection of Malassezia in the digestive tract has always raised questions, as this yeast is known to belong to the skin mycobiota. The question of whether it is a true living colonizer of the digestive tract remains a subject of debate. Although data from mouse models are difficult to extrapolate to humans, the presence of Malassezia had significant consequences on the outcomes of Crohn’s disease models. Therefore, particular attention should be paid to fungal gut alterations in patients with IBD.
3.5.2. Colorectal Cancer
Colorectal cancer is one of the most frequent chronic diseases affecting the digestive tract, and the gut microbiota can contribute towards this disease [82]. As with the bacterial microbiota, the existing studies on the gut mycobiota show fungal dysbiosis in colorectal cancer patients with an enrichment of certain fungal genera including Malassezia. In a Chinese study conducted by Gao et al. [83], the gut mycobiota was analyzed by sequencing stool samples from 74 colorectal cancer patients, 29 colon-polyp patients and 28 healthy controls. The impacts of anatomic position and tumor stage on the fungal dysbiosis were also investigated. The authors found fungal dysbiosis in colon polyps and colorectal cancer with decreased diversity in polyps patients. An increased proportion of opportunistic fungi, including Trichosporon and Malassezia were observed in stool samples of patients compared to controls. They concluded that these opportunistic fungi and altered biodiversity may favor the progression of CRC. Similar results were found by analyzing the shotgun metagenomic sequences in fecal samples of 73 patients with colorectal cancer and 92 control subjects [84]. The authors identified fungal biomarkers associated with colorectal cancer including Malassezia with a higher prevalence in colorectal cancer patients compared to controls. They then further validated their results in an independent cohort of 90 patients with CRC, 42 patients with adenoma, and 66 control subjects of published repository sequences from Germany and France. This observed association supports the idea that Malassezia may be a biomarker and contribute to colorectal cancer tumorigenesis.
The mechanisms by which Malassezia may contribute to the tumorigenesis of colorectal cancer remain unknown. We found no studies on this topic. However, the contribution of Malassezia in tumorigenesis has been shown by Aykut et al. [85] in mice models with pancreatic ductal adenocarcinoma. In their study, the authors found that the tumor mycobiome was distinct from that of the gut or normal pancreas and that Malassezia spp. were very abundant in pancreatic tumor tissues. Using mouse models, they showed that fungi, including Malassezia spp., can migrate from the gut to the pancreas and can accelerate the progression of ductal pancreatic adenocarcinoma via the complement cascade activation. Interestingly, pancreatic oncogenesis was strongly increased by Malassezia compared to other common fungi, such as Candida, Saccharomyces and Aspergillus. Currently, there are no established hypotheses on the potential mechanisms of Malassezia involvement in cancer, and further studies are needed to investigate the association of Malassezia with CRC.
3.6. Malassezia Neuroinfection in Neurodegenerative Diseases
Neurodegenerative diseases, including Alzheimer’s disease, amyotrophic lateral sclerosis, Parkinson’s disease, and multiple sclerosis, are multifactorial and represent a major human health issue. Although these diseases share some common features, as the deposition of misfolded protein aggregates in distinct central nervous system (CNS) regions, the exact etiological causes are partially known and remain a subject of research. The idea that infectious agents may be among the etiological causes or risk factors for neurodegeneration has been widely discussed and numerous studies have been conducted on viral and bacterial infections [86,87,88]. The neuroinfection by fungi in neurodegenerative diseases has triggered a considerable interest in research since the dawn of next-generation sequencing (NGS) technology. In recent years, a Spanish team has conducted intense research aiming to highlight the possible involvement of fungi in neurodegenerative diseases [13,89,90]. In their studies, they found common fungi that are more abundant in the affected areas of the brain in patients compared to healthy subjects. Among these fungi, Malassezia was widely detected. In the case of Alzheimer’s disease for example, Malassezia was detected by NGS on brain tissue samples in 94.7% of patients, compared to 12.5% of the controls, in two studies involving 19 patients and 16 healthy controls [89,90]. By comparing the results of Alzheimer’s disease patients with younger and elderly controls, they found that the percentage of Malassezia was very high in the Alzheimer’s disease patients (≈4.4%) compared to the younger and elderly controls ( < 0.5%) [90]. In younger and elderly controls, this low percentage of Malassezia suggests that the high burden of Malassezia in the CNS of patients is probably related to the disease and not to age. The authors argued that these results could be used to guide targeted antifungal therapy for Alzheimer’s disease patients. This will serve to answer one of the most important questions, namely, are disseminated fungal infections the cause of Alzheimer’s disease, a risk factor, or simply a consequence of neurodegeneration? [89]. In addition to Alzheimer’s disease, a high Malassezia detection rate was also found in the CNS of patients with amyotrophic lateral sclerosis (100% of 11 patients), Parkinson’s disease (100% of six patients) and multiple sclerosis (90% of 10 patients) patients [13,91,92]. All these studies aimed to demonstrate the presence of fungi in the altered areas of the CNS. However, based on these data, we cannot conclude whether fungi, including Malassezia, have an etiological responsibility or play a role in the exacerbation of neurodegenerative diseases. All that can be currently concluded is that there is a relatively high Malassezia colonization rate in the CNS of patients who suffer from neurodegenerative diseases.
To date, our knowledge on the possible pathogenesis mechanisms involving fungi such as Malassezia in CNS patients with neurodegenerative diseases is limited. It remains to be clarified whether fungal infections of the CNS are the consequence of immune dysfunctions and/or the patients’ genetic background. It was shown that CARD9 plays a critical role in the control of fungal invasion by recruiting neutrophils into the CNS, knowing that CARD9-deficient humans develop fungal infections targeting the CNS [93]. CARD9 is one of the crucial receptors involved in antifungal immunity as shown in other diseases such as Crohn’s disease [81]. In CNS infection, microglial cells actively participate in the production of proinflammatory cytokines. It was shown that microglial IL-1β and CXCL1 production depends on CARD9, and the specific deletion of microglial CARD9 impairs the neutrophil recruitment toward the C. albicans–infected CNS [94]. Given these data, it is plausible that the fungal infections observed in patients with neurodegenerative diseases could be due to deficiencies in the molecules involved in antifungal immunity.
4. Discussion
It is recognized that the skin is the main ecological niche of Malassezia, and its responsibility for some skin infections, such as pityriasis versicolor and seborrheic dermatitis, is no longer the subject of debate. The different anatomical sites, outside the skin, in which Malassezia has been detected, either in the context of chronic diseases or in healthy individuals, are illustrated in Figure 2. However, its causative role in other inflammatory skin diseases including psoriasis and atopic dermatitis remain under discussion. Given the multifactorial cause of these two diseases, the exact role of Malassezia in their physiopathogenesis remains to be clarified. Regarding psoriasis, Malassezia overgrowth, especially within skin lesions (Table 1) suggests that Malassezia may play a role in its exacerbation [16,17]. However, whether Malassezia plays pathogenic role by causing the lesions or whether it is a simple opportunistic colonizer of pre-existing psoriasis lesions remain under discussion. Regarding atopic dermatitis, we found no evidence of any link between Malassezia abundance on the skin and the severity of this disease. In contrast, increased anti-Malassezia IgE antibodies levels have been associated with AD flare-ups (Table 3).
Independently of seborrheic dermatitis in HIV patients, there are conflicting results regarding Malassezia skin colonization, which are probably due to the heterogeneity in the diagnostic procedures used or in the patients’ profiles, including their compliance with antiviral treatments. It is difficult to evaluate the impact of Malassezia skin colonization on HIV infection. However, the abundance of Malassezia within the gastrointestinal tract deserves a special attention, since most patients had relatively low CD4 cell counts [72]. HIV infection is associated with a depletion of CD4+ cells, especially in Th17 within the gastrointestinal tract [71,95]. Because Th17 cells are known to participate in the immunological response against Malassezia in the skin [71], this intestinal immune dysregulation in HIV patients may be one explanation for the overgrowth of Malassezia in the gastrointestinal tract. This hypothesis remains to be confirmed in future studies which should focus on both the clinical and immunological impacts of the presence of Malassezia in the gastrointestinal tract of HIV patients. Regarding Crohn’s disease, Malassezia has been shown to play a role in inflammation using human samples and a mouse model of induced colitis [81]. This role has been explained by the fact that Malassezia produces ligands at the aryl hydrocarbon receptor, which is involved in several functions such as skin homeostasis or the production of proinflammatory cytokines [81]. Moreover, Malassezia was found to be remarkably abundant within the digestive tract of colorectal cancer patients, suggesting that it might be used as a colorectal cancer biomarker [83,84]. Although there are no studies on the link between Malassezia and the pathogenesis of CRC, it has been shown that Malassezia can migrate from the gut to other organs, such as the pancreas, where it is involved in pancreatic tumorigenesis [85].
Additionally, DNA-based analysis of the upper and lower respiratory mycobiota has also shown that Malassezia is highly prevalent in both healthy and diseased subjects. In particular, in the case of CF, the abundance of Malassezia has been associated with CF pulmonary exacerbation [65]. There are no studies on the possible pathophysiological mechanisms of Malassezia on the lungs. Most studies found no difference in the prevalence of Malassezia between CRS patients and healthy subjects in the sinuses, which was consistent with its commensal nature [33].
Regarding neurovegetative diseases, Malassezia was found to be highly abundant in CNS patients [89,90]. It should be borne in mind that all the arguments and evidence provided in these studies have sought to highlight the existence of fungal infections and have not targeted any specific fungal agent. It is, therefore, difficult to identify a significant contribution of each fungal species detected in neuropathology to these different neurodegenerative diseases. Furthermore, these results on fungal colonization in CNS patients remain to be confirmed by other independent teams around the world. If confirmed, one particular consideration would appear to be essential to understand the role and contribution of commonly found fungi such as Malassezia spp. If truly present, is the fungus the inciting force for disease, is it incidental, or does it function in an additive/synergistic manner that leads to disease progression? Are Malassezia the instigator of the disease or an opportunist that colonizes tissues that have been injured by another factor? In the future, it would be interesting, for example, to measure antibodies against Malassezia in the serum of patients, to use a specific probe for fluorescence in situ hybridization (FISH), as well as to use specific anti-Malassezia immunostaining or auto-immunohistochemistry to thoroughly investigate Malassezia spp. in these diseases. Animal models of neurodegenerative diseases infected by Malassezia spp. may provide greater insight into neurological infections. The study of the genetic background of patients may also be important in terms of better understanding these infections.
Overall, our literature review highlighted some important information. First, although the studies tend to transpose the results of the mice models to humans, it is necessary to consider this with hindsight, because human and animal microbiota are different. Furthermore, it is easy to inoculate a known amount of a microorganism into a mouse model and to measure the effects on the host. However, in the clinical setting, it is difficult to assess the number of microorganisms present in a given site at a given time. Second, it should be noted that there are methodological biases from one study to another, which are due to the heterogeneity of either the sampling methods, DNA extraction, DNA-based detection methods, or culture methods used. With respect to DNA-based approaches, some studies have targeted the 18S rRNA gene, while others targeted either the ITS1 or the ITS2 regions and rarely both at the same time. However, there may be an amplification bias of either of two regions on detected fungi [96,97]. In addition, most of these DNA-based assays have either detected or not detected all the DNA that was present in the samples, and they do not distinguish DNA from dead or live cells. This can be problematic because the detection of a microorganism’s DNA does not necessarily link this microorganism with the disease. In the clinical setting, the detected DNA may originate from dead cells or from contamination. The problem of contamination is likely to be excluded because the studies were conducted by different teams around the world. The issue of the viability of Malassezia detected on these different niches must be elucidated. Since Malassezia-specific culture media are not routinely used, especially for internal diseases, the use of molecular viability techniques, such as viability PCR with propidium monoazide, could be used as a solution.
5. Conclusions
The presence of Malassezia yeast, with various levels of abundance, appears to be ubiquitous in the human body and may be associated with several diseases of the internal organs including Crohn’s disease, CRS, diseases of the respiratory tract and, probably, neurovegetative diseases. It has been shown to be involved in psoriasis and atopic dermatitis flare-ups, which is not surprising, as these yeasts are a resident member of the skin mycobiome. Independently of SD, Malassezia colonization appears to be associated with the CD4 count in patients living with HIV. Whether Malassezia is an emerging pathogen or an ancient under-diagnosed pathogen involved in diseases of the internal organs remains to be elucidated, and further studies are needed to fill the knowledge gaps on the pathophysiology of the presence of Malassezia yeast in the different human organs.
Author Contributions
A.A.: conceptualization; data curation; investigation; formal analysis; methodology; visualization; writing of the original draft. S.R.: conceptualization; methodology; investigation; formal analysis; resources; supervision; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Institut Hospitalo-Universitaire (IHU) Méditerranée Infection and by the French Government under the “Investissements d’avenir” (Investments for the Future) programme managed by the Agence Nationale de la Recherche (ANR, fr: National Agency for Research), (reference: Méditerranée Infection 10-IAHU-03) and the Région Provence-Alpes-Côte d’Azur, European ERDF funding (European regional development fund) and PRIMMI ((Plateformes de Recherche et d’Innovation Mutualisées Méditerranée Infection).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hirai, A.; Kano, R.; Makimura, K.; Duarte, E.R. Malassezia nana sp. nov., a Novel Lipid-Dependent Yeast Species Isolated from Animals. Int. J. Syst. Evol. Microbiol. 2004, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Akaza, N.; Akamatsu, H.; Sasaki, Y.; Takeoka, S.; Kishi, M.; Mizutani, H.; Sano, A.; Hirokawa, K.; Nakata, S.; Matsunaga, K. Cutaneous malassezia Microbiota of Healthy Subjects Differ by Sex, Body Part and Season. J. Dermatol. 2010, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; Sequencing, N.C. Human Skin Fungal Diversity. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Zhao, H.; Li, C.; Rajapakse, M.P.; Wong, W.C.; Xu, J.; Saunders, C.W.; Reeder, N.L.; Reilman, R.A.; Scheynius, A.; et al. Genus-Wide Comparative Genomics of Malassezia delineates Its Phylogeny, Physiology, and Niche Adaptation on Human Skin. PLoS Genet. 2015, 11, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Gaitanis, G.; Magiatis, P.; Hantschke, M.; Bassukas, I.D.; Velegraki, A. The Malassezia Genus in Skin and Systemic Diseases. Clin. Microbiol. Rev. 2012, 106–141. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.K.; Batra, R.; Bluhm, R.; Boekhout, T.; Dawson, T.L. Skin Diseases Associated with Malassezia Species. J. Am. Acad. Dermatol. 2004, 51, 785–798. [Google Scholar] [CrossRef]
- Chryssanthou, E.; Broberger, U.; Petrini, B. Malassezia pachydermatis fungaemia in a Neonatal Intensive Care Unit. Acta Paediatr. Int. J. Paediatr. 2001, 90, 323–327. [Google Scholar] [CrossRef]
- Al-Sweih, N.; Ahmad, S.; Joseph, L.; Khan, S.; Khan, Z. Malassezia pachydermatis fungemia in a Preterm Neonate Resistant to Fluconazole and Flucytosine. Med. Mycol. Case Rep. 2014, 5, 9–11. [Google Scholar] [CrossRef]
- Iatta, R.; Cafarchia, C.; Cuna, T.; Montagna, O.; Laforgia, N.; Gentile, O.; Rizzo, A.; Boekhout, T.; Otranto, D.; Montagna, M.T. Bloodstream Infections by Malassezia and Candida Species in Critical Care Patients. Med. Mycol. 2014, 264–269. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, A.K.; David, M.S.; Li, L.; Heider, T.N.; Peterson, J.D.; Montano, E.A.; Dongari-Bagtzoglou, A.; Diaz, P.I.; Strausbaugh, L.D. Redefining the Human Oral Mycobiome with Improved Practices in Amplicon-Based Taxonomy: Discovery of Malassezia as a Prominent Commensal. PLoS ONE 2014, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The Gut Mycobiome of the Human Microbiome Project Healthy Cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Hoggard, M.; Zoing, M.; Biswas, K.; Taylor, M.W.; Douglas, R.G. The Sinonasal Mycobiota in Chronic Rhinosinusitis and Control Patients. Rhinology 2019, 57, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Pisa, D.; Alonso, R.; Carrasco, L. Parkinson’s Disease: A Comprehensive Analysis of Fungi and Bacteria in Brain Tissue. Int. J. Biol. Sci. 2020, 16, 1135–1152. [Google Scholar] [CrossRef] [PubMed]
- Boehncke, W.H.; Schön, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef]
- Amaya, M.; Tajima, M.; Okubo, Y.; Sugita, T.; Nishikawa, A.; Tsuboi, R. Molecular Analysis of Malassezia Microflora in the Lesional Skin of Psoriasis Patients. J. Dermatol. 2007, 34, 619–624. [Google Scholar] [CrossRef]
- Rudramurthy, S.M.; Honnavar, P.; Chakrabarti, A.; Dogra, S.; Singh, P.; Handa, S. Association of Malassezia Species with Psoriatic Lesions. Mycoses 2014, 57, 483–488. [Google Scholar] [CrossRef]
- Gomez-Moyano, E.; Crespo-Erchiga, V.; Martínez-Pilar, L.; Godoy Diaz, D.; Martínez-García, S.; Lova Navarro, M.; Vera Casaño, A. Do Malassezia Species Play a Role in Exacerbation of Scalp Psoriasis? J. Mycol. Med. 2014, 24, 87–92. [Google Scholar] [CrossRef]
- Stehlikova, Z.; Kostovcik, M.; Kostovcikova, K.; Kverka, M.; Juzlova, K.; Rob, F.; Hercogova, J.; Bohac, P.; Pinto, Y.; Uzan, A.; et al. Dysbiosis of Skin Microbiota in Psoriatic Patients: Co-occurrence of Fungal and Bacterial Communities. Front. Microbil. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Squiquera, L.; Galimberti, R.; Morelli, L.; Plotkin, L.; Milicich, R.; Kowalckzuk, A.; Leoni, J. Antibodies to Proteins from Pityrosporum ovale in the Sera from Patients with Psoriasis. Clin. Exp. Dermatol. 1994, 19, 289–293. [Google Scholar] [CrossRef]
- Mathov, I.; Plotkin, L.; Abatangelo, C.; Galimberti, R.; Squiquera, L. Antibodies from Patients with Psoriasis Recognize N -Acetylglucosamine Terminals in Glycoproteins from Pityrosporum ovale. Clin. Exp. Immunol. 1996, 105, 79–83. [Google Scholar] [CrossRef]
- Baker, B.S.; Garjoch, A.P.J.J.; Hardman, C.; Fry, L. Differential T-Cell Reactivity to the Round and Oval Forms of Pityrosporum in the Skin of Patients with Psoriasis. Br. J. Dermatol. 1997, 136, 319–325. [Google Scholar] [CrossRef]
- Baroni, A.; Paoletti, I.; Ruocco, E.; Agozzino, M.; Tufano, M.A.; Donnarumma, G. Possible Role of Malassezia Furfur in Psoriasis: Modulation of TGF-β1, Integrin, and HSP70 Expression in Human Keratinocytes and in the Skin of Psoriasis-Affected Patients. J. Cutan. Pathol. 2004, 31, 35–42. [Google Scholar] [CrossRef]
- Kanda, N.; Tani, K.; Enomoto, U.; Nakai, K.; Watanabe, S. The Skin Fungus-Induced Th1- and Th2-Related Cytokine, Chemokine and Prostaglandin E2 Production in Peripheral Blood Mononuclear Cells from Patients with Atopic Dermatitis and Psoriasis vulgaris. Clin. Exp. Allergy 2002, 2, 1243–1250. [Google Scholar] [CrossRef]
- Lober, C.W.; Belew, P.W.; Rosenberg, E.W.; Bale, G. Patch Tests with Killed Sonicated Microflora in Patients with Psoriasis. Arch. Dermatol. 1982, 118, 322–325. [Google Scholar] [CrossRef]
- Ebbens, F.A.; Georgalas, C.; Fokkens, W.J. Fungus as the Cause of Chronic Rhinosinusitis: The Case Remains Unproven. Curr. Opin. Otolaryngol. Head Neck Surg. 2009, 17, 43–49. [Google Scholar] [CrossRef]
- Sivasubramaniam, R.; Douglas, R. The Microbiome and Chronic Rhinosinusitis. World J. Otorhinolaryngol.-Head Neck Surg. 2018, 4, 216–221. [Google Scholar] [CrossRef]
- Cleland, E.J.; Bassiouni, A.; Bassioni, A.; Boase, S.; Dowd, S.; Vreugde, S.; Wormald, P.-J. The Fungal Microbiome in Chronic Rhinosinusitis: Richness, Diversity, Postoperative Changes and Patient Outcomes. Int. Forum Allergy Rhinol. 2014, 4, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Gelber, J.T.; Cope, E.K.; Goldberg, A.N.; Pletcher, S.D. Evaluation of Malassezia and Common Fungal Pathogens in Subtypes of Chronic Rhinosinusitis. Int. Forum Allergy Rhinol. 2016, 6, 950–955. [Google Scholar] [CrossRef]
- Jung, W.H.; Croll, D.; Cho, J.H.; Kim, Y.R.; Lee, Y.W. Analysis of the Nasal Vestibule Mycobiome in Patients with Allergic Rhinitis. Mycoses 2015, 58, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Paulino, L.C.; Tseng, C.; Strober, B.E.; Blaser, M.J. Molecular Analysis of Fungal Microbiota in Samples from Healthy Human Skin and Psoriatic Lesions. J. Clin. Microbiol. 2006, 44, 2933–2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittar, F.; Gouriet, F.; Khelaifia, S.; Raoult, D.; Ranque, S. FastFung: A Novel Medium for the Culture and Isolation of Fastidious Fungal Species from Clinical Samples. J. Microbiol. Methods 2020, 180, 19–21. [Google Scholar] [CrossRef]
- Abdillah, A.; Khelaifia, S.; Raoult, D.; Bittar, F.; Ranque, S. Comparison of Three Skin Sampling Methods and Two Media for Culturing Malassezia Yeast. J. Fungi 2020, 6, 1–7. [Google Scholar] [CrossRef]
- Lee, K.; Zhang, I.; Kyman, S.; Kask, O.; Kathryn, E. Co-Infection of Malassezia sympodialis with Bacterial Pathobionts Pseudomonas aeruginosa or Staphylococcus aureus Leads to Distinct Sinonasal Inflammatory Responses in a Murine Acute Sinusitis Model. Front. Cell. Infect. Microbiol. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Roesner, L.M.; Ernst, M.; Chen, W.; Begemann, G.; Kienlin, P.; Raulf, M.K.; Lepenies, B.; Werfel, T. Human Thioredoxin, a Damage-Associated Molecular Pattern and Malassezia-Crossreactive Autoallergen, Modulates Immune Responses via the C-Type Lectin Receptors Dectin-1 and Dectin-2. Sci. Rep. 2019, 9, 11210. [Google Scholar] [CrossRef] [Green Version]
- Bieber, T. Atopic Dermatitis. Ann. Dermatol. 2010, 22, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Sonesson, A.; Bartosik, J.; Christiansen, J.; Roscher, I.; Nilsson, F.; Schmidtchen, A.; Bäck, O. Sensitization to Skin-Associated Microorganisms in Adult Patients with Atopic Dermatitis is of Importance for Disease Severity. Acta Derm.-Venereol. 2013, 93, 340–345. [Google Scholar] [CrossRef] [Green Version]
- Kaga, M.; Sugita, T.; Nishikawa, A.; Wada, Y.; Hiruma, M.; Ikeda, S. Molecular Analysis of the Cutaneous Malassezia Microbiota from the Skin of Patients with Atopic Dermatitis of Different Severities. Mycoses 2009, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Yim, S.M.; Kim, J.Y.; Ko, J.H.; Lee, Y.W.; Choe, Y.B.; Ahn, K.J. Molecular Analysis of Malassezia Microflora on the Skin of the Patients with Atopic Dermatitis. Ann. Dermatol. 2010, 22, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, E.; Tanaka, T.; Tajima, M.; Tsuboi, R.; Nishikawa, A.; Sugita, T. Characterization of the Skin Fungal Microbiota in Patients with Atopic Dermatitis and in Healthy Subjects. Microbiol. Immunol. 2011, 55, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Glatz, M.; Bosshard, P.; Schmid-Grendelmeier, P. The Role of Fungi in Atopic Dermatitis. Immunol. Allergy Clin. NA 2017, 37, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Scalabrin, D.M.F.; Bavbek, S.; Perzanowski, M.S.; Wilson, B.B.; Platts-Mills, T.A.E.; Wheatley, L.M. Use of Specific IgE in Assessing the Relevance of Fungal and Dust Mite Allergens to Atopic Dermatitis: A Comparison with Asthmatic and Nonasthmatic Control Subjects. J. Allergy Clin. Immunol. 1999, 104, 1273–1279. [Google Scholar] [CrossRef]
- Bayrou, O.; Pecquet, C.; Flahault, A.; Artigou, C.; Abuaf, N.; Leynadier, F. Head and Neck Atopic Dermatitis and Malassezia-Furfur-Specific IgE Antibodies. Dermatology 2005, 211, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Hiragun, M.; Hiragun, T.; Ishii, K.; Suzuki, H.; Tanaka, A.; Yanase, Y.; Mihara, S.; Haruta, Y.; Kohno, N.; Hide, M. Elevated Serum IgE against MGL_1304 in Patients with Atopic Dermatitis and Cholinergic Urticaria. Allergol. Int. 2014, 63, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittermann, I.; Wikberg, G.; Johansson, C.; Lupinek, C.; Lundeberg, L.; Crameri, R.; Valenta, R.; Scheynius, A. IgE Sensitization Profiles Differ between Adult Patients with Severe and Moderate Atopic Dermatitis. PLoS ONE 2016, 11, e0156077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodská, P.; Panzner, P.; Pizinger, K.; Schmid-Grendelmeier, P. IgE-Mediated Sensitization to Malassezia in Atopic Dermatitis: More Common in Male Patients and in Head and Neck Type. Dermatitis 2014, 25, 120–126. [Google Scholar] [CrossRef]
- Lintu, P.; Savolainen, J.; Kortekangas-Savolainen, O.; Kalimo, K. Systemic Ketoconazole is an Effective Treatment of Atopic Dermatitis with IgE-Mediated Hypersensitivity to Yeasts. Allergy Eur. J. Allergy Clin. Immunol. 2001, 56, 512–517. [Google Scholar] [CrossRef]
- Yamasaki, S.; Matsumoto, M.; Takeuchi, O.; Matsuzawa, T.; Ishikawa, E.; Sakuma, M.; Tateno, H.; Uno, J.; Hirabayashi, J.; Mikami, Y.; et al. C-Type Lectin Mincle is an Activating Receptor for Pathogenic Fungus, Malassezia. Proc. Natl. Acad. Sci. USA 2009, 106, 1897–1902. [Google Scholar] [CrossRef] [Green Version]
- Sparber, F.; LeibundGut-Landmann, S. Host Responses to Malassezia spp. in the Mammalian Skin. Front. Immunol. 2017, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Johansson, H.J.; Vallhov, H.; Holm, T.; Gehrmann, U.; Andersson, A.; Johansson, C.; Blom, H.; Carroni, M.; Lehtiö, J.; Scheynius, A. Extracellular Nanovesicles Released from the Commensal Yeast Malassezia sympodialis are Enriched in Allergens and Interact with Cells in Human Skin. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Han, Y.; Sun, Y.; Jiang, H.; Liu, M.; Qi, R.; Gao, X. Extracellular Vesicles Derived from Malassezia Furfur Stimulate IL-6 Production in Keratinocytes as Demonstrated in In Vitro and In Vivo Models. J. Dermatol. Sci. 2019, 93, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Vallhov, H.; Johansson, C.; Veerman, R.E.; Scheynius, A. Extracellular Vesicles Released from the Skin Commensal Yeast Malassezia sympodialis Activate Human Primary Keratinocytes. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, S.; Zou, Z.; Tao, A.; Ai, Y. Activation Profile of THP-1 Derived Dendritic Cells Stimulated by Allergen Mal f 1 beyond its IgE-Binding Ability. Int. Immunopharmacol. 2018, 62, 139–146. [Google Scholar] [CrossRef]
- Ong, P.Y.; Ferdman, R.M.; Church, J.A. Association of Microbial IgE Sensitizations with Asthma in Young Children with Atopic Dermatitis. Ann. Allergy Asthma Immunol. 2012, 108, 212–213. [Google Scholar] [CrossRef]
- Van Woerden, H.C.; Gregory, C.; Brown, R.; Marchesi, J.R.; Hoogendoorn, B.; Matthews, I.P. Differences in Fungi Present in Induced Sputum Samples from Asthma Patients and Non-Atopic Controls: A Community Based Case Control Study. BMC Infect. Dis. 2013, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Al Bataineh, M.T.; Hamoudi, R.A.; Dash, N.R.; Ramakrishnan, R.K.; Almasalmeh, M.A.; Sharif, H.A.; Al-Hajjaj, M.S.; Hamid, Q. Altered Respiratory Microbiota Composition and Functionality Associated with Asthma Early in Life. BMC Infect. Dis. 2020, 20, 697. [Google Scholar] [CrossRef]
- Man, W.H.; Piters, W.A.A.D.S.; Bogaert, D. The Microbiota of the Respiratory Tract: Gatekeeper to Respiratory Health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Nagano, Y.; Elborn, S.; Millar, C.; Walker, J.M.; Goldsmith, C.E.; Rendall, J.; Moore, J.E. Comparison of Techniques to Examine the Diversity of Fungi in Adult Patients with Cystic Fibrosis. Med. Mycol. 2010, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Delhaes, L.; Monchy, S.; Fréalle, E.; Hubans, C.; Salleron, J.; Leroy, S.; Prevotat, A.; Wallet, F.; Wallaert, B.; Dei-Cas, E.; et al. The Airway Microbiota in Cystic Fibrosis: A Complex Fungal and Bacterial Community-Implications for Therapeutic Management. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Mounier, J.; Gouëllo, A.; Keravec, M.; Le Gal, S.; Pacini, G.; Debaets, S.; Nevez, G.; Rault, G.; Barbier, G.; Héry-Arnaud, G. Use of Denaturing High-Performance Liquid Chromatography (DHPLC) to Characterize the Bacterial and Fungal Airway Microbiota of Cystic Fibrosis Patients. J. Microbiol. 2014, 52, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Willger, S.D.; Grim, S.L.; Dolben, E.L.; Shipunova, A.; Hampton, T.H.; Morrison, H.G.; Filkins, L.M.; Toole, G.A.O.; Moulton, L.A.; Ashare, A.; et al. Characterization and Quantification of the Fungal Microbiome in Serial Samples from Individuals with Cystic Fibrosis. Microbiome 2014, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chotirmall, S.H.; O’Donoghue, E.; Bennett, K.; Gunaratnam, C.; O’Neill, S.J.; McElvaney, N.G. Sputum Candida Albicans Presages FEV 1 Decline and Hospital-Treated Exacerbations in Cystic Fibrosis. Chest 2010, 138, 1186–1195. [Google Scholar] [CrossRef]
- Masoud-Landgraf, L.; Badura, A.; Eber, E.; Feierl, G.; Marth, E.; Buzina, W. Modified Culture Method Detects a High Diversity of Fungal Species in Cystic Fibrosis Patients. Med. Mycol. 2014, 52, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemmer, A.W.; Yilmaz, E.; Mittal, R.; Müller-Edenborn, B.; Haas, A.; Holzinger, A.; Schulze, A. Malassezia Furfur Colonising the Respiratory Tract of Mechanically Ventilated Neonates. Z. Geburtshilfe Neonatol. 2008, 212, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Blaes, A.H.; Cavert, W.P.; Morrison, V.A. Malassezia: Is it a Pulmonary Pathogen in the Stem Cell Transplant Population? Transpl. Infect. Dis. 2009, 11, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Soret, P.; Vandenborght, L.E.; Francis, F.; Coron, N.; Enaud, R.; Avalos, M.; Schaeverbeke, T.; Berger, P.; Fayon, M.; Thiebaut, R.; et al. Respiratory Mycobiome and Suggestion of Inter-Kingdom Network during Acute Pulmonary Exacerbation in Cystic Fibrosis. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tragiannidis, A.; Bisping, G.; Koehler, G.; Groll, A.H. Minireview: Malassezia Infections in Immunocompromised Patients. Mycoses 2010, 53, 187–195. [Google Scholar] [CrossRef]
- Forrestel, A.K.; Kovarik, C.L.; Mosam, A.; Gupta, D.; Maurer, T.A.; Micheletti, R.G. Diffuse HIV-Associated Seborrheic Dermatitis-A Case Series. Int. J. STD AIDS 2016, 27, 1342–1345. [Google Scholar] [CrossRef] [PubMed]
- Pechère, M.; Remondat, C.; Bertrand, C.; Didierjean, L.; Saurat, J.H. A Simple Quantitative Culture of Malassezia spp. In HIV-Positive Persons. Dermatology 1995, 191, 348–349. [Google Scholar] [CrossRef]
- Moreno-Coutiño, G.; Sánchez-Cárdenas, C.D.; Bello-Hernández, Y.; Fernández-Martínez, R.; Arroyo-Escalante, S.; Arenas, R. Isolation of Malassezia Spp. in HIV-Positive Patients with and without Seborrheic Dermatitis. An. Bras. De Dermatol. 2019, 94. [Google Scholar] [CrossRef]
- Schechtman, R.; Hay, R.J.; Midgley, G. HIV Disease and Malassezia Yeasts: A Quantitative Study of Patients Presenting with Seborrhoeic Dermatitis. Br. J. Dermatol. 1995, 133, 694–698. [Google Scholar] [CrossRef]
- Krzyściak, P.; Bakuła, Z.; Gniadek, A.; Garlicki, A.; Tarnowski, M.; Wichowski, M.; Jagielski, T. Prevalence of Malassezia Species on the Skin of HIV-Seropositive Patients. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hamad, I.; Abdallah, R.A.; Ravaux, I.; Mokhtari, S.; Tissot-Dupont, H.; Michelle, C.; Stein, A.; Lagier, J.C.; Raoult, D.; Bittar, F. Metabarcoding Analysis of Eukaryotic Microbiota in the Gut of HIV-Infected Patients. PLoS ONE 2018, 13, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suhr, M.J.; Banjara, N.; Hallen-Adams, H.E. Sequence-Based Methods for Detecting and Evaluating the Human Gut Mycobiome. Lett. Appl. Microbiol. 2015, 62, 209–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liguori, G.; Lamas, B.; Richard, M.L.; Brandi, G.; da Costa, G.; Hoffmann, T.W.; Di Simone, M.P.; Calabrese, C.; Poggioli, G.; Langella, P.; et al. Fungal Dysbiosis in Mucosa-Associated Microbiota of Crohn’s Disease Patients. J. Crohn’s Colitis 2016, 10, 296–305. [Google Scholar] [CrossRef]
- Frykman, P.K.; Nordenskjöld, A.; Kawaguchi, A.; Hui, T.T.; Granström, L.; Cheng, Z.; Tang, J.; Underhill, D.M.; Iliev, I.; Funari, V.A. Characterization of Bacterial and Fungal Microbiome in Children with Hirschsprung Disease with and without a History of Enterocolitis: A Multicenter Study. PLoS ONE 2015, 10, e0124172. [Google Scholar] [CrossRef] [Green Version]
- Quinton, J.F.; Sendid, B.; Reumaux, D.; Duthilleul, P.; Cortot, A.; Grandbastien, B.; Charrier, G.; Targan, S.R.; Colombel, J.F.; Poulain, D. Anti-Saccharomyces cerevisiae Manna Antibodies Combined with Antineutrophil Cytoplasmic Autoantibodies in Inflammatory Bowel Disease: Prevalence and Diagnostic Role. Gut 1998, 42, 788–791. [Google Scholar] [CrossRef] [Green Version]
- Rivas, M.A.; Beaudoin, M.; Gardet, A.; Stevens, C.; Zhang, C.K.; Boucher, G.; Ripke, S.; Ellinghaus, D.; Burtt, N.; Fennell, T.; et al. Deep Resequencing of GWAS Loci Identifies Independent Rare Variants Associated with Inflammatory Bowel Disease. Nat. Genet. 2011, 43, 1066–1073. [Google Scholar] [CrossRef] [Green Version]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.-P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal Microbiota Dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Jawhara, S.; Thuru, X.; Standaert-Vitse, A.; Jouault, T.; Mordon, S.; Sendid, B.; Desreumaux, P.; Poulain, D. Colonization of Mice by Candida albicans Is Promoted by Chemically Induced Colitis and Augments Inflammatory Responses through Galectin-3. J. Infect. Dis. 2008, 197, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Chiaro, T.R.; Soto, R.; Stephens, W.Z.; Kubinak, J.L.; Petersen, C.; Gogokhia, L.; Bell, R.; Delgado, J.C.; Cox, J.; Voth, W.; et al. A Member of the Gut Mycobiota Modulates Host Purine Metabolism Exacerbating Colitis in Mice. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Limon, J.J.; Tang, J.; Li, D.; Targan, S.R.; Mcgovern, D.P.B.; Underhill, D.M.; Limon, J.J.; Tang, J.; Li, D.; Wolf, A.J.; et al. Malassezia Is Associated with Crohn’s Disease and Exacerbates Colitis in Mouse Models. Cell Host Microbe 2019, 25, 377–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.A.; Pezet, D.; Bonnet, M. Gut Microbiota Imbalance and Colorectal Cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Kong, C.; Li, H.; Huang, L.; Qu, X.; Qin, N.; Qin, H. Dysbiosis Signature of Mycobiota in Colon Polyp and Colorectal Cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2457–2468. [Google Scholar] [CrossRef]
- Coker, O.O.; Nakatsu, G.; Dai, R.Z.; Wu, W.K.K.; Wong, S.H.; Ng, S.C.; Chan, F.K.L.J.J.Y.S.; Yu, J. Enteric Fungal Microbiota Dysbiosis and Ecological Alterations in Colorectal Cancer. Gut 2018, 68. [Google Scholar] [CrossRef] [PubMed]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The Fungal Mycobiome Promotes Pancreatic Oncogenesis via Activation of MBL. Nature 2019, 574, 264–267. [Google Scholar] [CrossRef]
- Itzhaki, R.F.; Wozniak, M.A.; Appelt, D.M.; Balin, B.J. Infiltration of the Brain by Pathogens Causes Alzheimer’s Disease. Neurobiol. Aging 2004, 25, 619–627. [Google Scholar] [CrossRef]
- Miklossy, J.; Kis, A.; Radenovic, A.; Miller, L.; Forro, L.; Martins, R.; Reiss, K.; Darbinian, N.; Darekar, P.; Mihaly, L.; et al. Beta-amyloid Deposition and Alzheimer’s Type Changes Induced by Borrelia spirochetes. Neurobiol. Aging 2006, 27, 228–236. [Google Scholar] [CrossRef] [Green Version]
- De Chiara, G.; Marcocci, M.E.; Sgarbanti, R.; Civitelli, L.; Ripoli, C.; Piacentini, R.; Garaci, E.; Grassi, C.; Palamara, A.T. Infectious Agents and Neurodegeneration. Mol. Neurobiol. 2012, 614–638. [Google Scholar] [CrossRef] [Green Version]
- Alonso, R.; Pisa, D.; Aguado, B.; Carrasco, L. Identification of Fungal Species in Brain Tissue from Alzheimer ’s Disease by Next-Generation Sequencing. J. Alzheimer. Dis. 2017, 58, 55–67. [Google Scholar] [CrossRef]
- Alonso, R.; Pisa, D.; Fernández-Fernández, A.M.; Carrasco, L. Infection of Fungi and Bacteria in Brain Tissue from Elderly Persons and Patients with Alzheimer’s Disease. Front. Aging Neurosci. 2018, 10, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Alonso, R.; Pisa, D.; Fernández-Fernández, A.M.; Rábano, A.; Carrasco, L. Fungal Infection in Neural Tissue of Patients with Amyotrophic Lateral Sclerosis. Neurobiol. Dis. 2017, 108, 249–260. [Google Scholar] [CrossRef]
- Alonso, R.; Fernández-Fernández, A.M.; Pisa, D.; Carrasco, L. Multiple Sclerosis and Mixed Microbial Infections. Direct Identification of Fungi and Bacteria in Nervous Tissue. Neurobiol. Dis. 2018, 117, 42–61. [Google Scholar] [CrossRef]
- Drummond, R.A.; Collar, A.L.; Swamydas, M.; Rodriguez, A.; Lim, J.K.; Mendez, L.M.; Fink, D.L.; Hsu, A.P.; Zhai, B.; Karauzum, H.; et al. CARD9-Dependent Neutrophil Recruitment Protects against Fungal Invasion of the Central Nervous System. PLoS Pathog. 2015, 11, e1005293. [Google Scholar] [CrossRef]
- Drummond, R.A.; Swamydas, M.; Oikonomou, V.; Zhai, B.; Dambuza, I.M.; Schaefer, B.C.; Bohrer, A.C.; Mayer-Barber, K.D.; Lira, S.A.; Iwakura, Y.; et al. CARD9+ Microglia Promote Antifungal Immunity via IL-1β- and CXCL1-Mediated Neutrophil Recruitment. Nat. Immunol. 2019, 20, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Bixler, S.L.; Mattapallil, J.J. Loss and Dysregulation of Th17 Cells during HIV Infection. Clin. Dev. Immunol. 2013, 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamad, I.; Ranque, S.; Azhar, E.I.; Yasir, M. Culturomics and Amplicon-based Metagenomic Approaches for the Study of Fungal Population in Human Gut Microbiota. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Mbareche, H.; Veillette, M.; Bilodeau, G.; Duchaine, C. Comparison of the Performance of ITS1 and ITS2 as Barcodes in Amplicon-Based Sequencing of Bioaerosols. PeerJ 2020, 2020, 1–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Selection flow-chart of the studies included in the review.

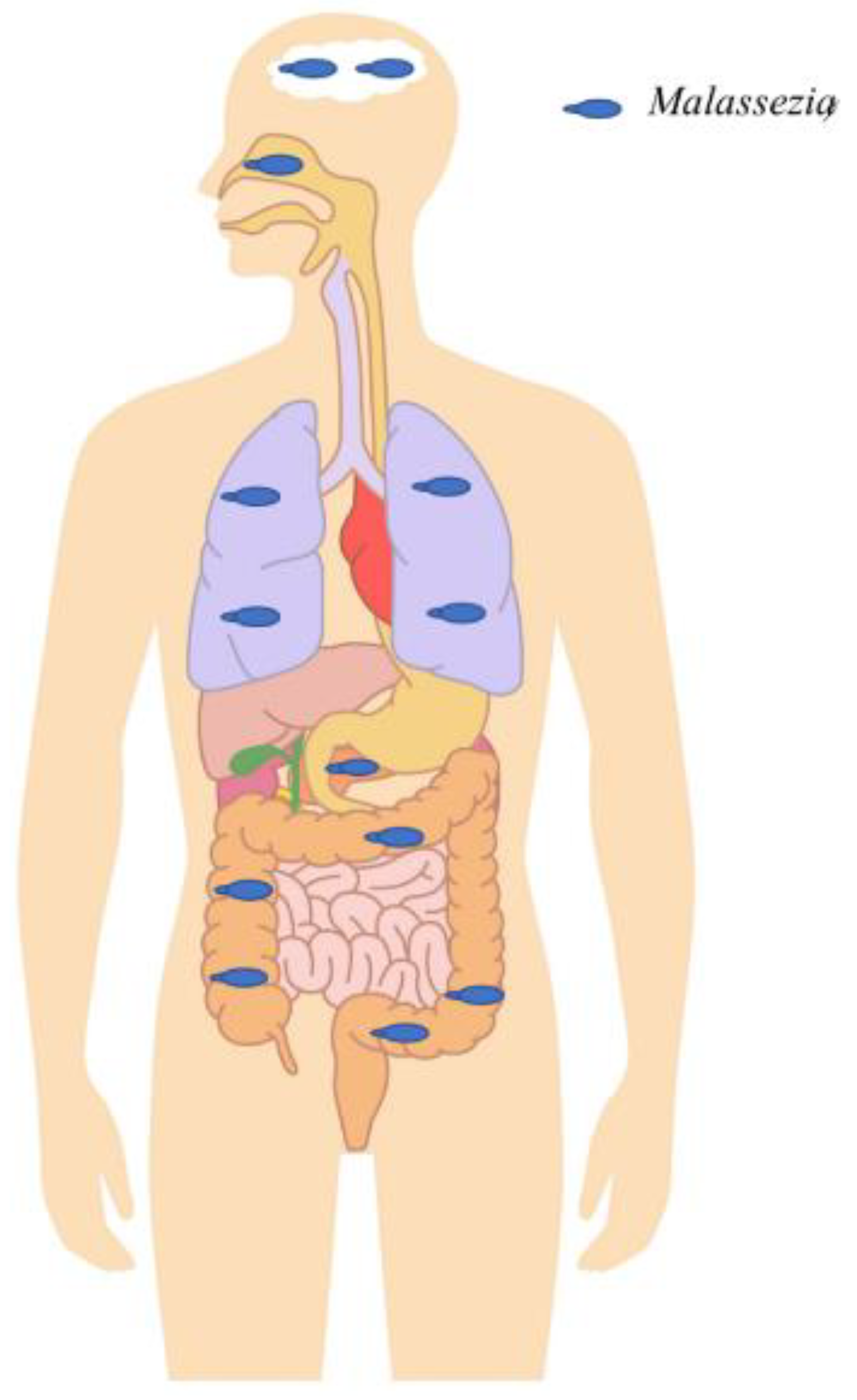
Figure 2.
Malassezia in different anatomical sites detected in the context of chronic diseases or healthy individuals outside the skin. Malassezia was abundantly detected in the digestive tract and was shown to be associated with Crohn’s disease by exacerbating colitis and inflammation. Furthermore, it was highly abundant in colorectal cancer patients compared to healthy subjects, suggesting a potential role in tumorigenesis. It was also shown to be able to migrate from the gastrointestinal tract lumen to the pancreas and to exacerbate pancreatic adenocarcinoma progression. High abundance was also found in the gastrointestinal tracts of HIV patients, probably due to HIV-related immunosuppression. It was highly detected in the upper and lower respiratory tract. Within the upper respiratory tract, it has been shown to interact with specific bacteria (Pseudomonas aeruginosa and Staphylococcus aureus) and to increase chronic rhinosinusitis-associated inflammation. Within the lower respiratory tract, it was shown to be associated with cystic fibrosis pulmonary exacerbations. Finally, Malassezia were detected with a relatively high prevalence and abundance in brain tissue samples of patients with neurodegenerative diseases, such as Alzheimer’s disease, Parkinson’s disease, etc.
Figure 2.
Malassezia in different anatomical sites detected in the context of chronic diseases or healthy individuals outside the skin. Malassezia was abundantly detected in the digestive tract and was shown to be associated with Crohn’s disease by exacerbating colitis and inflammation. Furthermore, it was highly abundant in colorectal cancer patients compared to healthy subjects, suggesting a potential role in tumorigenesis. It was also shown to be able to migrate from the gastrointestinal tract lumen to the pancreas and to exacerbate pancreatic adenocarcinoma progression. High abundance was also found in the gastrointestinal tracts of HIV patients, probably due to HIV-related immunosuppression. It was highly detected in the upper and lower respiratory tract. Within the upper respiratory tract, it has been shown to interact with specific bacteria (Pseudomonas aeruginosa and Staphylococcus aureus) and to increase chronic rhinosinusitis-associated inflammation. Within the lower respiratory tract, it was shown to be associated with cystic fibrosis pulmonary exacerbations. Finally, Malassezia were detected with a relatively high prevalence and abundance in brain tissue samples of patients with neurodegenerative diseases, such as Alzheimer’s disease, Parkinson’s disease, etc.


{kind=link}
{kind=link}
Table 1.
Studies showing the association of Malassezia with psoriasis.
| Psoriasis Patients (PP) and/or Controls | Samples | Methods | Malassezia | References |
|---|---|---|---|---|
| 22 PP vs. 30 healthy subjects (HS) | lesional and non-lesional skin | Nested PCR | High detection (64–96%) in PP Species diversity (3.7 ± 1.6 in PP vs. 2.8 ± 0.8 in HS) | [15] |
| 50 PP vs. 50 HS | lesional and non-lesional skin | Culture/ITS2 PCR-RFLP and D1/D2 sequencing | 68% of PP cases vs. 60% of HS In PP, more colonies isolated in psoriatic lesions (53 colonies) than in non-lesional skin (19), especially on the scalp (p = 0.03). High species richness in PP than HS | [16] |
| 34 PP vs. 25 HS | skin lesion and non-lesion | ITS1 metabarcoding | A significantly higher abundance of M. restricta and M. sympodialis in psoriatic lesions than HS | [18] |
Table 2.
Culture and PCR results on sinus biopsies.
| Patients | Sex | Age (Years) | Samples | Culture | Malassezia PCR |
|---|---|---|---|---|---|
| Patient 1 | Female | 77 | Sinus | Negative | Positive |
| Patient 2 | Female | 27 | Sinus | Negative | Positive |
| Patient 3 | Male | 52 | Sinus | Negative | Positive |
| Patient 4 | Male | 56 | Sinus | Negative | Positive |
| Patient 5 | Female | 44 | Sinus | Negative | Positive |
| Patient 6 | Female | 53 | Sinus | Negative | Positive |
| Patient 7 | Female | 13 | Sinus 1 | Negative | Positive |
| Sinus 2 | Negative | Positive | |||
| Patient 8 | Male | 48 | Sinus | Negative | Negative |
| Patient 9 | Male | 25 | Sinus 1 | Negative | Positive |
| Sinus 2 | Negative | Positive | |||
| Patient 10 | Female | 53 | Sinus | Negative | NA |
| Patient 11 | Female | 72 | Sinus 1 | Negative | NA |
| Sinus 2 | Negative | NA |
NA: not applicable; DNA not available.
Table 3.
Studies associating Malassezia with allergic diseases.
| Patients and/or Controls | Samples | Methods | Malassezia | References |
|---|---|---|---|---|
| 23 CRS vs. 11 controls | Sinonasal swabs | 18S pyrosequencing | 100% of all sinus samples Relative abundance of 50.09% in CRS patients vs. 57.5% in 11 controls | [27] |
| 21 CRS vs. seven controls | Sinus brushings | qPCR | 68% of all samples with no prevalence variation among the groups (p > 0.99) M. restricta (46%) than M. globosa (14%, p = 0.029) | [28] |
| 106 CRS vs. 38 controls | Mucosal swabs | ITS2 metabarcoding | 100% of subjects with an abundance of 86% of the sequences | [12] |
| 56 AD vs. 32 controls | lesional and non-lesional skin | qPCR | High Malassezia colonization in patients with severe AD, ≈ two- to fivefold that in mild and moderate AD patients and healthy subjects (p < 0.05) | [37] |
| 106 head and neck AD (HNAD) vs. 61 controls | Blood | Anti-Malassezia IgE assays (Pharmacia CAP System) | Significant correlation between M. furfur IgE levels and severity in HNAD patients (p < 0.0001) | [42] |
| 63 AD vs. 23 controls | Blood | Anti-M. globosa IgE by Enzyme-linked immunosorbent assay (ELISA) | High significantly IgE and correlated with severity in AD patients compared to normal controls (p < 0.001) | [43] |
| 53 severe AD and 126 AD moderate vs. 140 controls | Blood | Anti-M. sympodialis IgE (ImmunoCAP™) | 62% of severe AD compared to 39% of moderate AD (p < 0.01), No positive controls | [44] |
| 74 HNAD vs. 99 Non-HNAD | Blood | Anti-Malassezia IgE (ImmunoCAP™) | Significantly higher levels of IgE in HNAD patients than non-HAND (p < 0.001) | [45] |
| 73 AD, 156 asthmatic and 212 control patients | Blood | Anti-M. furfur IgE Fluoroimmuno assay (CAP system) | AD (53%), asthmatic (1%) and non-asthmatic control subjects (0.5%) | [41] |
| 30 asthma patients vs. 13 controls | Sputum | 18S pyrosequencing | Only in asthma patients with a percentage of reads from 0.012% to 21.651% | [54] |
| 21 asthma patients vs. 19 controls | Sputum | ITS2 metabarcoding | Mean abundance of 67.69%, 27.04%, 2.02% and 2.94% in pediatric asthma, adult asthma, healthy adult, and healthy pediatric participants, respectively. Significant abundance of Malassezia in the airways of asthmatic patients receiving steroid therapy combined with leukotriene receptor antagonists. | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Abdillah, A.; Ranque, S. Chronic Diseases Associated with Malassezia Yeast. J. Fungi 2021, 7, 855. https://doi.org/10.3390/jof7100855
AMA Style
Abdillah A, Ranque S. Chronic Diseases Associated with Malassezia Yeast. Journal of Fungi. 2021; 7(10):855. https://doi.org/10.3390/jof7100855
Chicago/Turabian StyleAbdillah, Abdourahim, and Stéphane Ranque. 2021. "Chronic Diseases Associated with Malassezia Yeast" Journal of Fungi 7, no. 10: 855. https://doi.org/10.3390/jof7100855
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.