1. Introduction
Arbuscular mycorrhizal (AM) fungi are an important component of the plant soil microbiome. AM fungi are a low diversity monophyletic group of fungi living in close association with many terrestrial plant species [
1]. The intimate association between plant and fungi occurs at the root level, and the main benefit consists of the exchange of inorganic nutrients from soil, provided by the fungus, with carbon fixed during the plant photosynthesis [
2]. In addition, the symbiosis provides further advantages for the plants, allowing them to better tolerate biotic and abiotic stresses [
3], thus increasing fruit yield and quality (e.g., [
4,
5,
6]).
Due to the low diversity of AM fungi (ca. 300 described species [
1]) able to colonize 80% of terrestrial plant species, the symbiosis has been considered as low specific. However, findings point towards non-random association patterns in plant-AM fungal identities [
7], and the wide functional diversity exhibited by fungal taxa [
8] highlights the importance of the knowledge of plant-fungal partner identity when optimizing agricultural production systems. Furthermore, it has been shown that different varieties of the same crop can respond differently to the same AM fungal isolates [
9,
10]. Other authors have found differences in the composition of AM fungal communities associated with different varieties, even when initially exposed to the same AM fungal community. For example, Taylor et al. [
11] found that different onion genotypes displayed subtly different preferences for AM fungi from a mixed community, suggesting a selection process controlled by the plant and/or fungi (see also [
12,
13]).
Vitis vinifera is one of the most important crops in modern agriculture. Its economic and cultural impact is undeniable [
14,
15], especially in the Mediterranean region which harbors 40% of the world total vineyard area [
16]. The availability of water for Mediterranean agriculture is a persistent and growing problem due to climate change, representing a limitation and a threat to the future of our primary sector [
17]. As shown when associated with other plants, AM fungi provide multiple functions to grapes and therefore to vineyards that can help overcome current restrictions to their cultivation: They increase grape uptake of N [
18,
19], reduce soil Cu toxicity [
20], improve plant drought resistance [
21], and protect against pests [
22] (but see [
23]). Moreover, it has been suggested that the whole microbiome and, particularly, AM fungi can alter the biochemical composition of grapevines [
24,
25], which is a key factor in the commercial market of grapevine subproducts. Despite this importance, there are few studies addressing the composition of associated AM fungal communities with vineyards. The scarce evidence suggests that AM fungal assemblages in vineyards are strongly influenced by soil type [
26,
27] and, at a lesser extent, by host plant [
28] and management practices [
29,
30,
31] presenting low phenological variation [
28,
32]. The cultivation of vineyards is done through the grafting of different cultivars onto target rootstock resistant to
Phylloxera; therefore, several cultivars are supported by genetically identical rootstocks. However, none has focused on potential differences of grafted cultivars even when it has been demonstrated that rootstocks differ in their response to AM fungi [
21].
Here, we focus on analyzing the AM fungal community associated with two cultivars grafted onto the same rootstock, a local cultivar name Callet and a widely-distributed one, Merlot. The use of cultivars locally adapted can be one of the major adaptation practices to face climate change as they have been described as physiologically better adapted to deficient irrigation, particularly with regard to water use efficiency [
33,
34]. In fact, a recent work pointed out that under similar water-deficit stress, Callet (and other local cultivars) displayed a better physiological performance compared to the widespread ones [
35]. In this frame, we particularly aim to resolve the following questions:
- (i)
Quantify the diversity of AM fungi associated with the vineyard, both those directly colonizing roots and those present at the agroecosystem level (surrounding soil);
- (ii)
Investigate the AM fungal community composition associated with each vineyard cultivar;
- (iii)
Quantify the relative importance of environmental/spatial factors in the distribution of AM fungal communities.
The obtained information will help to determine which characteristics of the crop may be decisive when selecting the most related AM fungal species, and also will help inform who directs the establishment of the symbiosis: Plant or fungus. If fungi drive the symbiotic relationship, the AM fungal community composition will be primarily determined by environmental/spatial factors and not by the vine cultivar. On the contrary, if the vine cultivars select their community composition, the symbiosis would be more likely driven by the plant.
2. Materials and Methods
The experiment was carried out in the commercial vineyard Can Axartell in Pollença (UTM: 31S 501616.434, 4409438.756, Mallorca Island, Spain) under the appellation Vi de la Terra Mallorca, and it is organic certified. The experimental plot comprised 2.4 ha with Merlot (clone 181) and Callet (local red cultivar) cultivars, both grafted on SO4 rootstock (clone 5). The plantation was settled down in 1999 with a training system maintained as organically certified, and has a density of 3200 plants per ha (2.5 between rows and 1.25 m within rows). A drip irrigation system is available with one drip per plant (2.3 L/m2). The irrigation system allows adjustment according to demand, watering only in the driest months of July and August on a weekly basis.
The soil in the area comes from marls and limestone-marls of the Late Jurassic and Cretaceous periods. Quaternary sedimentary materials are also found in the area. According to Reference Soil Groups [
36] the dominant soil is Calcaric Regosol. The soil is of chalky nature with a high-water retention capacity, high clay content (USDA, sand 17%, silt 31%, clay 52%), alkaline (pH 8.30), total organic carbon of 20.7 g/Kg, total nitrogen of 1.6 g/Kg, C/N ratio of 7.50, and cation exchange capacity CEC of 259.7 meq/Kg.
The climatic conditions are typically Mediterranean, with mild winters and hot dry summers. However, as the property is located quite close to the sea (7 km), the conditions are milder than expected, both in winter and summer. Average annual rainfall is approximately 700 mm with a mean temperature of 16.5 °C.
Regarding the management of the soil in the vineyard, a spontaneous green cover is maintained in alternate rows (one not tilled, another tilled) in the alleys between vine rows (inter-rows). The cover is maintained in the central part of the inter-rows, while the vegetation between vine plants in the same row is removed several times a year by shallow cultivation in a strip about 1 m wide. In the alley, the green cover is mowed and added to the soil as a green organic fertilizer in spring. In this way, the height of the green cover manages to control the water stress in the vine plants during the end of spring and summer. The green cover is changed every several years from one row to the next, tilling the entire plot. At the time of sampling, all sampled plants were kept weed-free by shallow cultivation at the row and mowed the alleys.
2.1. Sampling and DNA Extraction
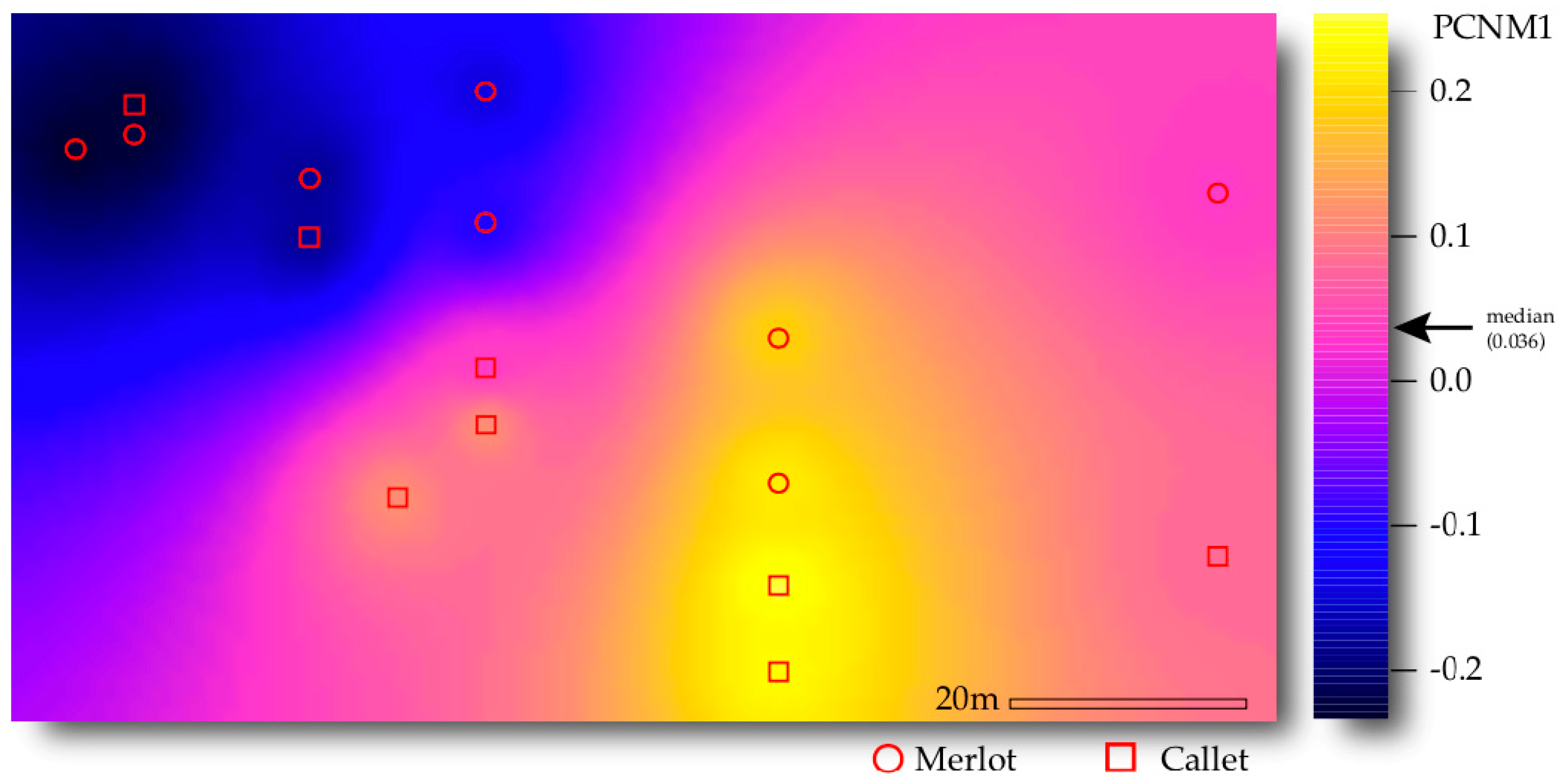
Soil sampling was performed 10 July 2018, during fruit development stage. Eight plants of each cultivar (either Callet or Merlot) were sampled and their spatial coordinates inside the studied plot recorded. Using a retro-scaler, a hole 40 cm in diameter was made as close as possible to the plant in the alley orientated to the north. Roots and 1 kg of soil were collected at 0–30 cm depth. Only the roots near the sampled plant were collected. The absence of surrounding weeds assured that the roots belonged to the vine. Samples were immediately placed in sterile bags and transported on ice for laboratory analyses.
Once in the laboratory, the thinnest roots were collected, discarding the thickest and oldest ones. The soil was sieved through 5.0 mm mesh, homogenized, and 20 g per sample kept at −20 °C. The roots were washed several times with abundant water until no soil was left and rinsed with distilled water. They were then cut into 0.5 cm pieces, homogenized, and stored in two 100 mg aliquots frozen at −80 °C.
Total soil DNA was extracted from samples of 0.25 g of sieved soil using the DNeasy® PowerSoil® Kit (Qiagen Inc., Mississauga, ON, Canada) in accordance with the manufacturer’s instructions. The quality of the extracts was assessed using NanoDrop spectrophotometer (Thermo Fisher Scientific, Delaware City, DE, USA) and then stored at −80 °C for further analysis.
Frozen roots were taken out of the freezer and quickly ground to a fine powder under liquid nitrogen using a mortar and pestle. Two genomic DNA extractions per root sample were performed (100 mg each) using the DNeasy Plant Mini Kit (Qiagen Inc., Mississauga, ON, Canada), following the manufacturer’s instructions.
2.2. DNA Amplification and Sequencing
Extracted DNA was processed to identify AM fungal taxa by Illumina Miseq-sequencing of the 18S rRNA gene using the Glomeromycota-specific primers NS31 [
37] and AML2 [
38]. Library preparation and Illumina sequencing were carried out at the IPBLN Genomics Facility (CSIC, Granada, Spain). Amplicon libraries were generated through a two-step PCR strategy. The first step was carried out in a final volume of 10 µL, containing 1× KAPA HiFi HotStart ReadyMix DNA polymerase (Roche Diagnostics, West Sussex, UK), 0.2 µM forward and reverse primers, and 10 ng of the template DNA. Cycling conditions were: 95 °C 3 min, (95 °C 30 s, 58 °C 30 s, 72 °C 30 s) × 30, 72 °C 5 min. PCRs were triplicated and pooled together. A second PCR step attached dual combinatorial indices and Illumina sequencing adapters using Nextera XT v2 index kit. PCR conditions were: 95 °C 3 min, (95 °C 30 s, 55 °C 30 s, 72 °C 30 s) × 8, 72 °C 5 min. All PCRs were validated through visualization on 1.8 % (
w/
v) agarose gel and purified using the NucleoMag® NGS Clean-up and Size Select Kit (Macherey-Nagel, Düren, Germany). Concentrations were measured on a Qubit® fluorometer (Thermo). Amplicons were equimolarly pooled and a final library mix was run on a Bioanalyzer HS DNA chip (Agilent, Santa Clara, CA, USA) to verify quality and size distribution. The library pool was then diluted and denatured as recommended by the Illumina MiSeq library preparation guide. The 300 × 2 nt paired-end sequencing was conducted on a MiSeq sequencer. Samples were demultiplexed, and barcodes were removed and returned as individual per-sample fastq files from the sequencing facility.
2.3. Bioinformatic Analyses
The initial 4,617,181 MiSeq sequences were analyzed with the amplicon sequence variant (ASV, hereafter) analysis pipeline known as Divisive Amplicon Denoising Algorithm (DADA2 v. 1.8.; Ref. [
39]). Briefly, forward and reverse sequences were trimmed to 295 and 290 bp, respectively. Primers were removed and a quality score set up to a minimum of 2. Sequences were dereplicated to keep unique sequences, and the error rate model inferred and used to implement the sample inference algorithm to remove Illumina sequencing errors. Forward and reverse reads were merged, and the sequence abundance table generated. Chimeric sequences based on the local dataset were removed (5.06% of quality filtered and merged reads). DADA2 gave 860 ASVs comprising 3,443,644 non-chimeric reads. The taxonomic assignment was determined for each ASV against the 16S/18S SILVA release 132 ([
40] accessed 09/2018) using the RDP algorithm [
41]. The SILVA database includes only a small set of representative sequences from Glomeromycota. Therefore, to improve taxonomic assignment, we amended the SILVA database with every classified sequence (i.e., those identified as a virtual taxa) in the Glomeromycotan specific database MaarjAM [
42] (accessed January 2019). This database comprised 28,137 sequences including their taxonomic assignment. The taxonomic assignment was then reassessed against this combined database using RDP, and non-Glomeromycotan sequences were discarded, resulting in 636 ASVs that comprised 3,296,623 reads.
To remove further errors that were thought not to be removed by standard DADA2 pipelines, we then applied LULU algorithm [
43] and obtained 240 corrected ASVs. Since DADA2-LULU infers unique original sequences in the DNA template, the result could correspond to an infra-specific level. Thus, the 240 ASVs were clustered by blasting against MaarjAM and named as the corresponding virtual taxa (VT, hereafter) when showing an identity higher than 97%. Those ASVs with low query cover (<90%) or low E-value during the blast were discarded (4/240). ASVs non-fitting at a minimum of 97% were aligned together with the rest of ASVs using MAFFT [
44] and clustered at 97% using VSEARCH [
45] implemented in MOTHUR [
46]. Those ASVs clustering with VT-named ASVs were added to the existing cluster and those clustering alone were considered as new VTs. With 3,295,222 reads, we finally obtained 56 VT, five of which were defined as new VT non-included in MaarjAM database. The bioinformatic pipeline is available as
supplementary material (Supplementary Material, Methods S1).
Data were deposited and are available in the Sequence Read Archive under Bioproject ID PRJNA679172. Representative sequences of the detected VT were deposited in GenBank under the accession numbers MW285643-MW285698. Processed raw data is available as
Supplementary Material, Data S1.
2.4. Statistical Analyses
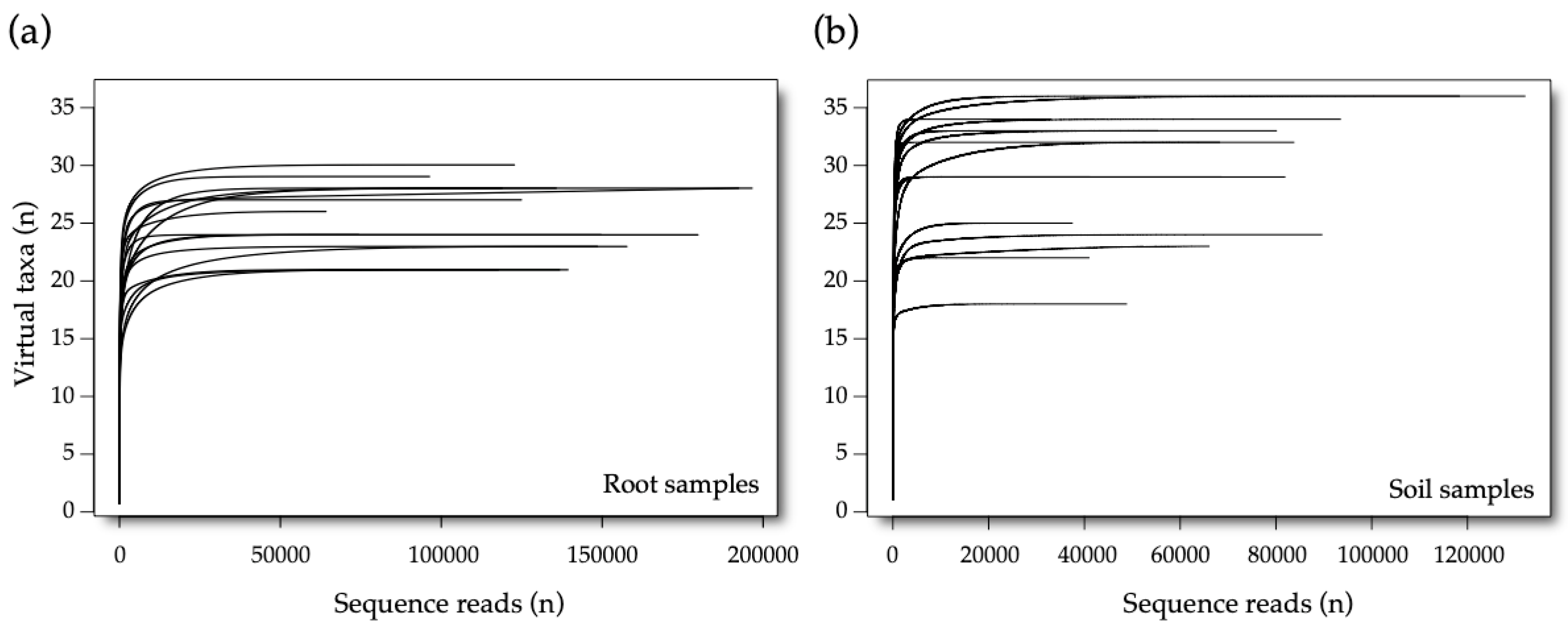
Prior to subsequent analyses, the sequencing information of the two subreplicates per root sample were pooled into a unique sample. The diversity coverage of the sequencing was checked by visualizing rarefaction curves by means of the
rarecurve function (
vegan R package [
47]).
To build the VT abundance matrix, read counts per VT and sample were used as a proxy of abundance. The VT abundance matrix was subjected to Hellinger transformation for subsequent analyses [
48].
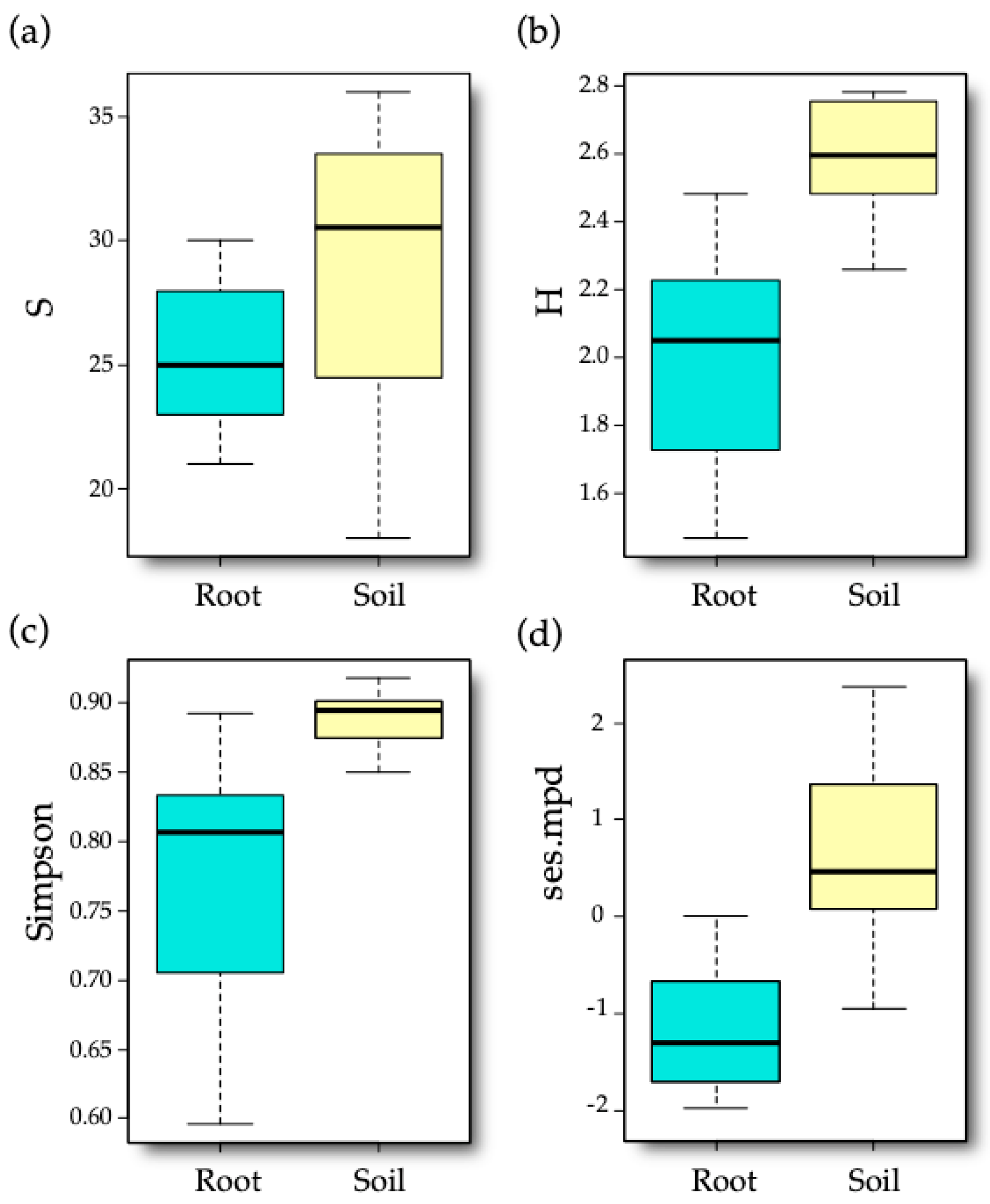
The VT abundance matrix was relativized to total row sums and used to obtain VT richness, Simpson (1-D) dominance, and Shannon indices at a sample level. The phylogenetic diversity was obtained by calculating the standardized effect size of the mean pairwise phylogenetic distance (ses.mpd) of AM fungal communities [
49]. For that, the most abundant ASV per VT was selected as representative sequences and aligned using MAFFT 7.0. The Tamura-Nei nucleotide substitution model with a discrete gamma distribution was found to be the best fitted using MEGA X [
50], and it was used to correct the evolutionary distance matrix between aligned sequences. The ses.mpd was calculated using the VT abundance matrix plus the evolutionary distance matrix of the VTs in each sample and compared to 999 null communities obtained using the independent swap algorithm which maintains species occurrence frequency and sample species richness (
ses.mpd function,
picante R package [
51]). The mean values of ses.mpd per treatment were then used to judge the clustering or segregation against null communities. Significance of the calculated index was assessed with a
t-test.
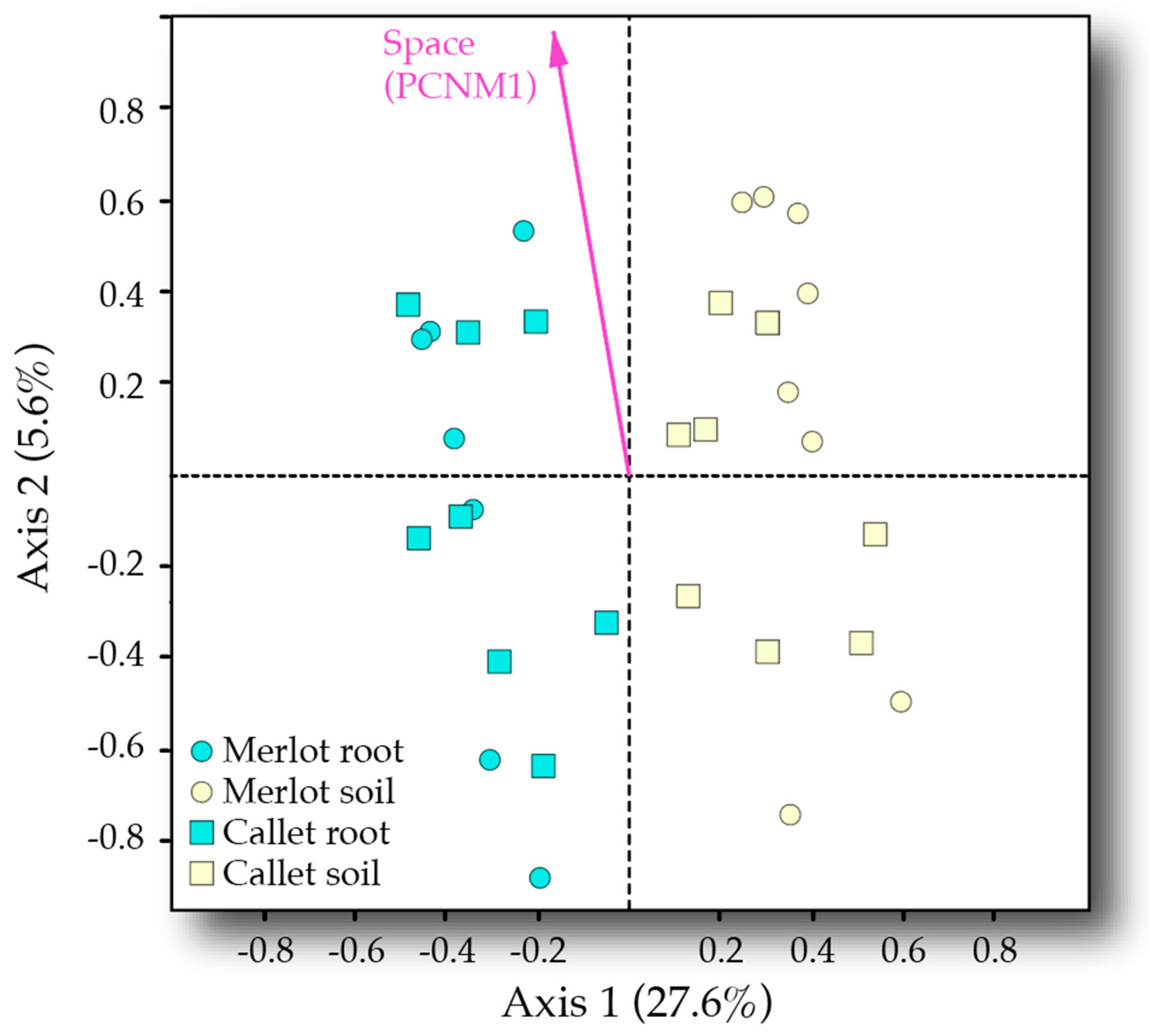
The spatial distribution of samples was decomposed via principal coordinates of neighbor matrices (PCNM). The significance of PCNM axes with positive eigenvalues on VT abundance distribution was evaluated by means of permutational multivariate analysis of variance (PERMANOVA, McArdle and Anderson 2001,
adonis function,
vegan R package [
47]), using 999 permutations and Euclidean distance as measures of dissimilarity. This dissimilarity, as the abundance matrix was Hellinger-transformed, must be considered a Hellinger-based dissimilarity [
48]. Those non-significant axes were discarded from further analyses.
The impact of vineyard cultivar, sample type (either soil or root), and their interaction on the mentioned diversity indices was tested via generalized least-squares models (
gls function,
nlme R package [
52]), with variance structures applied for the sample type using the
varIdent function to account for heteroscedastic variance in the model residual [
53]. The models were also tested including spatial autocorrelation as covariate, both the
x and
y coordinates, and the selected first axis of PCNM decomposition.
The effects of the experimental variables: Cultivar, sample type, and spatial position (PCNM axes), and their interaction on community composition was addressed by means of PERMANOVA (999 permutations, Euclidean distance). As PERMANOVA is sensitive to changes in multivariate dispersion among samples, the
betadisper function (
vegan R package) was used to assess differences in multivariate dispersion across factor levels. When finding significant interactions across explanatory variables, partial PERMANOVAs were run for each factor level. In the case of PCNM axes, their values were split in quartiles and analyzed in four different PERMANOVAs. To visualize the found patterns, we used a redundancy analysis (RDA) [
54] ordination plot constraining by the mentioned experimental variables (
rda function,
vegan R package).
AM fungal VT indicative of particular groups of samples were identified using Dufrêne-Legendre indicator species analysis [
55], implemented by the
indval() function (
labdsv R package [
56]).

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}


