Consensus Multilocus Sequence Typing Scheme for Pneumocystis jirovecii

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Investigated Loci and Typing Schemes
2.1. Nuclear rRNA Gene Cluster
2.2. Mitochondrial Genes
2.3. Nuclear Genes
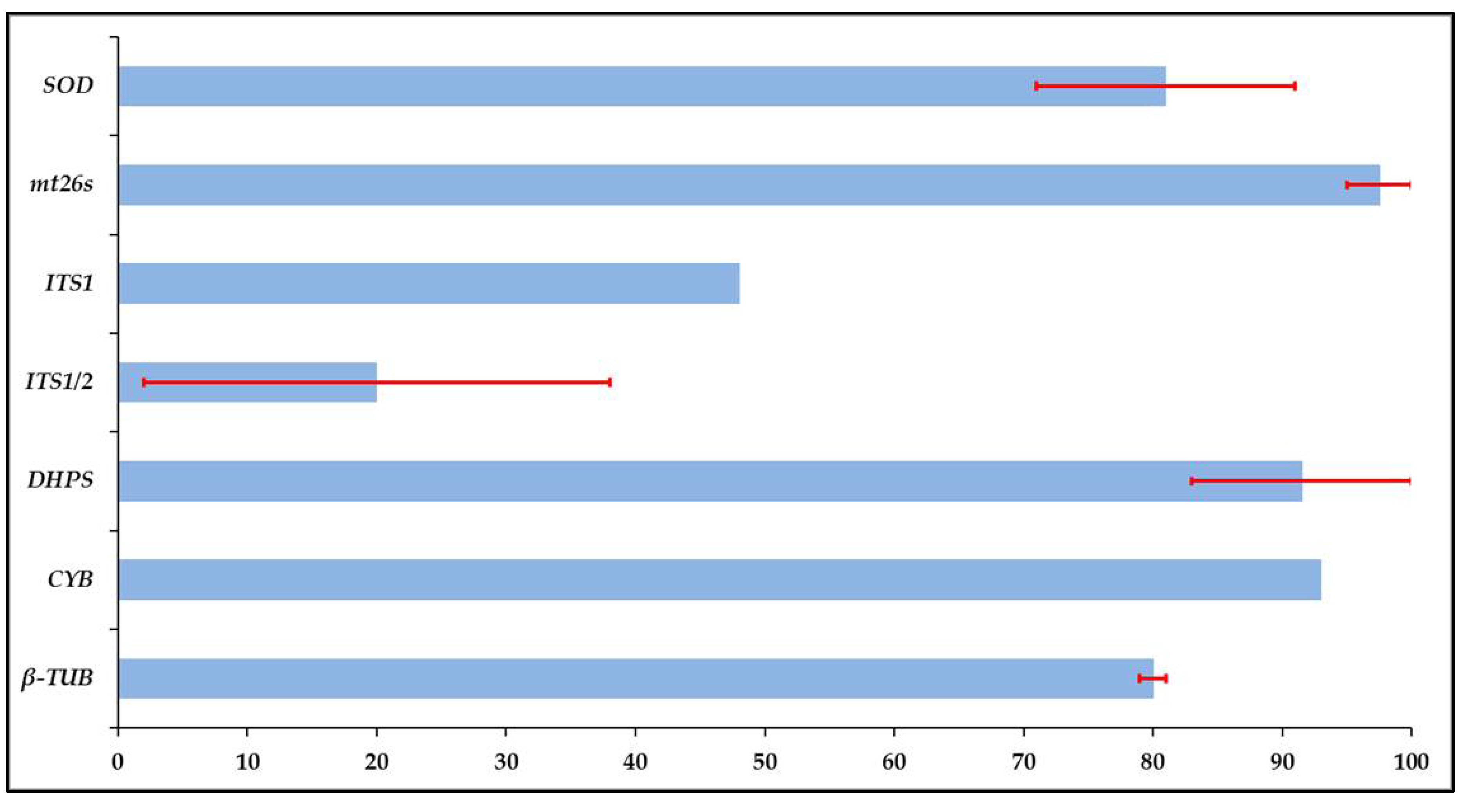
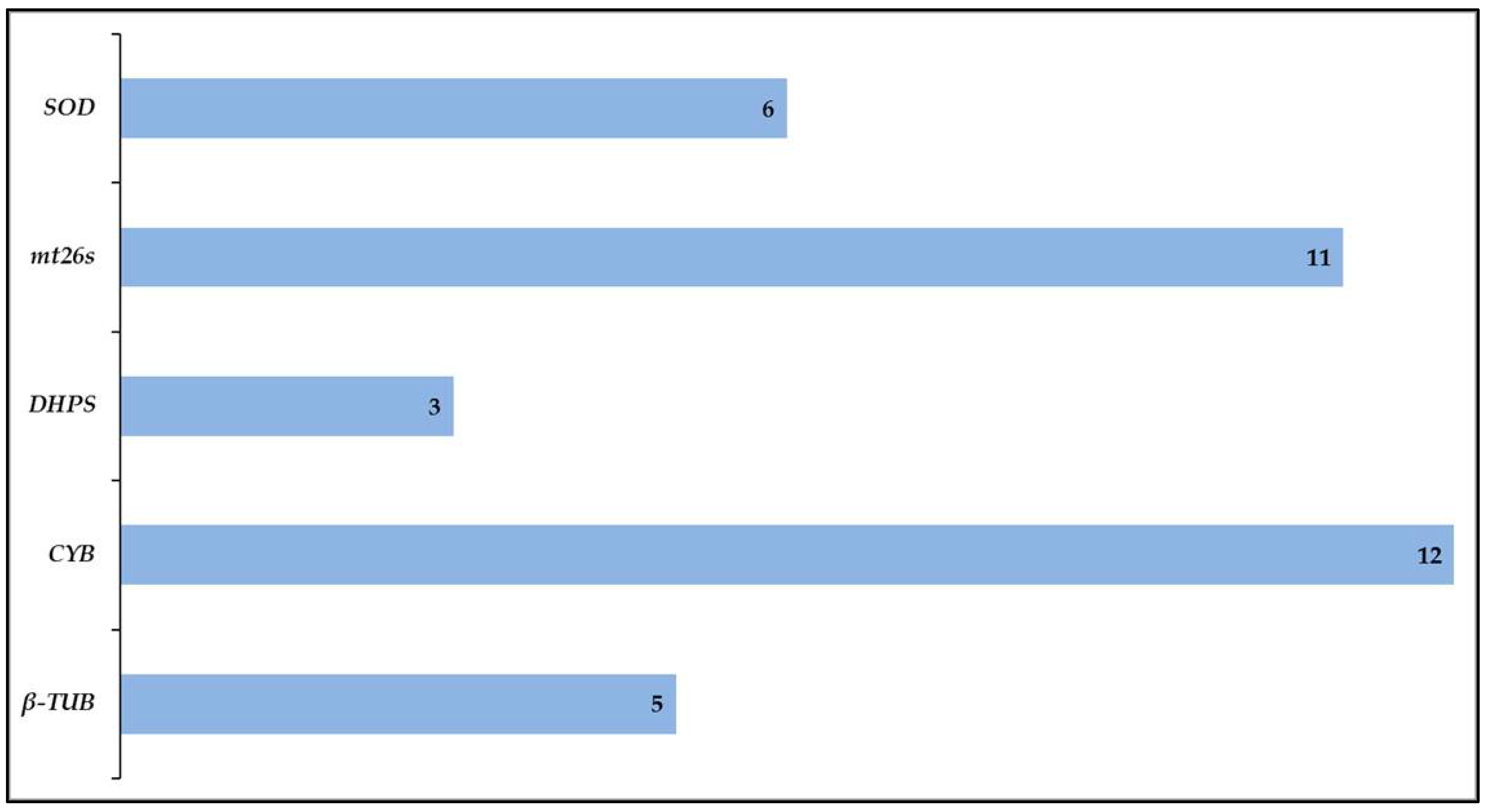
3. Amplification Rate and Variation of Target Loci
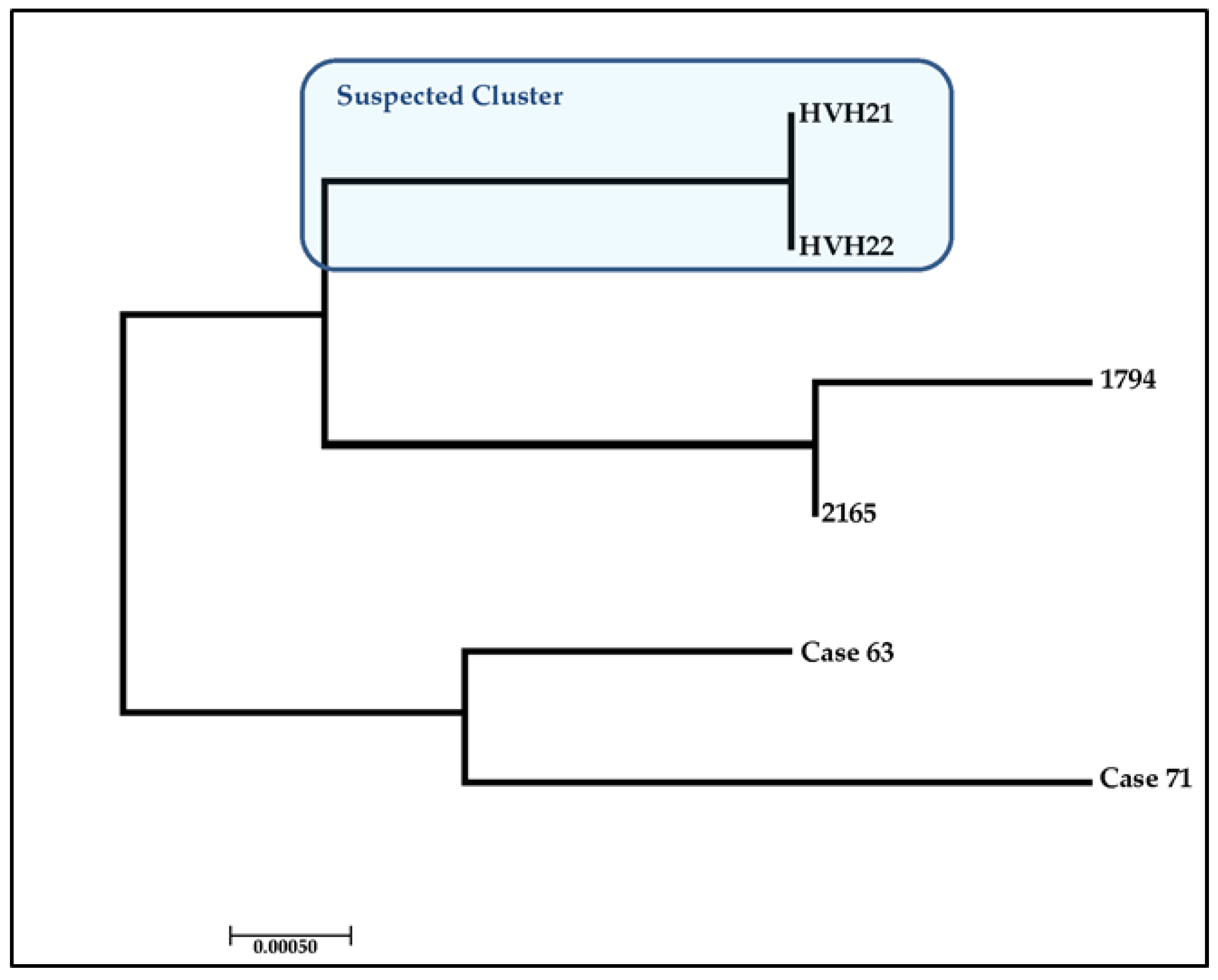
4. Case Study: Assessing the Ability to Discriminate between Clinical Isolates
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meenakshi, K.; Gowtham, R.R.; Usha, K. Pneumocystis jirovecii Pneumonia: A Revisit to the Old Malady. J. Clin. Diagn. Res. 2019, 13, DE01–DE08. [Google Scholar] [CrossRef]
- Morris, A.; Lundgren, J.D.; Masur, H.; Walzer, P.D.; Hanson, D.L.; Frederick, T.; Huang, L.; Beard, C.B.; Kaplan, J.E. Current epidemiology of Pneumocystis pneumonia. Emerg. Infect. Dis. 2004, 10, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Phipps, L.M.; Chen, S.C.-A.; Kable, K.; Halliday, C.L.; Firacative, C.; Meyer, W.; Wong, G.; Nankivell, B.J. Nosocomial Pneumocystis jirovecii pneumonia: Lessons from a cluster in kidney transplant recipients. Transplantation 2011, 92, 1327–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokulska, M.; Kicia, M.; Wesołowska, M.; Hendrich, A.B. Pneumocystis jirovecii—From a commensal to pathogen: Clinical and diagnostic review. Parasitol. Res. 2015, 114, 3577–3585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maitte, C.; Leterrier, M.; Le Pape, P.; Miegeville, M.; Morio, F. Multilocus sequence typing of Pneumocystis jirovecii from clinical samples: How many and which loci should be used? J. Clin. Microbiol. 2013, 51, 2843–2849. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.-J.; Bartlett, M.S.; Shaw, M.M.; Queener, S.F.; Smith, J.W.; Ortiz-Rivera, M.; Leibowitz, M.J.; Lee, C.-H. Typing of Pneumocystis carinii strains that infect humans based on nucleotide sequence variations of internal transcribed spacers of rRNA genes. J. Clin. Microbiol. 1994, 32, 2904–2912. [Google Scholar] [CrossRef] [Green Version]
- Tsolaki, A.G.; Miller, R.F.; Underwood, A.P.; Banerji, S.; Wakefield, A.E. Genetic diversity at the internal transcribed spacer regions of the rRNA operon among isolates of Pneumocystis carinii from AIDS patients with recurrent pneumonia. J. Infect. Dis. 1996, 174, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Keely, S.P.; Stringer, J.R. Multi-locus genotype switching in Pneumocystis carinii sp. f. hominis: Evidence for reinfection. J. Eukaryot. Microbiol. 1996, 43, 50S. [Google Scholar]
- Hauser, P.M.; Francioli, P.; Bille, J.; Telenti, A.; Blanc, D.S. Typing of Pneumocystis carinii f. sp. hominis by single-strand conformation polymorphism of four genomic regions. J. Clin. Microbiol. 1997, 35, 3086–3091. [Google Scholar]
- Keely, S.P.; Stringer, J.R. Sequences of Pneumocystis carinii f. sp. hominis strains associated with recurrent pneumonia vary at multiple loci. J. Clin. Microbiol. 1997, 35, 2745–2747. [Google Scholar]
- Olsson, M.; Lidman, C.; Latouche, S.; Björkman, A.; Roux, P.; Linder, E.; Wahlgren, M. Identification of Pneumocystis carinii f. sp. hominis gene sequences in filtered air in hospital environments. J. Clin. Microbiol. 1998, 36, 1737–1740. [Google Scholar] [PubMed]
- Tsolaki, S.; Ortona, E.; Mazars, E.; Margutti, P.; Tamburrini, E.; Siracusano, A.; Guyot, K.; Nigou, M.; Roux, P. Biodiversity of Pneumocystis carinii hominis: Typing with different DNA regions. J. Clin. Microbiol. 1997, 35, 383–387. [Google Scholar]
- Tsolaki, A.G.; Beckers, P.; Wakefield, A.E. Pre-AIDS era isolates of Pneumocystis carinii f. sp. hominis: High genotypic similarity with contemporary isolates. J. Clin. Microbiol. 1998, 36, 90–93. [Google Scholar] [PubMed]
- Lee, C.-H.; Helweg-Larsen, J.; Tang, X.; Jin, S.; Li, B.; Bartlett, M.S.; Lu, J.-J.; Lundgren, B.; Lundgren, J.D.; Olsson, M. Update on Pneumocystis carinii f. sp. hominis typing based on nucleotide sequence variations in internal transcribed spacer regions of rRNA genes. J. Clin. Microbiol. 1998, 36, 734–741. [Google Scholar] [PubMed]
- Santos, L.D.; Lacube, P.; Latouche, S.; Kac, G.; Mayaud, C.; Marteau, M.; Poirot, J.L.; Maury, E.; Guillot, J.; Roux, P. Contribution of Dihydropteroate synthase gene typing for Pneumocystis carinii f. sp. hominis epidemiology. J. Eukaryot. Microbiol. 1999, 46, 133S. [Google Scholar]
- Ma, L.; Borio, L.; Masur, H.; Kovacs, J.A. Pneumocystis carinii dihydropteroate synthase but not dihydrofolate reductase gene mutations correlate with prior trimethoprim-sulfamethoxazole or dapsone use. J. Infect. Dis. 1999, 180, 1969–1978. [Google Scholar] [CrossRef] [Green Version]
- Beard, C.B.; Carter, J.L.; Keely, S.P.; Huang, L.; Pieniazek, N.J.; Moura, I.N.; Roberts, J.M.; Hightower, A.W.; Bens, M.S.; Freeman, A.R. Genetic variation in Pneumocystis carinii isolates from different geographic regions: Implications for transmission. Emerg. Infect. Dis. 2000, 6, 265–272. [Google Scholar] [CrossRef]
- Hosoya, N.; Takahashi, T.; Wada, M.; Endo, T.; Nakamura, T.; Sakashita, H.; Kimura, K.; Ohnishi, K.; Nakamura, Y.; Mizuochi, T. Genotyping of Pneumocystis carinii f. sp. hominis isolates in Japan based on nucleotide sequence variations in internal transcribed spacer regions of rRNA genes. Microbiol. Immunol. 2000, 44, 591–596. [Google Scholar]
- Hauser, P.M.; Blanc, D.S.; Sudre, P.; Manoloff, E.S.; Nahimana, A.; Bille, J.; Weber, R.; Francioli, P. Genetic diversity of Pneumocystis carinii in HIV-positive and-negative patients as revealed by PCR–SSCP typing. Aids 2001, 15, 461–466. [Google Scholar] [CrossRef]
- Hauser, P.M.; Sudre, P.; Nahimana, A.; Francioli, P.; Group, S. Prophylaxis failure is associated with a specific Pneumocystis carinii genotype. Clin. Infect. Dis. 2001, 33, 1080–1082. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Kovacs, J.A. Genetic analysis of multiple loci suggests that mutations in the Pneumocystis carinii f. sp. hominis dihydropteroate synthase gene arose independently in multiple strains. Antimicrob. Agents Chemother. 2001, 45, 3213–3215. [Google Scholar] [CrossRef] [Green Version]
- Volpe, G.; Sbaiz, L.; Avanzini, C.; Caramello, P.; Savoia, D. Genetic diversity of Pneumocystis carinii isolated from human immunodeficiency virus-positive patients in Turin, Italy. J. Clin. Microbiol. 2001, 39, 2995–2998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meshnick, S.R.; Hossler, P.A.; Enger, K.S.; Kazanjian, P.; Rest, J.S.; Mindell, D.; Li, B.; Lee, C.; Nimri, L.F.; Carter, J.L. Distribution of DHPS mutations among ITS subtypes of P. carinii f. sp. hominis. J. Eukaryot. Microbiol. 2001. [Google Scholar] [CrossRef]
- Takahashi, T.; Kanda, T.; Iwamoto, A. Genetic diversity of drug targets including Dihydropteroate synthase, Dihydrofolate reductase and Cytochrome b, in Pneumocystis carinii f. sp. hominis isolates in Japan. Res. Commun. Mol. Pathol. Pharmacol. 2002, 112, 159–176. [Google Scholar] [PubMed]
- Miller, R.F.; Ambrose, H.E.; Novelli, V.; Wakefield, A.E. Probable mother-to-infant transmission of Pneumocystis carinii f. sp. hominis infection. J. Clin. Microbiol. 2002, 40, 1555–1557. [Google Scholar]
- Montes-Cano, M.A.; De la Horra, C.; Martin-Juan, J.; Varela, J.M.; Torronteras, R.; Respaldiza, N.; Medrano, F.J.; Calderón, E.J. Pneumocystis jiroveci genotypes in the Spanish population. Clin. Infect. Dis. 2004, 39, 123–128. [Google Scholar] [CrossRef]
- Matos, O.; Lee, C.-H.; Jin, S.; Li, B.; Costa, M.C.; Gonçalves, L.; Antunes, F. Pneumocystis jirovecii in Portuguese immunocompromised patients: Association of specific ITS genotypes with treatment failure, bad clinical outcome and childhood. Infect. Genet. Evol. 2003, 3, 281–285. [Google Scholar] [CrossRef]
- Miller, R.F.; Lindley, A.R.; Ambrose, H.E.; Malin, A.S.; Wakefield, A.E. Genotypes of Pneumocystis jirovecii isolates obtained in Harare, Zimbabwe, and London, United Kingdom. Antimicrob. Agents Chemother. 2003, 47, 3979–3981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakefield, A.E.; Lindley, A.R.; Ambrose, H.E.; Denis, C.-M.; Miller, R.F. Limited asymptomatic carriage of Pneumocystis jirovecii in human immunodeficiency virus–infected patients. J. Infect. Dis. 2003, 187, 901–908. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.F.; Lindley, A.R.; Ambrose, H.E.; Aliouat-Denis, C.M.; Wakefield, A.E. Multilocus Genotyping of Pneumocystis jirovecii from Adult HIV-Infected Patients with Pneumocystis Pneumonia. J. Eukaryot. Microbiol. 2003, 50, 654–655. [Google Scholar] [CrossRef] [PubMed]
- Robberts, F.J.L.; Liebowitz, L.D.; Chalkley, L.J. Genotyping and coalescent phylogenetic analysis of Pneumocystis jirovecii from South Africa. J. Clin. Microbiol. 2004, 42, 1505–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahimana, A.; Rabodonirina, M.; Helweg-Larsen, J.; Meneau, I.; Francioli, P.; Bille, J.; Hauser, P.M. Sulfa resistance and Dihydropteroate synthase mutants in recurrent Pneumocystis carinii pneumonia. Emerg. Infect. Dis. 2003, 9, 864–867. [Google Scholar] [CrossRef] [PubMed]
- Höcker, B.; Wendt, C.; Nahimana, A.; Tönshoff, B.; Hauser, P.M. Molecular evidence of Pneumocystis transmission in pediatric transplant unit. Emerg. Infect. Dis. 2005, 11, 330–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medrano, F.J.; Montes-Cano, M.; Conde, M.; De La Horra, C.; Respaldiza, N.; Gasch, A.; Perez-Lozano, M.J.; Varela, J.M.; Calderon, E.J. Pneumocystis jirovecii in general population. Emerg. Infect. Dis. 2005, 11, 245–250. [Google Scholar] [CrossRef]
- Beard, C.B.; Fox, M.R.; Lawrence, G.G.; Guarner, J.; Hanzlick, R.L.; Huang, L.; del Rio, C.; Rimland, D.; Duchin, J.S.; Colley, D.G. Genetic differences in Pneumocystis isolates recovered from immunocompetent infants and from adults with AIDS: Epidemiological implications. J. Infect. Dis. 2005, 192, 1815–1818. [Google Scholar] [CrossRef] [Green Version]
- Valerio, A.; Tronconi, E.; Mazza, F.; Cargnel, A.; Fantoni, G.; Atzori, C. DHPS -Mutated Isolates of Pneumocystis jirovecii from HIV-Infected Individuals: Analysis of Related ITS Genotypes. J. Eukaryot. Microbiol. 2006, 53, S108–S109. [Google Scholar] [CrossRef]
- Costa, M.C.; Esteves, F.; Antunes, F.; Matos, O. Multilocus genotyping of Pneumocystis jirovecii in immunocompromised patients: Preliminary results. J. Eukaryot. Microbiol. 2006, 53, S104–S105. [Google Scholar] [CrossRef]
- Gupta, R.; Mirdha, B.R.; Guleria, R.; Mohan, A.; Kabra, S.K.; Kumar, L.; Agarwal, S.K.; Luthra, K. Use of different primer directed sequence amplification by polymerase chain reaction for identification of Pneumocystis jirovecii in clinical samples. Indian J. Chest Dis. Allied Sci. 2008, 50, 321–327. [Google Scholar]
- Esteves, F.; Montes-Cano, M.A.; De La Horra, C.; Costa, M.C.; Calderón, E.J.; Antunes, F.; Matos, O. Pneumocystis jirovecii multilocus genotyping profiles in patients from Portugal and Spain. Clin. Microbiol. Infect. 2008, 14, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Siripattanapipong, S.; Leelayoova, S.; Mungthin, M.; Worapong, J.; Tan-Ariya, P. Study of DHPS and DHFR genes of Pneumocystis jirovecii in Thai HIV-infected patients. Med. Mycol. 2008, 46, 389–392. [Google Scholar] [CrossRef]
- Esteves, F.; Gaspar, J.; Tavares, A.; Moser, I.; Antunes, F.; Mansinho, K.; Matos, O. Population structure of Pneumocystis jirovecii isolated from immunodeficiency virus-positive patients. Infect. Genet. Evol. 2010, 10, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Schmoldt, S.; Schuhegger, R.; Wendler, T.; Huber, I.; Söllner, H.; Hogardt, M.; Arbogast, H.; Heesemann, J.; Bader, L.; Sing, A. Molecular evidence of nosocomial Pneumocystis jirovecii transmission among 16 patients after kidney transplantation. J. Clin. Microbiol. 2008, 46, 966–971. [Google Scholar] [CrossRef] [Green Version]
- Gianella, S.; Haeberli, L.; Joos, B.; Ledergerber, B.; Wuethrich, R.-P.; Weber, R.; Kuster, H.; Hauser, P.-M.; Fehr, T.; Mueller, N.J. Molecular evidence of interhuman transmission in an outbreak of Pneumocystis jirovecii pneumonia in renal transplant recipients. Transpl. Infect. Dis. 2010, 12, 1–10. [Google Scholar] [CrossRef]
- van Hal, S.J.; Gilgado, F.; Doyle, T.; Barratt, J.; Stark, D.; Meyer, W.; Harkness, J. Clinical significance and phylogenetic relationship of novel Australian Pneumocystis jirovecii genotypes. J. Clin. Microbiol. 2009, 47, 1818–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montes-Cano, M.A.; Chabe, M.; Fontillon-Alberdi, M.; de la Horra, C.; Respaldiza, N.; Medrano, F.J.; Varela, J.M.; Dei-Cas, E.; Calderon, E.J. Vertical transmission of Pneumocystis jirovecii in humans. Emerg. Infect. Dis. 2009, 15, 125–127. [Google Scholar] [CrossRef]
- Esteves, F.; Tavares, A.; Costa, M.C.; Gaspar, J.; Antunes, F.; Matos, O. Genetic characterization of the UCS and Kex1 loci of Pneumocystis jirovecii. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Esteves, F.; Gaspar, J.; Marques, T.; Leite, R.; Antunes, F.; Mansinho, K.; Matos, O. Identification of relevant single-nucleotide polymorphisms in Pneumocystis jirovecii: Relationship with clinical data. Clin. Microbiol. Infect. 2010, 16, 878–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.; Vivancos, R.; Corless, C.; Wood, G.; Beeching, N.J.; Beadsworth, M.B. Increasing frequency of Pneumocystis jirovecii pneumonia in renal transplant recipients in the United Kingdom: Clonal variability, clusters, and geographic location. Clin. Infect. Dis. 2011, 53, 307–308. [Google Scholar] [CrossRef] [Green Version]
- Beser, J.; Botero-Kleiven, S.; Lebbad, M.; Hagblom, P.; Fernandez, V. A limited number of ITS haplotypes defines the diversity of Pneumocystis jirovecii strains in Sweden. Infect. Genet. Evol. 2011, 11, 948–954. [Google Scholar] [CrossRef]
- Pliquett, R.U.; Asbe-Vollkopf, A.; Hauser, P.M.; Presti, L.L.; Hunfeld, K.P.; Berger, A.; Scheuermann, E.H.; Jung, O.; Geiger, H.; Hauser, I.A. A Pneumocystis jirovecii pneumonia outbreak in a single kidney-transplant center: Role of cytomegalovirus co-infection. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 2429–2437. [Google Scholar] [CrossRef]
- Dimonte, S.; Berrilli, F.; D’Orazi, C.; D’Alfonso, R.; Placco, F.; Bordi, E.; Perno, C.F.; Di Cave, D. Molecular analysis based on mtLSU-rRNA and DHPS sequences of Pneumocystis jirovecii from immunocompromised and immunocompetent patients in Italy. Infect. Genet. Evol. 2013, 14, 68–72. [Google Scholar] [CrossRef] [PubMed]
- de Armas, Y.; Friaza, V.; Capó, V.; Durand-Joly, I.; Govín, A.; de La Horra, C.; Dei-Cas, E.; Calderón, E.J. Low genetic diversity of Pneumocystis jirovecii among Cuban population based on two-locus mitochondrial typing. Med. Mycol. 2012, 50, 417–420. [Google Scholar] [CrossRef]
- Curran, T.; McCaughey, C.; Coyle, P. V Pneumocystis jirovecii multilocus genotyping profiles in Northern Ireland. J. Med. Microbiol. 2013, 62, 1170–1174. [Google Scholar] [CrossRef] [Green Version]
- Debourgogne, A.; Favreau, S.; Ladriere, M.; Bourry, S.; Machouart, M. Characteristics of Pneumocystis pneumonia in Nancy from January 2007 to April 2011 and focus on an outbreak in nephrology. J. Mycol. Med. 2014, 24, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Nankivell, B.J.; Firacative, C.; Kable, K.; Chen, S.C.-A.; Meyer, W. Molecular epidemiology linking multihospital clusters of opportunistic Pneumocystis jirovecii pneumonia. Clin. Infect. Dis. 2013, 57, 1058–1059. [Google Scholar] [CrossRef] [PubMed]
- Rostved, A.A.; Sassi, M.; Kurtzhals, J.A.L.; Sørensen, S.S.; Rasmussen, A.; Ross, C.; Gogineni, E.; Huber, C.; Kutty, G.; Kovacs, J.A. Outbreak of Pneumocystis Pneumonia in renal and liver transplant patients caused by genotypically distinct strains of Pneumocystis jirovecii. Transplantation 2013, 96, 834–842. [Google Scholar] [CrossRef] [Green Version]
- Chua, K.Y.L.; Halliday, C.L.; Grote, D.; Meyer, W.; Kesson, A.M.; Chen, S.C.A. Colonisation with Pneumocystis jirovecii in Australian infants. Pathol. RCPA 2015, 47, 489–490. [Google Scholar] [CrossRef] [PubMed]
- Alanio, A.; Morio, F.; Gits-Muselli, M.; Desnos-Ollivier, M.; Maitte, C.; Bretagne, S. Short tandem repeat genotyping for P. jirovecii is more sensitive to mixed genotype than MLST. J. Mycol. Med. 2015, 25, 220–221. [Google Scholar] [CrossRef]
- Depypere, M.; Saegeman, V.; Lagrou, K. Typing of Pneumocystis jirovecii by multilocus sequencing: Evidence of outbreak? Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 911–916. [Google Scholar] [CrossRef]
- Desoubeaux, G.; Dominique, M.; Morio, F.; Thepault, R.-A.; Franck-Martel, C.; Tellier, A.-C.; Ferrandière, M.; Hennequin, C.; Bernard, L.; Salamé, E. Epidemiological outbreaks of Pneumocystis jirovecii pneumonia are not limited to kidney transplant recipients: Genotyping confirms common source of transmission in a liver transplantation unit. J. Clin. Microbiol. 2016, 54, 1314–1320. [Google Scholar] [CrossRef] [Green Version]
- Charpentier, E.; Garnaud, C.; Wintenberger, C.; Bailly, S.; Murat, J.-B.; Rendu, J.; Pavese, P.; Drouet, T.; Augier, C.; Malvezzi, P. Added value of next-generation sequencing for multilocus sequence typing analysis of a Pneumocystis jirovecii pneumonia outbreak. Emerg. Infect. Dis. 2017, 23, 1237–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vindrios, W.; Argy, N.; Le Gal, S.; Lescure, F.-X.; Massias, L.; Le, M.P.; Wolff, M.; Yazdanpanah, Y.; Nevez, G.; Houze, S. Outbreak of Pneumocystis jirovecii infection among heart transplant recipients: Molecular investigation and management of an interhuman transmission. Clin. Infect. Dis. 2017, 65, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Sokulska, M.; Kicia, M.; Wesołowska, M.; Piesiak, P.; Kowal, A.; Lobo, M.L.; Kopacz, Ż.; Hendrich, A.B.; Matos, O. Genotyping of Pneumocystis jirovecii in colonized patients with various pulmonary diseases. Med. Mycol. 2018, 56, 809–815. [Google Scholar] [CrossRef]
- Ricci, G.; Santos, D.W.; Kovacs, J.A.; Nishikaku, A.S.; de Sandes-Freitas, T.V.; Rodrigues, A.M.; Kutty, G.; Affonso, R.; Silva, H.T.; Medina-Pestana, J.O. Genetic diversity of Pneumocystis jirovecii from a cluster of cases of pneumonia in renal transplant patients: Cross-sectional study. Mycoses 2018, 61, 845–852. [Google Scholar] [CrossRef]
- Singh, Y.; Mirdha, B.R.; Guleria, R.; Kabra, S.K.; Mohan, A.; Chaudhry, R.; Kumar, L.; Dwivedi, S.N.; Agarwal, S.K. Genetic polymorphisms associated with treatment failure and mortality in pediatric Pneumocystosis. Sci. Rep. 2019, 9, 1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevez, G.; Guillaud-Saumur, T.; Cros, P.; Papon, N.; Vallet, S.; Quinio, D.; Minoui-Tran, A.; Pilorgé, L.; de Parscau, L.; Sizun, J. Pneumocystis primary infection in infancy: Additional French data and review of the literature. Med. Mycol. 2020, 58, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Sürgeç, E.; Can, H.; Döşkaya, M.; Karakavuk, M.; Atalay Şahar, E.; Değirmenci Döşkaya, A.; Pullukçu, H.; Taşbakan, M.; Sezai Taşbakan, M.; Akyol, D. Genotyping of Pneumocystis jirovecii isolates obtained from clinical samples by multilocus sequencing: A molecular epidemiology study conducted in Turkey. Arch. Microbiol. 2020, 202, 1647–1652. [Google Scholar] [CrossRef]
- Le Gal, S.; Toubas, D.; Totet, A.; Dalle, F.; Abou Bacar, A.; Le Meur, Y.; Nevez, G. Pneumocystis infection outbreaks in organ transplantation units in France: A Nation-wide survey. Clin. Infect. Dis. 2020, 70, 2216–2220. [Google Scholar] [CrossRef]
- Szydłowicz, M.; Jakuszko, K.; Szymczak, A.; Piesiak, P.; Kowal, A.; Kopacz, Ż.; Wesołowska, M.; Lobo, M.L.; Matos, O.; Hendrich, A.B. Prevalence and genotyping of Pneumocystis jirovecii in renal transplant recipients—Preliminary report. Parasitol. Res. 2019, 118, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, C.; Zuluaga, A.; Restrepo, A.; Tobón, A.; Cano, L.E.; Gonzalez, A. Molecular diagnosis and detection of Pneumocystis jirovecii DHPS and DHFR genotypes in respiratory specimens from Colombian patients. Diagn. Microbiol. Infect. Dis. 2012, 72, 204–213. [Google Scholar] [CrossRef]
- Taylor, S.M.; Meshnick, S.R.; Worodria, W.; Andama, A.; Cattamanchi, A.; Davis, J.L.; Yoo, S.D.; Byanyima, P.; Kaswabuli, S.; Goodman, C.D. Low prevalence of Pneumocystis pneumonia (PCP) but high prevalence of Pneumocystis dihydropteroate synthase (dhps) gene mutations in HIV-infected persons in Uganda. PLoS ONE 2012, 7, e49991. [Google Scholar] [CrossRef]
- Liu, Y.; Fahle, G.A.; Kovacs, J.A. Inability to culture Pneumocystis jirovecii. MBio 2018, 9, e00939-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, P.; Lavrard, I.; Poirot, J.L.; Chouaid, C.; Denis, M.; Olivier, J.L.; Nigou, M.; Miltgen, M. Usefulness of PCR for detection of Pneumocystis carinii DNA. J. Clin. Microbiol. 1994, 32, 2324–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, T.; Ma, Z.; Liu, F.; Du, W.-Q.; He, L.; Ma, L.; An, C.-L. Genotyping of Pneumocystis jirovecii by use of a new simplified nomenclature system based on the internal transcribed spacer regions and 5.8S rRNA gene of the rRNA operon. J. Clin. Microbiol. 2019, 57, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Valero, C.; Buitrago, M.J.; Gits-Muselli, M.; Benazra, M.; Sturny-Leclère, A.; Hamane, S.; Guigue, N.; Bretagne, S.; Alanio, A. Copy number variation of mitochondrial DNA genes in Pneumocystis jirovecii according to the fungal load in BAL specimens. Front. Microbiol. 2016, 7, 1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanio, A.; Gits-Muselli, M.; Mercier-Delarue, S.; Dromer, F.; Bretagne, S. Diversity of Pneumocystis jirovecii during infection revealed by ultra-deep pyrosequencing. Front. Microbiol. 2016, 7, 733. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Cissé, O.H.; Kovacs, J.A. A molecular window into the biology and epidemiology of Pneumocystis spp. Clin. Microbiol. Rev. 2018, 31, 1–49. [Google Scholar] [CrossRef] [Green Version]
- Nahimana, A.; Francioli, P.; Blanc, D.S.; Bille, J.; Wakefield, A.E.; Hauser, P.M. Determination of the copy number of the nuclear rDNA and Beta-tubulin genes of Pneumocystis carinii f. sp. hominis using PCR multicompetitors. J. Eukaryot. Microbiol. 2000, 47, 368–372. [Google Scholar] [CrossRef]
- Brancart, F.; Rodriguez-Villalobos, H.; Fonteyne, P.-A.; Peres-Bota, D.; Liesnard, C. Quantitative TaqMan PCR for detection of Pneumocystis jiroveci. J. Microbiol. Methods 2005, 61, 381–387. [Google Scholar] [CrossRef]
- Morilla, R.; González-Magaña, A.; Friaza, V.; de Armas, Y.; Medrano, F.J.; Calderón, E.; de la Horra, C. Genetic polymorphisms of Superoxide Dismutase locus of Pneumocystis jirovecii in Spanish population. Front. Public Health 2019, 7, 292. [Google Scholar] [CrossRef]
- Huang, L.; Crothers, K.; Atzori, C.; Benfield, T.; Miller, R.; Rabodonirina, M.; Helweg-Larsen, J. Dihydropteroate synthase gene mutations in Pneumocystis and sulfa resistance. Emerg. Infect. Dis. 2004, 10, 1721–1728. [Google Scholar] [CrossRef]
- Nahimana, A.; Rabodonirina, M.; Bille, J.; Francioli, P.; Hauser, P.M. Mutations of Pneumocystis jirovecii Dihydrofolate reductase associated with failure of prophylaxis. Antimicrob. Agents Chemother. 2004, 48, 4301–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matos, O.; Esteves, F. Pneumocystis jirovecii multilocus gene sequencing: Findings and implications. Future Microbiol. 2010, 5, 1257–1267. [Google Scholar] [CrossRef]
- Costa, M.C.; Esteves, F.; Antunes, F.; Matos, O. Genetic characterization of the Dihydrofolate reductase gene of Pneumocystis jirovecii isolates from Portugal. J. Antimicrob. Chemother. 2006, 58, 1246–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunkin, S.M.; Stringer, J.R. Translocation of surface antigen genes to a unique telomeric expression site in Pneumocystis carinii. Mol. Microbiol. 1996, 19, 283–295. [Google Scholar] [CrossRef]
- Procop, G.W.; Haddad, S.; Quinn, J.; Wilson, M.L.; Henshaw, N.G.; Reller, L.B.; Artymyshyn, R.L.; Katanik, M.T.; Weinstein, M.P. Detection of Pneumocystis jiroveci in respiratory specimens by four staining methods. J. Clin. Microbiol. 2004, 42, 3333–3335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakefield, A.E.; Pixley, F.J.; Banerji, S.; Sinclair, K.; Miller, R.F.; Moxon, E.R.; Hopkin, J.M. Amplification of mitochondrial ribosomal RNA sequences from Pneumocystis carinii DNA of rat and human origin. Mol. Biochem. 1990, 43, 69–76. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Fisher, M.E.; Scesney, P.E. Visibility of Critical-Exponent Renormalization. Phys. Rev. A 1970, 3, 825. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0. Mol. Biol. Evol. 2015, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- De Boer, M.G.J.; de Fijter, J.W.; Kroon, F.P. Outbreaks and clustering of Pneumocystis pneumonia in kidney transplant recipients: A systematic review. Med. Mycol. 2011, 49, 673–680. [Google Scholar] [PubMed] [Green Version]
- Kim, J.E.; Han, A.; Lee, H.; Ha, J.; Kim, Y.S.; Han, S.S. Impact of Pneumocystis jirovecii pneumonia on kidney transplant outcome. BMC Nephrol. 2019, 20, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanio, A.; Hauser, P.M.; Lagrou, K.; Melchers, W.J.G.; Helweg-Larsen, J.; Matos, O.; Cesaro, S.; Maschmeyer, G.; Einsele, H.; Donnelly, J.P. ECIL guidelines for the diagnosis of Pneumocystis jirovecii pneumonia in patients with haematological malignancies and stem cell transplant recipients. J. Antimicrob. Chemother. 2016, 71, 2386–2396. [Google Scholar] [CrossRef] [PubMed]
- Alanio, A.; Desoubeaux, G.; Sarfati, C.; Hamane, S.; Bergeron, A.; Azoulay, E.; Molina, J.M.; Derouin, F.; Menotti, J. Real-time PCR assay-based strategy for differentiation between active Pneumocystis jirovecii pneumonia and colonization in immunocompromised patients. Clin. Microbiol. Infect. 2011, 17, 1531–1537. [Google Scholar] [CrossRef]
- Lu, Y.; Ling, G.; Qiang, C.; Ming, Q.; Wu, C.; Wang, K.; Ying, Z. PCR diagnosis of Pneumocystis pneumonia: A bivariate meta-analysis. J. Clin. Microbiol. 2011, 49, 4361–4363. [Google Scholar] [CrossRef] [Green Version]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [Green Version]
- Le Gal, S.; Damiani, C.; Rouillé, A.; Grall, A.; Tréguer, L.; Virmaux, M.; Moalic, E.; Quinio, D.; Moal, M.-C.; Berthou, C. A cluster of Pneumocystis infections among renal transplant recipients: Molecular evidence of colonized patients as potential infectious sources of Pneumocystis jirovecii. Clin. Infect. Dis. 2012, 54, e62–e71. [Google Scholar] [CrossRef]
- Helweg-Larsen, J.; Benfield, T.L.; Eugen-Olsen, J.; Lundgren, J.D.; Lundgren, B. Effects of mutations in Pneumocystis carinii Dihydropteroate synthase gene on outcome of AIDS-associated P. carinii pneumonia. Lancet 1999, 354, 1347–1351. [Google Scholar] [CrossRef]
- Suárez, I.; Roderus, L.; van Gumpel, E.; Jung, N.; Lehmann, C.; Fätkenheuer, G.; Hartmann, P.; Plum, G.; Rybniker, J. Low prevalence of DHFR and DHPS mutations in Pneumocystis jirovecii strains obtained from a German cohort. Infection 2017, 45, 341–347. [Google Scholar] [CrossRef]
- Iliades, P.; Meshnick, S.R.; Macreadie, I.G. Mutations in the Pneumocystis jirovecii DHPS gene confer cross-resistance to sulfa drugs. Antimicrob. Agents Chemother. 2005, 49, 741–748. [Google Scholar] [CrossRef] [Green Version]
- Ponce, C.A.; Chabé, M.; George, C.; Cárdenas, A.; Durán, L.; Guerrero, J.; Bustamante, R.; Matos, O.; Huang, L.; Miller, R.F. High prevalence of Pneumocystis jirovecii Dihydropteroate synthase gene mutations in patients with a first episode of Pneumocystis pneumonia in Santiago, Chile, and clinical response to trimethoprim-sulfamethoxazole therapy. Antimicrob. Agents Chemother. 2017, 61, e01290-16. [Google Scholar] [CrossRef] [Green Version]
- Rabodonirina, M.; Nahimana, A.; Weber, R.; Francioli, P.; Bille, J.; Hauser, P.M. Geographical variation in the prevalence of Pneumocystis jirovecii Dihydropteroate synthase mutations within Western Europe. J. Eukaryot. Microbiol. 2006, 53, S112–S113. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Schemes (Included Loci) and Reference | Country | Total # of Samples | # of Samples Sequenced | # of Sequence Types | Genetic Locus | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5.8S | 18S | 23S | 26S | ITS1 | ITS2 | ITS1/2 | msg | mt26S | β-TUB | TS | arom/EPSP | mtSSU | DHPS | UCS | Kex1 | CYB | SOD | DHRF | TRR1 | |||||
| Scheme 1 (5.8S, 18S, 26S, ITS1, ITS2) | ||||||||||||||||||||||||
| [6] | USA | 15 | 15 | 6 | 1 | 1 | 1 | 2 | 3 | NG | ||||||||||||||
| [7] | GBR | 24 | 24 | NG | 1 | 1 | 1 | 5 | 7 | NG | ||||||||||||||
| Scheme 2 (ITS1, ITS2, msg, mt26S) | ||||||||||||||||||||||||
| [8] | USA | 15 | 15 | NG | NG | NG | NG | NG | ||||||||||||||||
| [38] | IND | 180 | 29 | NG | NG | NG | NG | NG | ||||||||||||||||
| Scheme 3 (26S, β-TUB, ITS1, mt26S) | ||||||||||||||||||||||||
| [9] | CHE | 11 | 11 | NG | 3 | 3 | 3 | 2 | ||||||||||||||||
| [20] | EUR | 212 | 212 | 6 | 6 | 3 | 4 | 3 | ||||||||||||||||
| [19] | EUR | 91 | 91 | 28 | NG | NG | NG | NG | ||||||||||||||||
| [33] | DEU | 7 | 7 | 2 | NG | NG | NG | NG | ||||||||||||||||
| [42] | DEU | 20 | 14 | NG | 2 | 4 | 4 | 1 | ||||||||||||||||
| [43] | CHE | 19 | 7 | 1(+) | 2 | 4 | 3 | 2 | ||||||||||||||||
| [50] | DE | 18 | 18 | NG | 2 | 3 | 3 | 2 | ||||||||||||||||
| [53] | GBR | 670 | 31 | NG | NA | 5 | 4 | NA | ||||||||||||||||
| [54] | FRA | 13 | 10 | 3 | 2 | 2 | 3 | 1 | ||||||||||||||||
| Scheme 4 (ITS1, ITS2, mt26S) | ||||||||||||||||||||||||
| [10] | USA | 15 | 15 | 6 | 4 | 3 | 7 | 4 | ||||||||||||||||
| [11] | SWE, FRA | 7 | 7 | NG | 4 | 4 | NG | 3 | ||||||||||||||||
| [48] | GBR | 27 | 27 | NG | NG | NG | 2 | 3 | ||||||||||||||||
| Scheme 5 (5.8S, ITS1, ITS2, mt26S, TS) | ||||||||||||||||||||||||
| [12] | FRA, ITA | 20 | 18 | NG | 6 | 3 | 3 | 10 | 4 | 1 | ||||||||||||||
| Scheme 6 (arom, ITS1, ITS2, mt26S, mtSSU) | ||||||||||||||||||||||||
| [13] | NLD | 6 | 6 | NG | NG | NG | 9 | 3 | 1 | 2 | ||||||||||||||
| Scheme 7 (ITS1, ITS2) | ||||||||||||||||||||||||
| [14] | GLO | 207 | 207 | NG | 15 | 14 | NG | |||||||||||||||||
| [18] | JPN | 24 | 24 | NG | 11 | 11 | NG | |||||||||||||||||
| [31] | ZAF | 20 | 20 | NG | 11 | 13 | NG | |||||||||||||||||
| [49] | SWE | 64 | 64 | 12 | 10 | 12 | NG | |||||||||||||||||
| Scheme 8 (DHPS, ITS1, ITS2, mt26S) | ||||||||||||||||||||||||
| [15] | FRA | 14 | 14 | NG | NG | NG | NG | NG | NG | |||||||||||||||
| [39] | PRT, ESP | 108 | ✕ | NG | 12 | 10 | NG | 4 | 4 | |||||||||||||||
| [44] | AUS | 68 | 68 | NG | 8 | 9 | 16 | 2 | 2 | |||||||||||||||
| Scheme 9 (DHFR, DHPS) | ||||||||||||||||||||||||
| [16] | USA | 37 | 37 | NG | 4 | 2 | ||||||||||||||||||
| [32] | FRA | 33 | 33 | NG | 3 | 2 | ||||||||||||||||||
| [40] | THA | 29 | 18 | NG | 3 | 3 | ||||||||||||||||||
| [71] | USA | 13 | 13 | NG | 2 | 2 | ||||||||||||||||||
| [70] | COL | 98 | 45 | NG | 4 | 2 | ||||||||||||||||||
| Scheme 10 (DHPS, mt26S) | ||||||||||||||||||||||||
| [17] | USA | 324 | 191 | 14 | 4 | 4 | ||||||||||||||||||
| [26] | ESP | 255 | 79 | NG | 4 | 4 | ||||||||||||||||||
| [34] | ESP | 50 | 12 | NG | 4 | 1 | ||||||||||||||||||
| [35] | USA | 442 | ❋ | NG | 4 | 4 | ||||||||||||||||||
| [45] | ESP | 60 | 19 | NG | 3 | 1 | ||||||||||||||||||
| [51] | ITA | 67 | ⚑ | NG | 4 | 3 | ||||||||||||||||||
| Scheme 11 (26S, β-TUB, DHPS, ITS1, mt26S) | ||||||||||||||||||||||||
| [21] | USA | 22 | 22 | 10 | 2 | 5 | 4 | 2 | 3 | |||||||||||||||
| Scheme 12 (26S, β-TUB, ITS1, ITS2, mt26S) | ||||||||||||||||||||||||
| [22] | ITA | 25 | 18 | 15 | 4 | 6 | 6 | 4 | 3 | |||||||||||||||
| Scheme 13 (DHPS, ITS1, ITS2) | ||||||||||||||||||||||||
| [23] | USA | 57 | 37 | NG | 6 | 7 | NG | 3 | ||||||||||||||||
| [27] | PRT | 43 | 43 | NG | 15 | 14 | 17 | 2 | ||||||||||||||||
| [36] | ITA | 261 | 174 | NG | NG | NG | 9 | 3 | ||||||||||||||||
| Scheme 14 (CYB, DHFR, DHPS) | ||||||||||||||||||||||||
| [24] | JPN | 34 | 34 | NG | 4 | 2 | 1 | |||||||||||||||||
| Scheme 15 (DHPS, ITS1, ITS2, mtSSU) | ||||||||||||||||||||||||
| [25] | GBR | 2 | 2 | NG | NG | NG | 2 | 1 | 1 | |||||||||||||||
| Scheme 16 (DHPS, mt26S, mtSSU) | ||||||||||||||||||||||||
| [28] | GBR, ZWE | 51 | 30 | NG | 3 | 4 | 2 | |||||||||||||||||
| Scheme 17 (DHPS, mtSSU, mt26S, SOD) | ||||||||||||||||||||||||
| [29] | GBR | 16 | 16 | NG | NG | NG | NG | NG | ||||||||||||||||
| [30] | NG | 76 | 76 | 15 | 4 | 4 | 4 | 4 | ||||||||||||||||
| Scheme 18 (DHFR, DHPS, ITS1, ITS2) | ||||||||||||||||||||||||
| [37] | PRT | 68 | 68 | NG | NG | NG | 19 | 4 | 4 | |||||||||||||||
| Scheme 19 (UCS, Kex1) | ||||||||||||||||||||||||
| [39] | PRT | 87 | 35 | NG | 4 | 4 | ||||||||||||||||||
| Scheme 20 (CYB, DHFR, DHPS, mt26S, SOD) | ||||||||||||||||||||||||
| [46] | PRT | 102 | 78 | NG | 9 | 6 | 3 | 6 | 6 | |||||||||||||||
| Scheme 21 (β-TUB, CYB, DHFR, DHPS, mt26S, TRR1, TS, SOD) | ||||||||||||||||||||||||
| [47] | PRT | 70 | ► | 48 | 5 | 3 | ⌘ | 3 | 7 | 4 | 3 | ⌘ | ||||||||||||
| Scheme 22 (β-TUB, DHPS, ITS1/2, mt26S) | ||||||||||||||||||||||||
| [3] | AUS | 11 | 11 | 2 | 4 | 2 | 2 | 1 | ||||||||||||||||
| [55] | AUS | 48 | 48 | 4 | 2 | 2 | 1 | 1 | ||||||||||||||||
| [57] | AUS | 7 | 7 | NG | NG | NG | NG | NG | ||||||||||||||||
| Scheme 23 (mt26S, mtSSU) | ||||||||||||||||||||||||
| [52] | FRA, CUB, ESP | 75 | 75 | NG | 5 | 3 | ||||||||||||||||||
| Scheme 24 (26S, β-TUB, CYB, DHFR, DHPS, ITS1, mt26S, SOD) | ||||||||||||||||||||||||
| [5] | FRA | 23 | 23 | NG | 7 | 9 | 4 | 2 | 1 | 7 | 3 | 3 | ||||||||||||
| Scheme 25 (26S, ITS1, ITS2, mt26S) | ||||||||||||||||||||||||
| [56] | DNK | 22 | 18 | 3 | NG | NG | NG | NG | NG | |||||||||||||||
| Scheme 26 (CYB, ITS1, mt26S, SOD) | ||||||||||||||||||||||||
| [58] | FRA | 37 | 32 | NG | NG | NG | NG | NG | ||||||||||||||||
| Scheme 27 (β-TUB, CYB, DHFR, mt26S, SOD) | ||||||||||||||||||||||||
| [59] | BEL | 20 | 20 ^ | NG | NA | 4 | 2 | 4 | 2 | 2 | ||||||||||||||
| Scheme 28 (CYB, mt26S, SOD) | ||||||||||||||||||||||||
| [60] | FRA | 24 | ◎ | 14 | 6 | 5 | 3 | |||||||||||||||||
| [61] | FRA | 32 | 32 | 18 | 22 | 14 | 4 | |||||||||||||||||
| [62] | FRA | 7 | 7 | NG | 4 | 3 | 2 | |||||||||||||||||
| [63] | POL | 17 | ◉ | 8 | 13 | 6 | 2 | |||||||||||||||||
| [66] | FRA | 192 | 35 | 17 | 11 | 5 | 2 | |||||||||||||||||
| [67] | TUR | 31 | 26 | 6 | 4 | 5 | 3 | |||||||||||||||||
| [68] | REU, GUF, FRA | 47 | 47 | 23 | 5 | 9 | 3 | |||||||||||||||||
| Scheme 29 (23S, 26S, DHPS) | ||||||||||||||||||||||||
| [64] | BRA | 30 | 30 | 5 | 3 | 2 | 1 | |||||||||||||||||
| Scheme 30 (DHFR, DHPS, mt26S) | ||||||||||||||||||||||||
| [65] | IND | 37 | 37 | 13 | 3 | 3 | 2 | |||||||||||||||||
| Scheme31 (CYB, DHPS, mt26S, SOD) | ||||||||||||||||||||||||
| [69] | POL | 72 | N/A | N/A | 3 | 1 | 3 | 2 | ||||||||||||||||
| Locus | Primer Name | Ref. | Nucleotide Sequence | Product Size (Base Pairs) | PCR Conditions |
|---|---|---|---|---|---|
| β-TUB | PneumoβTub_F | - | 5′-TCATTAGGTGGTGGAACGGG-3′ | 303 | 95 °C 3 min; 45 cycles: 94 °C 30 s, 60 °C 45 s, 72 °C 45 s; 72 °C 7 min |
| PneumoβTub_R | 5′-ATCACCATATCCTGGATCCG-3′ | ||||
| SOD | MnSODFw | 5 | 5′-GGGTTTAATTAGTCTTTTTAGGCAC-3′ | 602 | |
| MnSODRw | 5′-CATGTTCCCACGCATCCTAT-3′ | ||||
| CYB | CytbFw | 5 | 5′-CCCAGAATTCTCGTTTGGTCTATT-3′ | 579 | 95 °C 3 min; 45 cycles: 94 °C 30 s, 55 °C 45 s, 72 °C 45 s; 72 °C 7 min |
| CytbRw | 5′-AAGAGGTCTAAAAGCAGAACCTCAA-3′ | ||||
| mt26S | PneumoLSU_F | - | 5′-TCAGGTCGAACTGGTGTACG-3′ | 297 | |
| PneumoLSU_R | 5′-TGTTCCAAGCCCACTTCTT-3′ |
| Strain Number | Country of Origin | Date of Sample | β-TUB | CYB | mt26S | SOD | Sequence Type |
|---|---|---|---|---|---|---|---|
| HVH21 | Spain | Jan 2015 | 2 | 1 | 3 | 1 | ST21 |
| HVH22 | Spain | Jan 2015 | 2 | 1 | 3 | 1 | ST21 |
| Case 63 | Australia | Dec 2016 | 1 | 3 | 1 | 4 | ST42 |
| Case 71 | New Zealand | May 2017 | 4 | 2 | 2 | 4 | ST44 |
| 1794 | Chile | Feb 2011 | 2 | 5 | 4 | 1 | ST7 |
| 2165 | Chile | Oct 2014 | 2 | 2 | 4 | 5 | ST2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasic, L.; Goterris, L.; Guerrero-Murillo, M.; Irinyi, L.; Kan, A.; Ponce, C.A.; Vargas, S.L.; Martin-Gomez, M.T.; Meyer, W. Consensus Multilocus Sequence Typing Scheme for Pneumocystis jirovecii. J. Fungi 2020, 6, 259. https://doi.org/10.3390/jof6040259
Pasic L, Goterris L, Guerrero-Murillo M, Irinyi L, Kan A, Ponce CA, Vargas SL, Martin-Gomez MT, Meyer W. Consensus Multilocus Sequence Typing Scheme for Pneumocystis jirovecii. Journal of Fungi. 2020; 6(4):259. https://doi.org/10.3390/jof6040259
Chicago/Turabian StylePasic, Lana, Lidia Goterris, Mercedes Guerrero-Murillo, Laszlo Irinyi, Alex Kan, Carolina A. Ponce, Sergio L. Vargas, M. Teresa Martin-Gomez, and Wieland Meyer. 2020. "Consensus Multilocus Sequence Typing Scheme for Pneumocystis jirovecii" Journal of Fungi 6, no. 4: 259. https://doi.org/10.3390/jof6040259