Influence of Calcium Silicate on the Chemical Properties of Pleurotus ostreatus var. florida (Jacq.) P. Kumm

 ,
,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Mushroom Strains
2.3. Substrate and Supplementation
2.4. Growing Cycle
2.5. Influence of Calcium Silicate in Biological Production Yields
2.5.1. Biological Efficiency
2.5.2. Yield Ratio
2.5.3. Productivity Rate
2.6. Chemical Composition
2.6.1. Soluble Sugars
2.6.2. Tocopherols
2.6.3. Ergosterol and Vitamin D2
2.6.4. Fatty Acids
2.7. Statistical Analysis
3. Results and Discussion
3.1. Biological Efficiency and Crop Yield Ratio
3.2. Effect of Supplementation on Individual Compounds
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nölle, N.; Argyropoulos, D.; Ambacher, S.; Müller, J.; Biesalski, H.K. Vitamin D2 enrichment in mushrooms by natural or artificial UV-light during drying. LWT Food Sci. Technol. 2017, 85, 400–404. [Google Scholar] [CrossRef]
- Naim, L.; Alsanad, M.A.; El Sebaaly, Z.; Shaban, N.; Fayssal, S.A.; Sassine, Y.N. Variation of Pleurotus ostreatus (Jacq. Ex Fr.) P. Kumm. (1871) performance subjected to differentdoses and timings of nano-urea. Saudi J. Biol. Sci. 2020, 27, 1573–1579. [Google Scholar] [CrossRef] [PubMed]
- Lavelli, V.; Proserpio, C.; Gallotti, F.; Laureati, M.; Pagliarini, E. Circular reuse of bio-resources: The role of: Pleurotus spp. in the development of functional foods. Food Funct. 2018, 9, 1353–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teimoori, B.B.; Pourianfar, H.R.; Moeini, M.J.; Janpoor, J. Chemically and physically induced mutagenesis in basidiospores of oyster mushroom Pleurotus ostreatus var. florida. Int. J. Adv. Res. 2014, 2, 915–921. [Google Scholar]
- Oliveira, R.; David, G.Q.; Silva, M.P.D.; Berti, C.L.F.; Peres, W.M. Pleurotus florida production of the Amazon southern different substrates. Rev. Agrotecnologia 2018, 9, 28–35. [Google Scholar] [CrossRef]
- Royse, D.J.; Baars, J.; Tan, Q. Current Overview of Mushroom Production in the World. In Edible and Medicinal Mushrooms: Technology and Applications; Zied, D.C., Pardo-Giménez, A., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; Volume 2010, pp. 5–13. [Google Scholar]
- Gonçalves, F.J.; de Araújo, L.G.; da Silva, G.B.; de Filippi, M.C.C. Controle Químico da Brusone em Arroz de Terras Altas: Efeitos Nos Fungos Não Alvos do Filoplano. Pesqui. Agropecu. Trop. 2012, 42, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Thongsook, T.; Kongbangkerd, T. Influence of calcium and silicon supplementation into Pleurotus ostreatus substrates on quality of fresh and canned mushrooms. Food Sci. Technol. Int. 2011, 17, 351–365. [Google Scholar] [CrossRef]
- Kumara, B.H.; Yogendra, N.D.; Prakash, N.B.; Kumar, A. Effect of calcium silicate and need based nitrogen on pests management in aerobic rice (Oryza sativa L.). Int. J. Plant Prot. 2016, 9, 133–136. [Google Scholar] [CrossRef]
- Parthiban, P.; Chinniah, C.; Baskaran, R.K.M.; Muthukrishnan, N.; Rajavel, D.S.; Karthick, K.S. Influence of calcium silicate application on the population of Aproaerema modicella Deventer (Lepidoptera: Gelechiidae) on groundnut. Entomon 2018, 43, 231–236. [Google Scholar] [CrossRef]
- Parthiban, P.; Chinniah, C.; Baskaran, K.M.; Suresh, K.; Karthick, S. Influence of Calcium Silicate Application on the Population of Sucking Pests of Groundnut (Arachis hypogaea L.). Silicon 2019, 11, 1687–1692. [Google Scholar] [CrossRef]
- Pozza, A.A.A.; Carvalho, J.G.; Guimares, P.T.G.; Figueiredo, F.C.; Araújo, A.R. Suprimento do silicato de cálcio e a eficiência nutricional de variedades de cafeeiro. Rev. Bras. Cienc. Solo 2009, 33, 1705–1714. [Google Scholar] [CrossRef] [Green Version]
- Korndorfer, A.P.; Cherry, R.; Nagata, R. Effect of calcium silicate on feeding and development of tropical sod webworms (Lepidoptera: Pyralidae). Fla. Entomol. 2004, 87, 393–395. [Google Scholar] [CrossRef]
- Tubana, B.S.; Babu, T.; Datnoff, L.E. A review of silicon in soils and plants and its role in US agriculture: History and future perspectives. Soil Sci. 2016, 181, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Saigusa, M.; Yamamoto, A.; Shibuya, K. Agricultural Use of Porous Hydrated Calcium Silicate: Effect of porous hydrated calcium silicate on resistance of rice plant (Oryza sativa L.) to rice blast (Pyricularia olyzae). Plant Prod. Sci. 2000, 3, 51–54. [Google Scholar] [CrossRef] [Green Version]
- Phattharachindanuwong, C.; Hansupalak, N.; Plank, J.; Chisti, Y. Template-assisted facile synthesis and characterization of hollow calcium silicate hydrate particles for use as reflective materials. Mater. Res. Bull. 2018, 97, 343–350. [Google Scholar] [CrossRef]
- Keeping, M.G.; Meyer, J.H. Calcium silicate enhances resistance of sugarcane to the African stalk borer Eldana saccharina Walker (Lepidoptera: Pyralidae). Agric. For. Entomol. 2002, 4, 265–274. [Google Scholar] [CrossRef]
- Zied, D.C.; Pardo-Giménez, A.; de Oliveira, G.A.; Carrasco, J.; Zeraik, M.L. Study of Waste Products as Supplements in the Production and Quality of Pleurotus ostreatus var. florida. Indian J. Microbiol. 2019, 59, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.C.N.d.; Kopytowski Filho, J.; Minhoni, M.T.D.A.; Coutinho, L.N.; Figueiredo, M.B. Productivity, biological efficiency, and number of Agaricus blazei mushrooms grown in compost in the presence of Trichoderma sp. and Chaetomium olivacearum contaminants. Braz. J. Microbiol. 2007, 38, 243–247. [Google Scholar] [CrossRef]
- Zied, D.C.; Prado, E.P.; Dias, E.S.; Pardo, J.E.; Pardo-Gimenez, A. Use of peanut waste for oyster mushroom substrate supplementation—Oyster mushroom and peanut waste. Braz. J. Microbiol. 2019, 50, 1021–1029. [Google Scholar] [CrossRef]
- Royse, D.J.; Rhodes, T.W.; Ohga, S.; Sanchez, J.E. Yield, mushroom size and time to production of Pleurotus cornucopiae (oyster mushroom) grown on switch grass substrate spawned and supplemented at various rates. Bioresour. Technol. 2004, 91, 85–91. [Google Scholar] [CrossRef]
- Pardo-Giménez, A.; Catalán, L.; Carrasco, J.; Álvarez-Ortí, M.; Zied, D.; Pardo, J. Effect of supplementing crop substrate with defatted pistachio meal on Agaricus bisporus and Pleurotus ostreatus production. J. Sci. Food Agric. 2016, 96, 3838–3845. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, J.; Zied, D.C.; Pardo, J.E.; Preston, G.M.; Pardo-Giménez, A. Supplementation in mushroom crops and its impact on yield and quality. AMB Express 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, L.; Pereira, E.; Calhelha, R.C.; Dueñas, M.; Carvalho, A.M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Bioactivity and chemical characterization in hydrophilic and lipophilic compounds of Chenopodium ambrosioides L. J. Funct. Foods 2013, 5, 1732–1740. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Mau, J.L.; Huang, S.J. Enhancement of antioxidant properties and increase of content of vitamin D2 and non-volatile components in fresh button mushroom, Agaricus bisporus (higher basidiomycetes) by γ-irradiation. Int. J. Med. Mushrooms 2014, 16, 137–147. [Google Scholar] [CrossRef]
- Barreira, J.C.M.; Oliveira, M.B.P.P.; Ferreira, I.C.F.R. Development of a Novel Methodology for the Analysis of Ergosterol in Mushrooms. Food Anal. Methods 2014, 7, 217–223. [Google Scholar] [CrossRef]
- Fernandes, Â.; Barros, L.; Martins, A.; Herbert, P.; Ferreira, I.C.F.R. Nutritional characterisation of Pleurotus ostreatus (Jacq. ex Fr.) P. Kumm. produced using paper scraps as substrate. Food Chem. 2015, 169, 396–400. [Google Scholar] [CrossRef] [Green Version]
- Bouzgarrou, C.; Amara, K.; Reis, F.S.; Barreira, J.C.M.; Skhiri, F.; Chatti, N.; Martins, A.; Barros, L.; Ferreira, I.C.F.R. Incorporation of tocopherol-rich extracts from mushroom mycelia into yogurt. Food Funct. 2018, 9, 3166–3172. [Google Scholar] [CrossRef] [Green Version]
- Bikle, D.D. Vitamin D metabolism, mechanism of action, and clinical applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Kozarski, M.; Klaus, A.; Jakovljevic, D.; Todorovic, N.; Vunduk, J.; Petrović, P.; Niksic, M.; Vrvic, M.M.; Van Griensven, L. Antioxidants of edible mushrooms. Molecules 2015, 20, 19489–19525. [Google Scholar] [CrossRef] [Green Version]
- Barros, L.; Correia, D.M.; Ferreira, I.C.F.R.; Baptista, P.; Santos-Buelga, C. Optimization of the determination of tocopherols in Agaricus sp. edible mushrooms by a normal phase liquid chromatographic method. Food Chem. 2008, 110, 1046–1050. [Google Scholar] [CrossRef]
- Nieto, I.J.; Carolina, C.A. The effect of different substrates on triterpenoids and fatty acids in fungi of the genus pleurotus. J. Chil. Chem. Soc. 2013, 58, 1580–1583. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Presidente Prudente (PP) | ||||||
|---|---|---|---|---|---|---|
| Mushroom Weight (g) | Number of Mushrooms | Biological Efficiency (%) | Yield Ratio (%) | Productivity Rate (% per Day) | ||
| Harvest number (HN) | First flush | 167 ± 52 * | 50 ± 21 * | 104 ± 32 * | 8 ± 2 * | 4 ± 1 * |
| Second flush | 110 ± 41 | 35 ± 16 | 69 ± 25 | 5 ± 2 | 2.0 ± 0.8 | |
| p-value (n = 10) | Student T test | <0.001 | 0.009 | <0.001 | <0.001 | <0.001 |
| Calcium silicate supplementation (CS) | Control | 164 ± 65 | 40 ± 22 | 103 ± 41 | 8 ± 3 | 3 ± 2 |
| 0.5% | 147 ± 59 | 43 ± 16 | 92 ± 36 | 7 ± 3 | 3 ± 1 | |
| 1% | 129 ± 45 | 47 ± 27 | 81 ± 28 | 6 ± 2 | 3 ± 1 | |
| 2% | 124 ± 52 | 39 ± 21 | 77 ± 32 | 6 ± 2 | 3 ± 1 | |
| 4% | 129 ± 48 | 43 ± 16 | 80 ± 30 | 6 ± 2 | 3 ± 1 | |
| p-value (n = 25) | Tukey’s HSD test | 0.296 | 0.897 | 0.296 | 0.296 | 0.571 |
| HN×CS (n = 50) | p-value | 0.826 | 0.332 | 0.826 | 0.826 | 0.697 |
| Mogi-das-Cruzes (MC) | ||||||
| Harvest number (HN) | First flush | 172 ± 27 * | 46 ± 6 * | 108 ± 13 * | 9 ± 1 * | 4 ± 1 * |
| Second flush | 135 ± 68 | 33 ± 4 | 84 ± 42 | 7 ± 2 | 2 ± 1 | |
| p-value (n = 10) | Student T test | 0.044 | 0.031 | 0.044 | 0.044 | <0.001 |
| Calcium silicate supplementation (CS) | Control | 174 ± 79 | 48 ± 30 | 108 ± 49 | 9 ± 4 | 4 ± 2 |
| 0.5% | 156 ± 43 | 35 ± 18 | 97 ± 27 | 8 ± 2 | 3 ± 1 | |
| 1% | 159 ± 50 | 41 ± 12 | 100 ± 31 | 8 ± 2 | 3 ± 1 | |
| 2% | 156 ± 44 | 43 ± 26 | 97 ± 27 | 8 ± 2 | 3 ± 1 | |
| 4% | 124 ± 94 | 30 ± 19 | 78 ± 59 | 6 ± 5 | 3 ± 2 | |
| p-value (n = 25) | Tukey’s HSD test | 0.529 | 0.350 | 0.529 | 0.529 | 0.708 |
| HN×CS (n = 50) | p-value | 0.455 | 0.264 | 0.455 | 0.455 | 0.430 |
| Presidente Prudente (PP) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
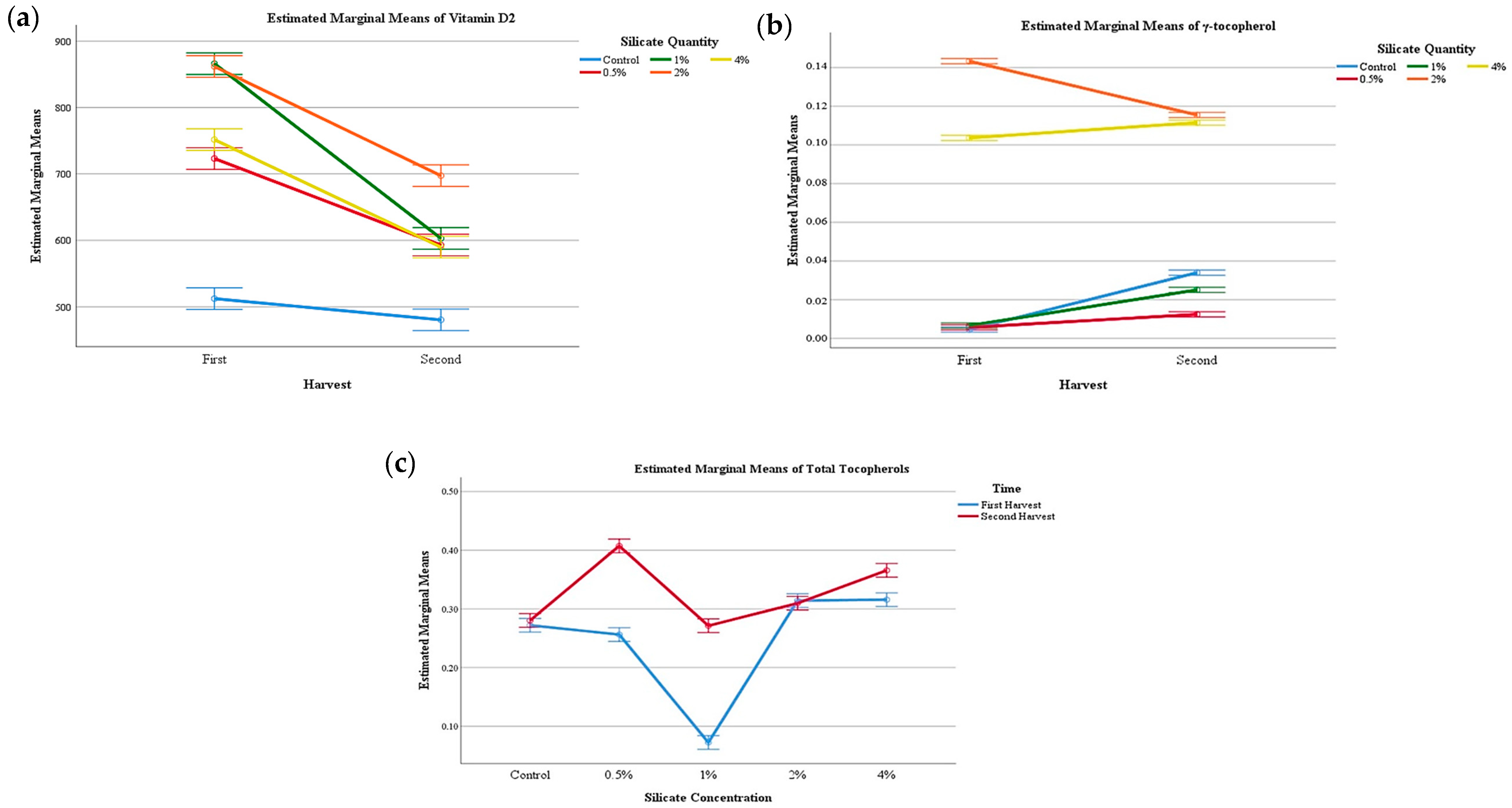
| Ergosterol (mg/100 g) | Vitamin D2 (µg/100 g) | Fructose (g/100 g) | Mannitol (g/100 g) | Trehalose (g/100 g) | Soluble Sugars (g/100 g) | α-Tocopherol (mg/100 g) | β-Tocopherol (mg/100 g) | γ-Tocopherol (mg/100 g) | Total Tocopherols (mg/100 g) | ||
| Harvest number (HN) | First flush | 107 ± 6 | 743 ± 133 | 0.16 ± 0.03 | 2.2 ± 0.2 | 19 ± 2 | 22 ± 2 | 0.002 ± 0.001 | 0.26 ± 0.03 | 0.05 ± 0.06 | 0.31 ± 0.07 |
| Second flush | 107 ± 8 | 593 ± 73 | 0.2 ± 0.1 | 2.2 ± 0.6 | 15 ± 4 | 17 ± 4 | 0.002 ± 0.001 | 0.3 ± 0.07 | 0.06 ± 0.04 | 0.36 ± 0.05 | |
| p-value (n = 10) | Student T test | 0.288 | 0.031 | 0.007 | 0.811 | <0.001 | <0.001 | 0.580 | <0.001 | <0.001 | <0.001 |
| Calcium silicate supplementation (CS) | Control | 101 ± 8 | 496 ± 27 | 0.20 ± 0.03 | 2.8 ± 0.5 | 15 ± 3 | 18 ± 3 | 0.002 ± 0.001 | 0.32 ± 0.03 | 0.02 ± 0.01 | 0.34 ± 0.04 |
| 0.5% | 109 ± 6 | 658 ± 71 | 0.13 ± 0.03 | 1.8 ± 0.4 | 18 ± 3 | 20 ± 3 | 0.002 ± 0.001 | 0.24 ± 0.04 | 0.009 ± 0.004 | 0.25 ± 0.04 | |
| 1% | 110 ± 1 | 734 ± 144 | 0.19 ± 0.08 | 1.91 ± 0.06 | 16 ± 8 | 18 ± 8 | 0.002 ± 0.001 | 0.34 ± 0.08 | 0.02 ± 0.01 | 0.36 ± 0.09 | |
| 2% | 113 ± 1 | 779 ± 90 | 0.16 ± 0.03 | 2.0 ± 0.2 | 19 ± 1 | 21 ± 2 | 0.001 ± 0.0001 | 0.26 ± 0.01 | 0.13 ± 0.01 | 0.39 ± 0.01 | |
| 4% | 101 ± 4 | 671 ± 89 | 0.2 ± 0.1 | 2.3 ± 0.2 | 18 ± 2 | 21 ± 2 | 0.001 ± 0.0001 | 0.23 ± 0.03 | 0.108 ± 0.004 | 0.34 ± 0.03 | |
| p-value (n = 25) | Tukey’s HSD test | <0.001 | <0.001 | 0.006 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| HN×CS (n = 50) | p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Mogi-das-Cruzes (MC) | |||||||||||
| Harvest number (HN) | First flush | 111 ± 6 | 899 ± 167 | 0.14 ± 0.03 | 1.9 ± 0.2 | 19±2 | 21 ± 2 | 0.003±0.001 * | 0.2 ± 0.1 | 0.05 ± 0.03 | 0.25 ± 0.09 |
| Second flush | 116 ±10 | 829 ± 175 | 0.18 ± 0.08 | 1.8 ± 0.4 | 17 ± 4 | 19 ± 4 | 0.0018 ± 0.0001 | 0.28 ± 0.06 | 0.04 ± 0.04 | 0.32 ± 0.05 | |
| p-value (n = 10) | Student T test | <0.001 | <0.001 | <0.001 | 0.062 | <0.001 | <0.001 | 0.009 | <0.001 | <0.001 | <0.001 |
| Calcium silicate supplementation (CS) | Control | 113 ± 4 | 555 ± 51 | 0.2 ± 0.1 | 2.3 ± 0.3 | 18 ± 2 | 20 ± 2 | 0.0025 ± 0.0009 | 0.261 ± 0.005 | 0.013 ± 0.003 | 0.28 ± 0.05 |
| 0.5% | 123 ± 1 | 951 ± 90 | 0.16 ± 0.03 | 1.8 ± 0.4 | 17 ± 5 | 19 ± 6 | 0.0022 ± 0.0003 | 0.32 ± 0.08 | 0.011 ± 0.003 | 0.33 ± 0.08 | |
| 1% | 100 ± 3 | 880 ± 50 | 0.15 ± 0.01 | 1.6 ± 0.2 | 16 ± 3 | 18 ± 3 | 0.002 ± 0.001 | 0.1 ± 0.1 | 0.04 ± 0.03 | 0.2 ± 0.1 | |
| 2% | 117 ± 7 | 1022 ± 31 | 0.10 ± 0.02 | 1.63 ± 0.05 | 18 ± 2 | 19 ± 2 | 0.003 ± 0.001 | 0.23 ± 0.02 | 0.08 ± 0.02 | 0.31 ± 0.01 | |
| 4% | 114 ± 6 | 913 ± 30 | 0.19 ± 0.06 | 2.0 ± 0.2 | 20 ± 2 | 22 ± 2 | 0.0014 ± 0.0007 | 0.25 ± 0.03 | 0.087 ± 0.006 | 0.34 ± 0.03 | |
| p-value (n = 25) | Tukey’s HSD test | <0.001 | <0.001 | 0.006 | <0.001 | <0.001 | <0.001 | 0.130 | <0.001 | <0.001 | <0.001 |
| HN×CS (n = 50) | p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.058 | <0.001 | <0.001 | <0.001 |
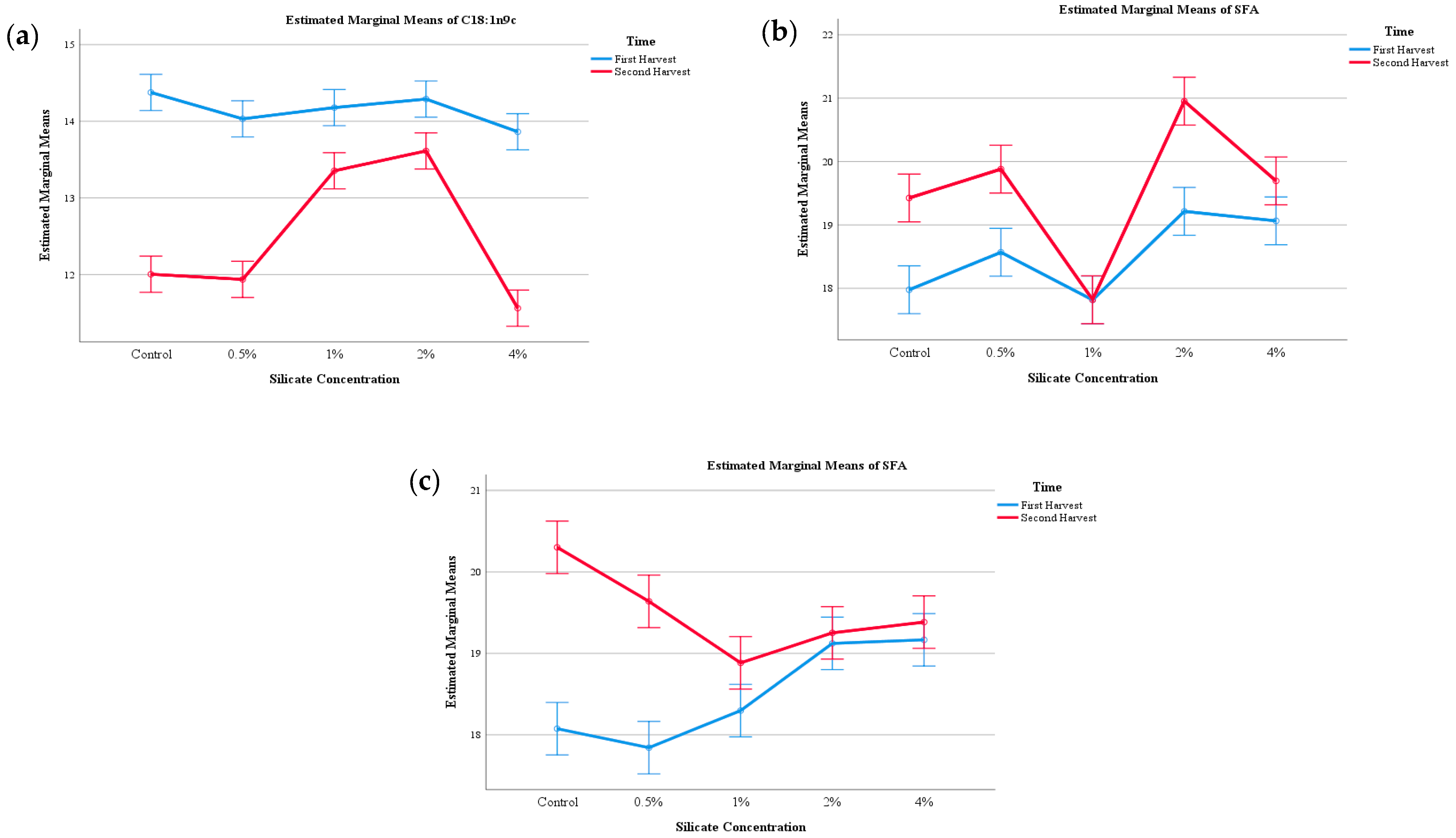
| Presidente Prudente (PP) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| C15:0 | C16:0 | C18:0 | C18:1n9C | C18:2n6c | SFAs | MUFAs | PUFAs | ||
| Harvest number (HN) | First flush | 1.4 ± 0.1 | 11.7 ± 0.4 | 2.8 ± 0.2 | 14.1 ± 0.3 | 65.8 ± 0.7 | 18.5 ± 0.6 | 14.6 ± 0.3 | 66.8 ± 0.7 |
| Second flush | 1.7 ± 0.2 | 11.9 ± 0.5 | 2.7 ± 0.2 | 12.5 ± 0.9 | 66 ± 1 | 19 ± 1 | 13.0 ± 0.9 | 67 ± 1 | |
| p-value (n = 10) | Student T test | <0.001 | 0.025 | <0.001 | <0.001 | 0.002 | <0.001 | <0.001 | 0.006 |
| Calcium silicate supplementation (CS) | Control | 1.7 ± 0.3 | 11.7 ± 0.2 | 2.79 ± 0.06 | 13 ± 1 | 66.6 ± 0.7 | 18.7 ± 0.8 | 14 ± 1 | 67.6 ± 0.6 |
| 0.5% | 1.5 ± 0.4 | 11.8 ± 0.2 | 3.0 ± 0.1 | 13 ± 1 | 66.4 ± 0.4 | 19.2 ± 0.7 | 13 ±1 | 67.3 ± 0.4 | |
| 1% | 1.3 ± 0.07 | 11.5 ± 0.2 | 2.7 ± 0.2 | 13.8 ± 0.4 | 66.9 ± 0.3 | 17.8 ± 0.1 | 14.3 ± 0.4 | 67.9 ± 0.4 | |
| 2% | 1.59 ± 0.07 | 12.1 ± 0.8 | 2.80 ± 0.07 | 13.9 ± 0.5 | 64.3 ± 0.8 | 20 ± 1 | 14.5 ± 0.4 | 65.4 ± 0.8 | |
| 4% | 1.6 ± 0.2 | 12.0 ± 0.6 | 2.65 ± 0.09 | 13.9 ± 0.5 | 66 ± 1 | 19.4 ± 0.5 | 13±1 | 67±1 | |
| p-value (n = 25) | Tukey’s HSD test | <0.001 | 0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| HN×CS (n = 50) | p-value | <0.001 | <0.001 | 0.001 | <0.001 | 0.002 | 0.001 | <0.001 | 0.001 |
| Mogi-das-Cruzes (MC) | |||||||||
| Harvest number (HN) | First flush | 1.29 ± 0.06 | 11.6 ± 0.3 | 2.7 ± 0.1 | 13.6 ± 0.6 | 66.4 ± 0.6 | 18.5 ± 0.6 | 14.1 ± 0.6 | 67.4 ± 0.6 |
| Second flush | 1.6 ± 0.1 | 12.4 ± 0.3 | 2.8 ± 0.2 | 13 ± 1 | 66 ± 1 | 19.4 ± 0.5 | 13 ± 1 | 67 ± 1 | |
| p-value (n = 10) | Student T test | <0.001 | <0.001 | 0.543 | <0.001 | 0.045 | <0.001 | <0.001 | 0.146 |
| Calcium silicate supplementation (CS) | Control | 1.5 ± 0.1 | 12.2 ± 0.4 | 2.7 ± 0.1 | 13.9 ± 0.2 | 65 ± 1 | 19 ± 1 | 14.3 ± 0.2 | 66 ± 1 |
| 0.5% | 1.4 ± 0.2 | 11.9 ± 0.6 | 2.7 ± 0.3 | 13 ± 1 | 66.8 ± 0.5 | 18 ± 1 | 13 ± 1 | 67.7 ± 0.5 | |
| 1% | 1.5 ± 0.3 | 11.9 ± 0.2 | 0.287 ± 0.03 | 13 ± 1 | 67.2 ± 0.8 | 18.6 ± 0.3 | 13.1 ± 0.1 | 68.3 ± 0.9 | |
| 2% | 1.39 ± 0.07 | 12.3 ± 0.4 | 2.7 ± 0.1 | 12.5 ± 0.1 | 66.6 ± 0.3 | 19.2 ± 0.2 | 13.1 ± 0.1 | 67.7 ± 0.3 | |
| 4% | 1.4 ± 0.1 | 11.9 ± 0.8 | 2.8 ± 0.1 | 14.0 ± 0.3 | 65.2 ± 0.8 | 19.2 ± 0.4 | 14.6 ± 0.4 | 66.1 ± 0.8 | |
| p-value (n = 25) | Tukey’s HSD test | <0.001 | 0.001 | <0.001 | <0.001 | <0.001 | 0.001 | <0.001 | <0.001 |
| HN×CS (n = 50) | p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoso, R.V.C.; Carocho, M.; Fernandes, Â.; Zied, D.C.; Cobos, J.D.V.; González-Paramás, A.M.; Ferreira, I.C.F.R.; Barros, L. Influence of Calcium Silicate on the Chemical Properties of Pleurotus ostreatus var. florida (Jacq.) P. Kumm. J. Fungi 2020, 6, 299. https://doi.org/10.3390/jof6040299
Cardoso RVC, Carocho M, Fernandes Â, Zied DC, Cobos JDV, González-Paramás AM, Ferreira ICFR, Barros L. Influence of Calcium Silicate on the Chemical Properties of Pleurotus ostreatus var. florida (Jacq.) P. Kumm. Journal of Fungi. 2020; 6(4):299. https://doi.org/10.3390/jof6040299
Chicago/Turabian StyleCardoso, Rossana V. C., Márcio Carocho, Ângela Fernandes, Diego Cunha Zied, Juan Diego Valenzuela Cobos, Ana M. González-Paramás, Isabel C. F. R. Ferreira, and Lillian Barros. 2020. "Influence of Calcium Silicate on the Chemical Properties of Pleurotus ostreatus var. florida (Jacq.) P. Kumm" Journal of Fungi 6, no. 4: 299. https://doi.org/10.3390/jof6040299