Salt Solution Treatments Trigger Antioxidant Defense Response against Gray Mold Disease in Table Grapes

 ,
,  ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Effect of Salts on Inoculated Berries (Indirect Antifungal Activity of Salts)
2.2. Plant Material, Crude Extract and Experimental Design
2.3. Defensive Enzyme Assays
2.3.1. Sample Preparation
2.3.2. Superoxide Dismutase (SOD) Enzyme Assays
2.3.3. Ascorbate Peroxidase (APX) Enzyme Assays
2.3.4. Peroxidase (POD) Enzyme Assays
2.4. Non-Enzymatic Assays
2.4.1. Sample Preparation
2.4.2. Total Phenolic Content
2.4.3. Total Flavonoid Content
2.5. Statistical Analysis
3. Results
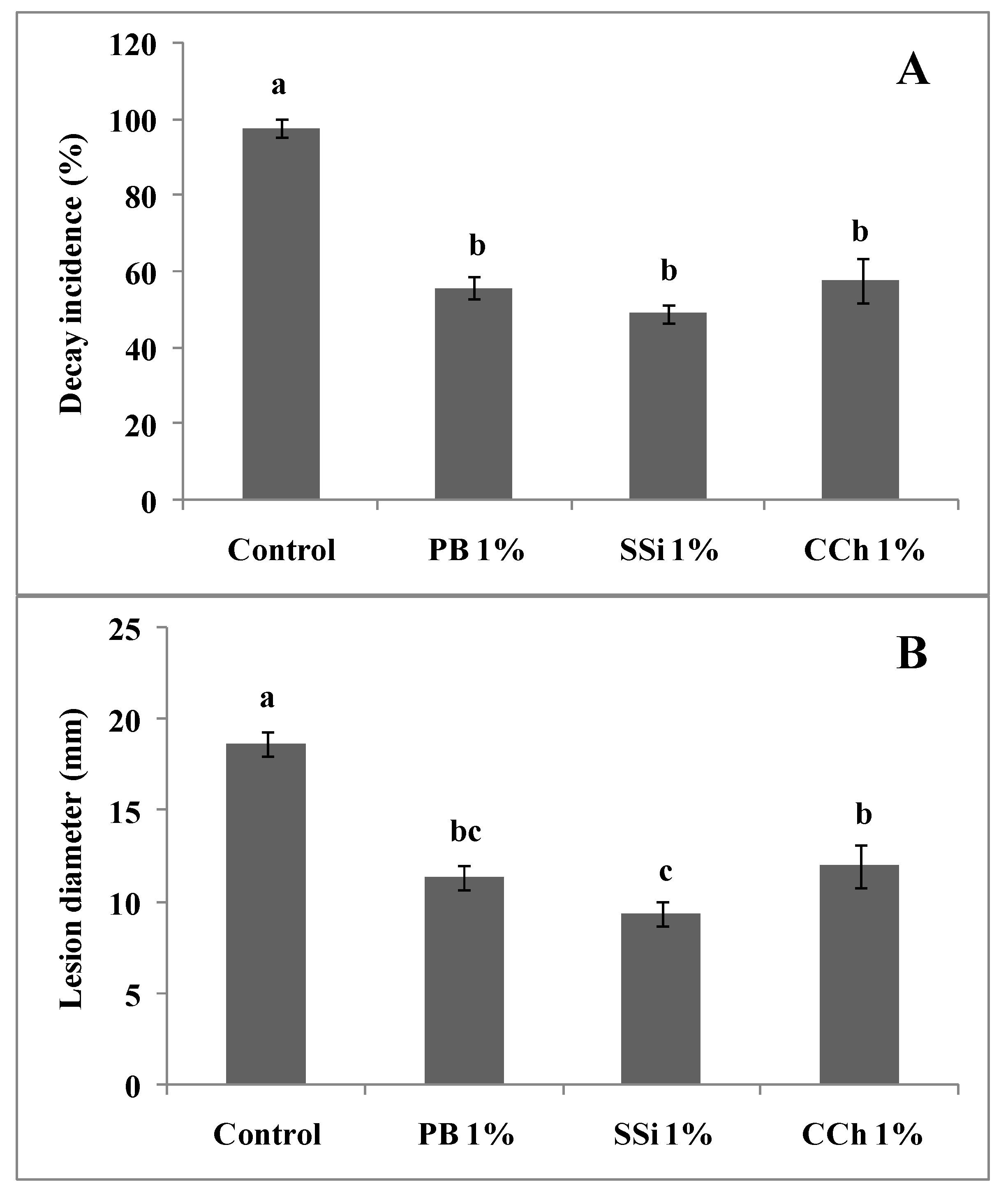
3.1. Effect of Salts on Inoculated Berries (Indirect Antifungal Activity of Salts)
3.2. Defensive Enzyme Assays
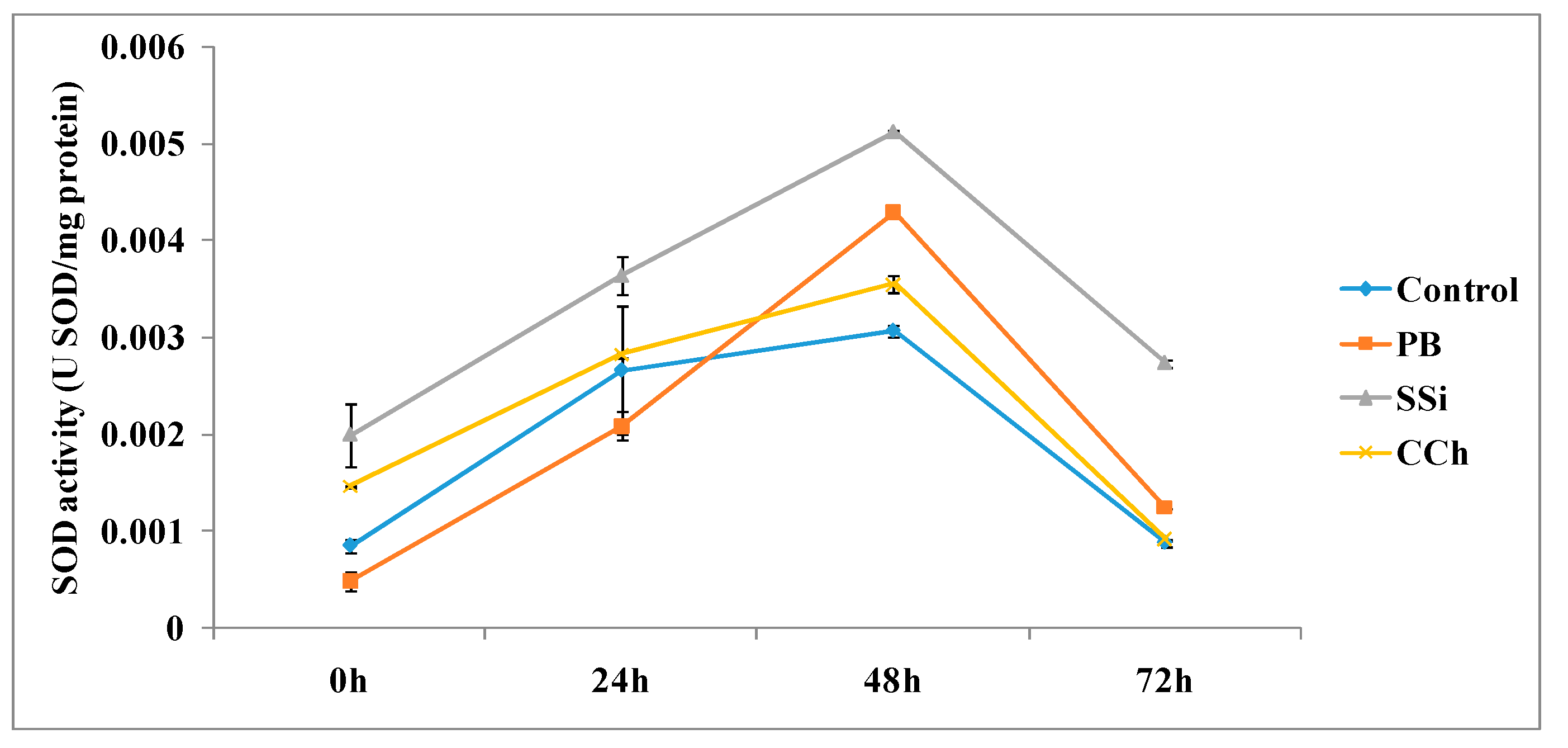
3.2.1. SOD Enzyme Assays
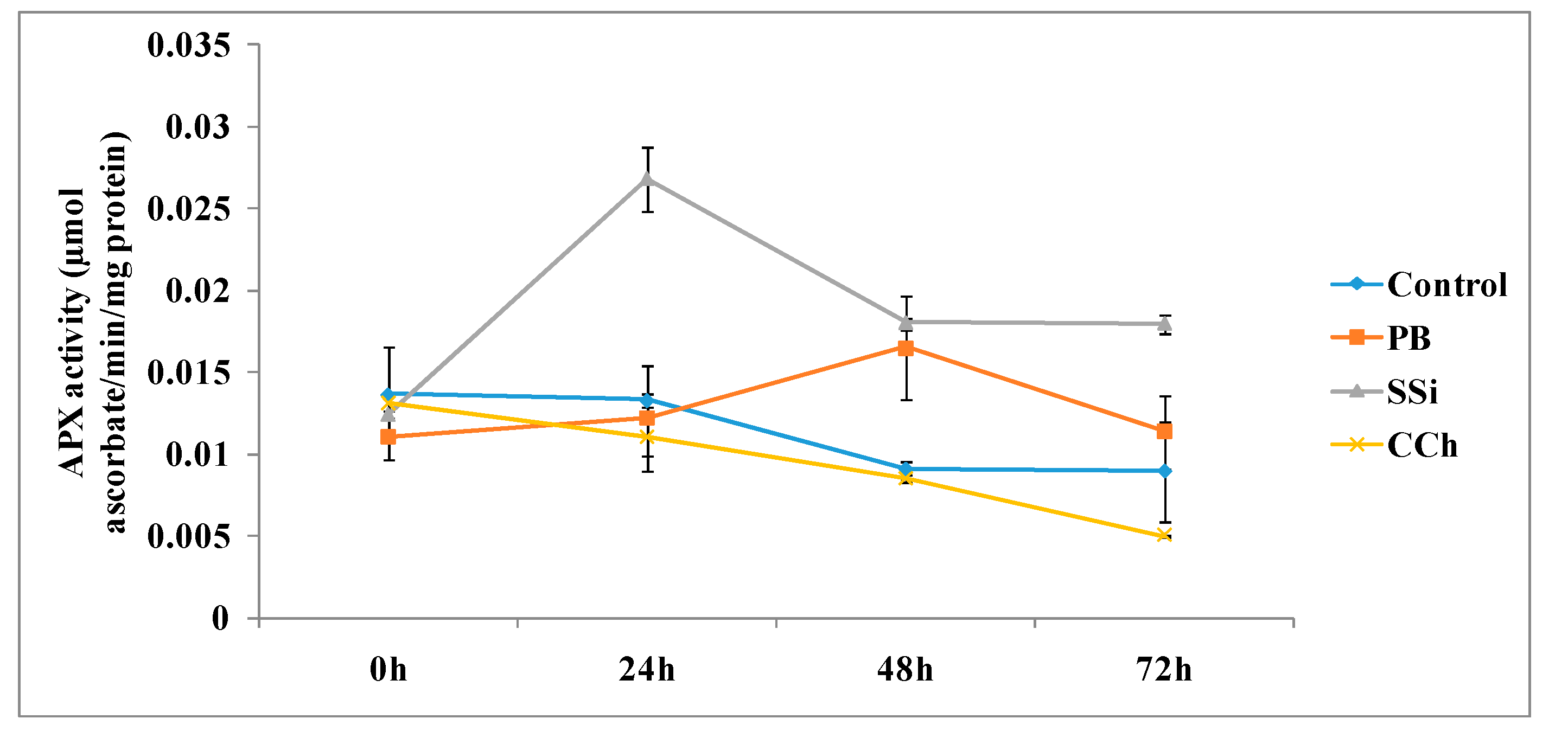
3.2.2. APX Enzyme Assays
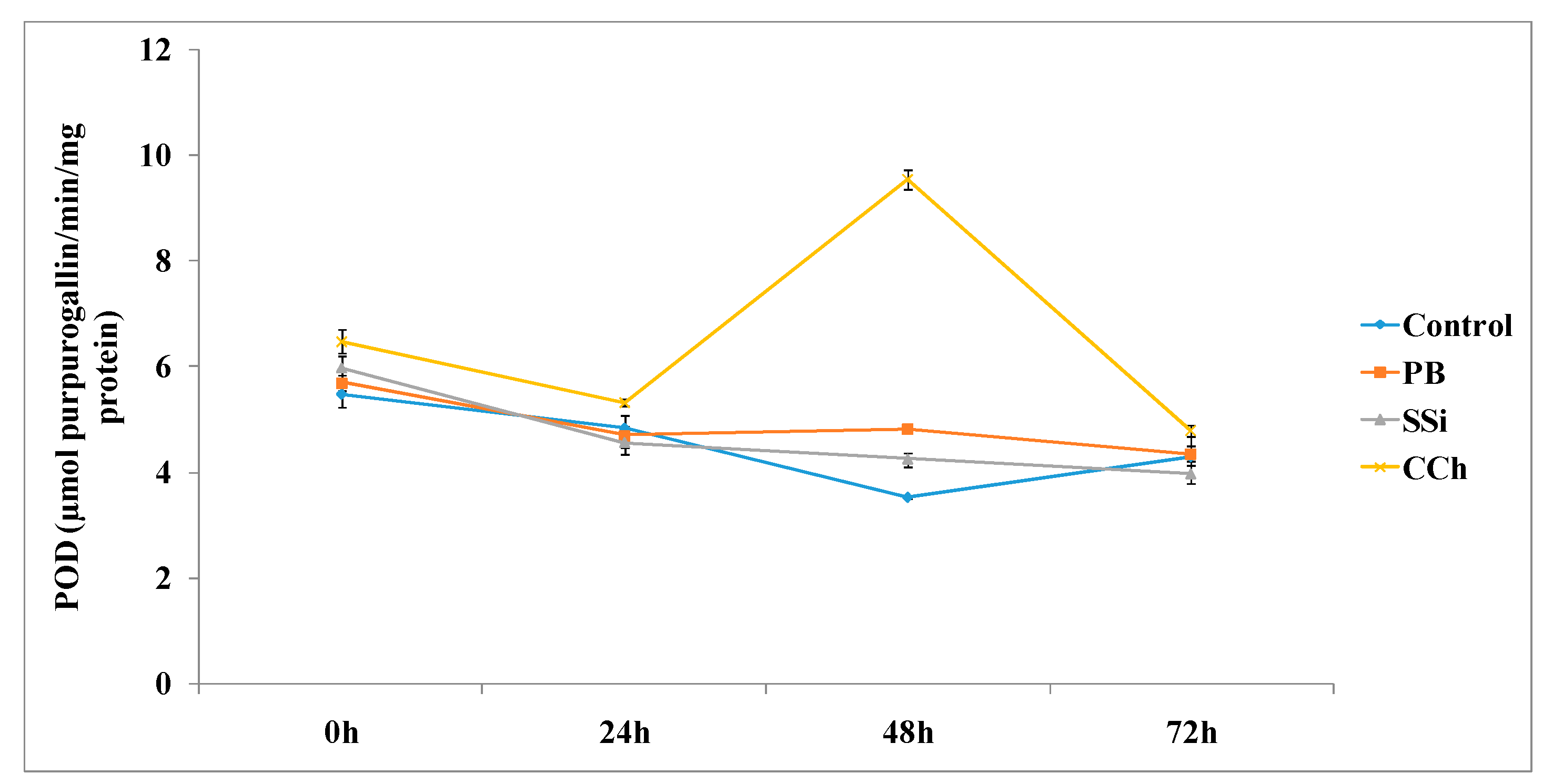
3.2.3. POD Enzyme Assays
3.3. Non-Enzymatic Assays
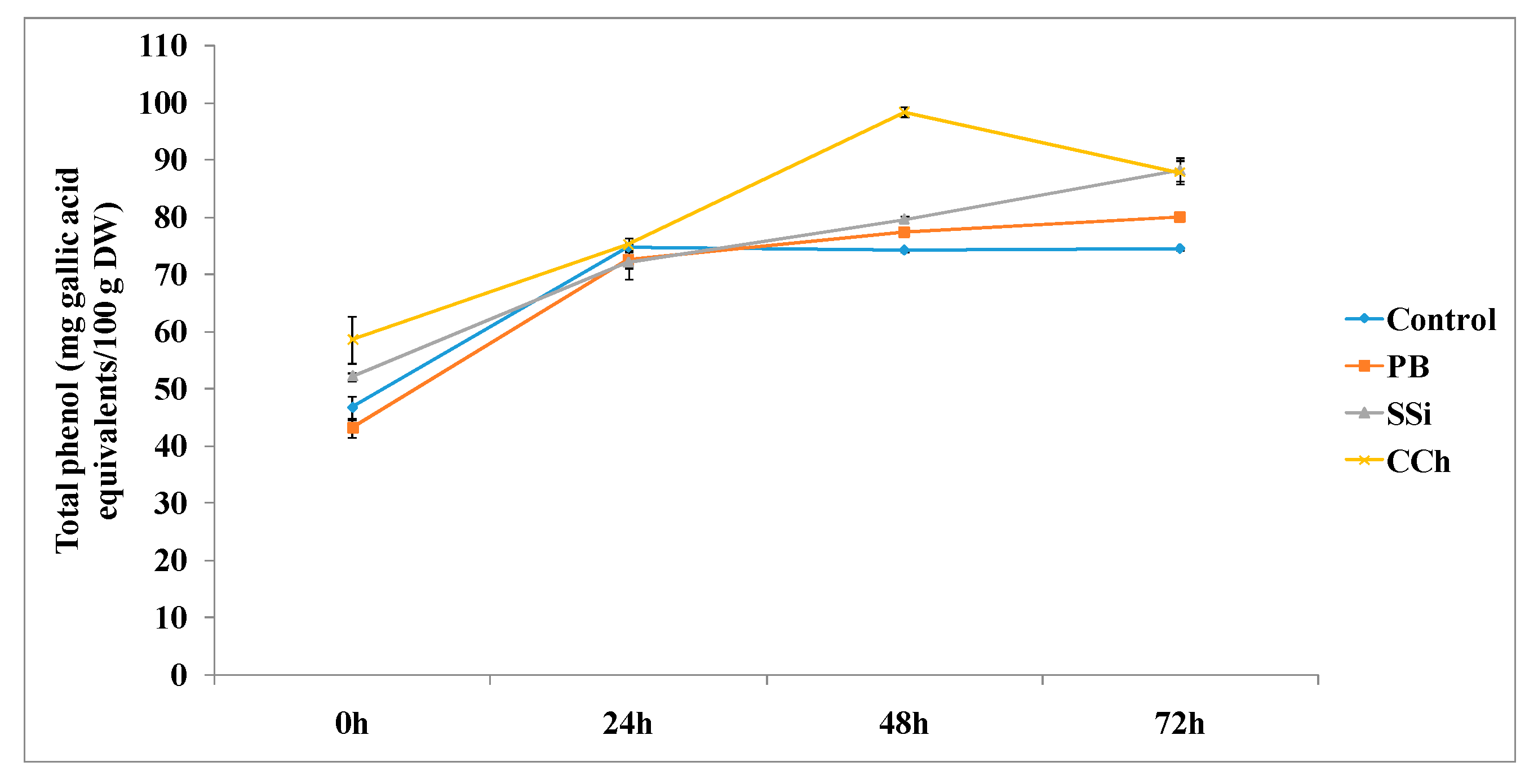
3.3.1. Total Phenolic Content
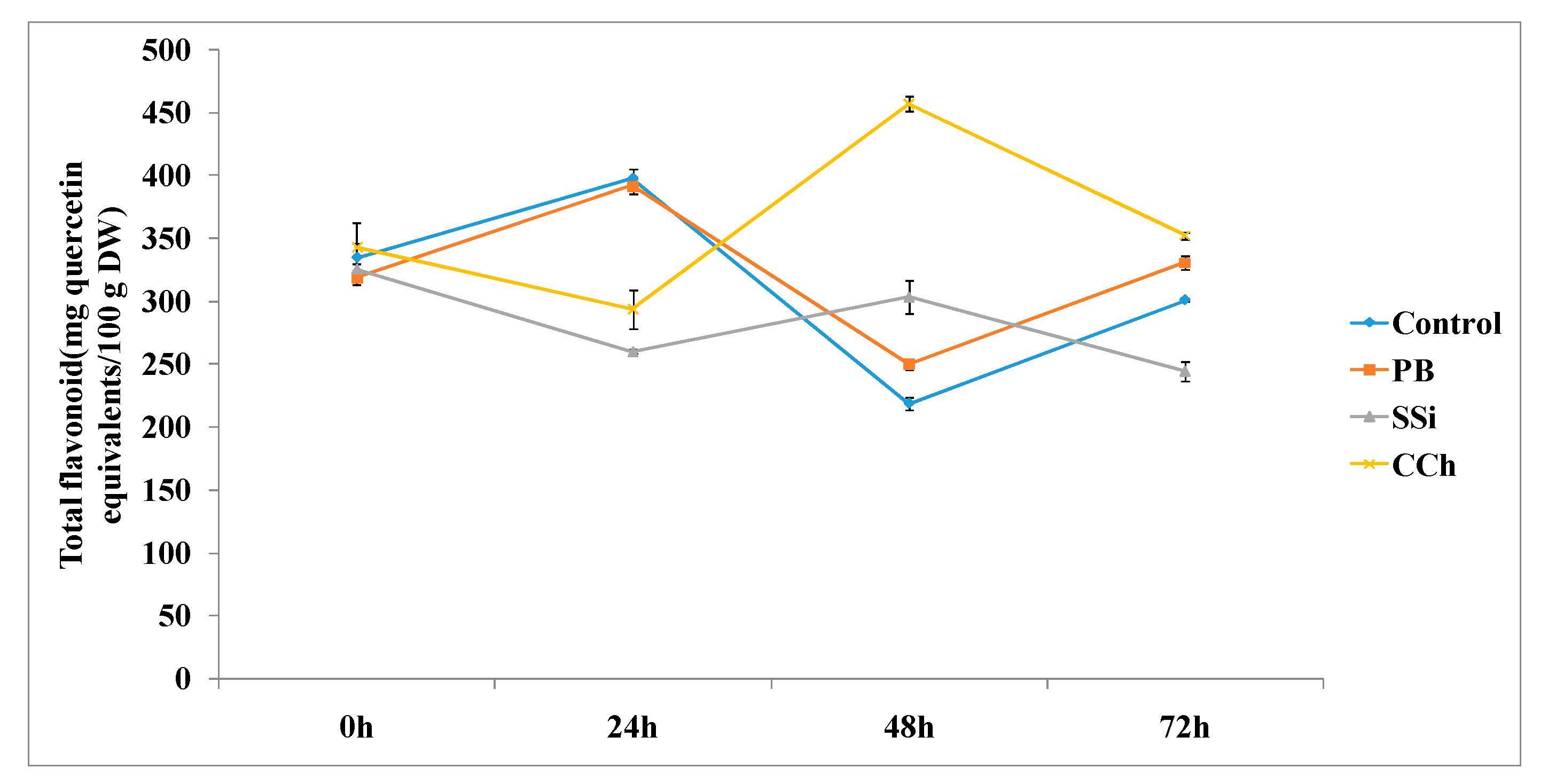
3.3.2. Total Flavonoid Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hashim, A.F.; Youssef, K.; Abd-Elsalam, K.A. Ecofriendly nanomaterials for controlling gray mold of table grapes and maintaining postharvest quality. Eur. J. Plant Pathol. 2019, 154, 377–388. [Google Scholar] [CrossRef]
- Feliziani, E.; Lichter, A.; Smilanick, J.L.; Ippolito, A. Disinfecting agents for controlling fruit and vegetable diseases after harvest. Postharvest Biol. Technol. 2016, 122, 53–69. [Google Scholar] [CrossRef]
- Youssef, K. Fungicidal efficacy of chemically-produced copper nanoparticles against Penicillium digitatum and Fusarium solani on citrus fruit. Philip. Agric. Sci. 2017, 100, 69–78. [Google Scholar]
- Nigro, F.; Schena, L.; Ligorio, A.; Pentimone, I.; Ippolito, A.; Salerno, M.G. Control of table grape storage rots by pre-harvest applications of salts. Postharvest Biol. Technol. 2006, 42, 142–149. [Google Scholar] [CrossRef]
- Youssef, K.; Roberto, S.R. Salt strategies to control Botrytis mold of ‘Benitaka’ table grapes and to maintain fruit quality during storage. Postharvest Biol. Technol. 2014, 95, 95–102. [Google Scholar] [CrossRef]
- Fallanaj, F.; Sanzani, S.M.; Youssef, K.; Zavanella, C.; Salerno, M.G.; Ippolito, A. A new perspective in controlling postharvest citrus rots: The use of electrolyzed water. Acta Hortic. 2015, 1065, 1599–1606. [Google Scholar] [CrossRef]
- Salem, E.A.; Youssef, K.; Sanzani, S.M. Evaluation of alternative means to control postharvest Rhizopus rot of peaches. Sci. Hortic. 2016, 198, 86–90. [Google Scholar] [CrossRef]
- Hussien, A.; Ahmed, Y.; Al-Essawy, A.-H.; Youssef, K. Evaluation of different salt-amended electrolysed water to control postharvest moulds of citrus. Trop. Plant Pathol. 2017, 43, 10–20. [Google Scholar] [CrossRef]
- Youssef, K.; Mustafa, Z.M.M.; Kamel, M.A.M.; Mounir, G.A. Cigar end rot of banana caused by Musicillium theobromae and its control in Egypt. Arch. Phytopathol. Plant Prot. 2020, 53, 162–177. [Google Scholar] [CrossRef]
- Youssef, K.; Roberto, S.R.; De Oliveira, A.G. Ultra-Structural Alterations in Botrytis cinerea-The Causal Agent of Gray Mold-Treated with Salt Solutions. Biomolecules 2019, 9, 582. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.R.; Newton, A.C.; Lyon, G.D. Induced resistance: Helping plants to help themselves. Biologist 2005, 52, 28–33. [Google Scholar]
- González-Aguilar, G.A.; Villa-Rodriguez, J.A.; Ayala-Zavala, J.F.; Yahia, E.M. Improvement of the antioxidant status of tropical fruits as a secondary response to some postharvest treatments. Trends Food Sci. Technol. 2010, 21, 475–482. [Google Scholar] [CrossRef]
- Siringam, K.; Juntawong, N.; Cha-Um, S.; Kirdmanee, C. Salt stress induced ion accumulation, ion homeostasis, membrane injury and sugar contents in salt-sensitive rice (Oryza sativa L. spp. indica) roots under isoosmotic conditions. Afr. J. Biotechnol. 2011, 10, 1340–1346. [Google Scholar]
- Ballester, A.-R.; Lafuente, M.T.; González-Candelas, L. Spatial study of antioxidant enzymes, peroxidase and phenylalanine ammonia-lyase in the citrus fruit–Penicilliumdigitatum interaction. Postharvest Biol. Technol. 2006, 39, 115–124. [Google Scholar] [CrossRef]
- Boguszewska, D.; Grudkowska, M.; Zagdanska, B. Drought-Responsive Antioxidant Enzymes in Potato (Solanum tuberosum L.). Potato Res. 2010, 53, 373–382. [Google Scholar] [CrossRef]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic Stress Tolerance in Plants: Myriad Roles of Ascorbate Peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [Green Version]
- De Pinto, M.C.; De Gara, L. Changes in the ascorbate metabolism of apoplastic and symplastic spaces are associated with cell differentiation. J. Exp. Bot. 2004, 55, 2559–2569. [Google Scholar] [CrossRef]
- Radhakrishnan, N.; Alphonse, A.J.; Balasubramanian, R. Effect of Acibenzolar-Smethyl (ASM) pre-treatment in inducing resistance against Pythium aphanidermatum infection in Curcuma longa. Crop Prot. 2011, 30, 24–32. [Google Scholar] [CrossRef]
- Youssef, K.; Ahmed, Y.; Ligorio, A.; D’Onghia, A.M.; Nigro, F.; Ippolito, A. First report of Penicillium ulaiense as a postharvest pathogen of orange fruit in Egypt. Plant. Pathol. 2010, 59, 1174. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases. Plant. Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Tiepo, A.N.; Constantino, L.V.; Madeira, T.B.; Gonçalves, L.S.A.; Pimenta, J.A.; Bianchini, E.; De Oliveira, A.L.M.; Oliveira, H.C.; Stolf-Moreira, R. Plant growth-promoting bacteria improve leaf antioxidant metabolism of drought-stressed Neotropical trees. Planta 2020, 251, 83. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant. Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Peixoto, P.H.P.; Cambraia, J.; Sant’Anna, R.; Mosquim, P.R.; Moreira, M.A. Aluminum effects on lipid peroxidation and on the activities of enzymes of oxidative metabolism in sorghum. Braz. J. Plant Physiol. 1999, 11, 137–143. [Google Scholar]
- Vazquez, G.; Fontenla, E.; Santos, J.; Freire, M.S.; González-Alvarez, J.; Antorrena, G. Antioxidant activity and phenolic content of chestnut (Castanea sativa) shell and eucalyptus (Eucalyptus globulus) bark extracts. Ind. Crop. Prod. 2008, 28, 279–285. [Google Scholar] [CrossRef]
- Swain, T.; Hillis, W.E. The phenolic constituents of Prunus domestica. I.—The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 1959, 10, 63–68. [Google Scholar] [CrossRef]
- Gurnani, N.; Gupta, M.; Mehta, D.; Mehta, B.K. Chemical composition, total phenolic and flavonoid contents, and in vitro antimicrobial and antioxidant activities of crude extracts from red chilli seeds (Capsicum frutescens L.). J. Taibah Univ. Sci. 2016, 10, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, K.; Kanematsu, S.; Asada, K.; Myśliwa-Kurdziel, B.; Barthélemy, X.; Strzałka, K.; Franck, F. Generation of Superoxide Anion and Localization of CuZn-Superoxide Dismutase in the Vascular Tissue of Spinach Hypocotyls: Their Association with Lignification. Plant. Cell Physiol. 1997, 38, 1118–1126. [Google Scholar] [CrossRef] [Green Version]
- Thinakaran, A.M.; Adaikkalam, C.; Asaithambi, V.; Nagarathinam, R. Sulfamethoxazole activates antioxidant defense response against Alternaria solani infection in Solanum lycopersicum Linn. Arch. Phytopathol. Plant. Prot. 2020, 1–22. [Google Scholar] [CrossRef]
- Chan, Z.; Tian, S. Induction of H2O2-metabolizing enzymes and total protein synthesis by antagonistic yeast and salicylic acid in harvested sweet cherry fruit. Postharvest Biol. Technol. 2006, 39, 314–320. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, L.; Zhao, J.; Bi, Y. Use of silicon oxide and sodium silicate for controlling Trichotheciumroseum postharvest rot in Chinese cantaloupe (Cucumis melo L.). Int. J. Food Sci. Technol. 2007, 42, 1012–1018. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2002; pp. 283–310. [Google Scholar]
- Ishikawa, T.; Shigeoka, S. Recent Advances in Ascorbate Biosynthesis and the Physiological Significance of Ascorbate Peroxidase in Photosynthesizing Organisms. Biosci. Biotechnol. Biochem. 2008, 72, 1143–1154. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, G.; Kata, A.; Goc, A.; Szechyńska-Hebda, M.; Skrzypek, E. Characteristics of the plant ascorbate peroxidase family. Acta Biol. Crac. Ser. Bot. 2007, 49, 7–17. [Google Scholar]
- Suzuki, N.; Miller, G.; Sejima, H.; Harper, J.; Mittler, R. Enhanced seed production under prolonged heat stress conditions in Arabidopsis thaliana plants deficient in cytosolic ascorbate peroxidase 2. J. Exp. Bot. 2012, 64, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Erkan, M.; Wang, S.Y.; Wang, C.Y. Effect of UV treatment on antioxidant capacity, antioxidant enzyme activity and decay in strawberry fruit. Postharvest Biol. Technol. 2008, 48, 163–171. [Google Scholar] [CrossRef]
- Youssef, K.; Sanzani, S.M.; Ligorio, A.; Ippolito, A.; Terry, L.A. Sodium carbonate and bicarbonate treatments induce resistance to postharvest green mold on citrus fruit. Postharv. Biol. Technol. 2014, 87, 61–69. [Google Scholar] [CrossRef]
- Hammerschmidt, R.; Nuckles, E.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant. Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Droby, S.; Chalutz, E.; Horev, B.; Cohen, L.; Gaba, V.; Wilson, C.L.; Wisniewski, M. Factors affecting UV-induced resistance in grapefruit against the green mold decay caused by Penicillium digitatum. Plant Pathol. 1993, 42, 418–424. [Google Scholar] [CrossRef]
- Cao, J.; Jiang, W.; He, H. Induced Resistance in Yali Pear (Pyrus bretschneideri Rehd.) Fruit against Infection by Penicillium expansum by Postharvest Infiltration of Acibenzolar-S-methyl. J. Phytopathol. 2005, 153, 640–646. [Google Scholar] [CrossRef]
- Nikraftar, F.; Taheri, P.; Rastegar, M.F.; Tarighi, S. Tomato partial resistance to Rhizoctonia solani involves antioxidative defense mechanisms. Physiol. Mol. Plant. Pathol. 2013, 81, 74–83. [Google Scholar] [CrossRef]
- Radhakrishnan, N.; Balasubramanian, R. Salicylic acid induced defense responses in Curcuma longa (L.) against Pythium aphanidermatum infection. Crop Prot. 2009, 28, 974–979. [Google Scholar] [CrossRef]
- Liu, H.; Guo, J.; Luo, L.; Liu, P.; Wang, B.; Cheng, Y.; Deng, B.; Long, C. Improvement of Hanseniaspora uvarum biocontrol activity against gray mold by the addition of ammonium molybdate and the possible mechanisms involved. Crop Prot. 2010, 29, 277–282. [Google Scholar] [CrossRef]
- Hiraga, S.; Sasaki, K.; Ito, H.; Ohashi, Y.; Matsui, H. A Large Family of Class III Plant Peroxidases. Plant Cell Physiol. 2001, 42, 462–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youwei, Y.; Yinzhe, R. Grape Preservation Using Chitosan Combined with?-Cyclodextrin. Int. J. Agron. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Al-Qurashi, A.D.; Awad, M.A. Postharvest chitosan treatment affects quality, antioxidant capacity, antioxidant compounds and enzymes activities of ‘El-Bayadi’ table grapes after storage. Sci. Hortic. 2015, 197, 392–398. [Google Scholar] [CrossRef]
- Raimbault, A.-K.; Marie-Alphonsine, P.-A.; Horry, J.-P.; Francois-Haugrin, M.; Romuald, K.; Soler, A. Polyphenol Oxidase and Peroxidase Expression in Four Pineapple Varieties (Ananas comosus L.) after a Chilling Injury. J. Agric. Food Chem. 2011, 59, 342–348. [Google Scholar] [CrossRef]
- Passardi, F.; Penel, C.; Dunand, C. Performing the paradoxical: How plant peroxidases modify the cell wall. Trends Plant. Sci. 2004, 9, 534–540. [Google Scholar] [CrossRef]
- Yamauchi, N. Quality Maintenance of Postharvest Horticultural Crops by Stress Treatments and Approach for the Elucidation of Its Mechanism. J. Jpn. Soc. Hortic. Sci. 2013, 82, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ghorbani, B.; Pakkish, Z.; Najafzadeh, R. Shelf life improvement of grape (Vitis vinifera L. cv. Rish Baba) using nitric oxide (NO) during chilling damage. Int. J. Food Prop. 2017, 20, S2750–S2763. [Google Scholar] [CrossRef]
- Petriccione, M.; Pagano, L.; Forniti, R.; Zampella, L.; Mastrobuoni, F.; Scortichini, M.; Mencarelli, F. Postharvest treatment with chitosan affects the antioxidant metabolism and quality of wine grape during partial dehydration. Postharvest Biol. Technol. 2018, 137, 38–45. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, K.; Roberto, S.R.; Tiepo, A.N.; Constantino, L.V.; de Resende, J.T.V.; Abo-Elyousr, K.A.M. Salt Solution Treatments Trigger Antioxidant Defense Response against Gray Mold Disease in Table Grapes. J. Fungi 2020, 6, 179. https://doi.org/10.3390/jof6030179
Youssef K, Roberto SR, Tiepo AN, Constantino LV, de Resende JTV, Abo-Elyousr KAM. Salt Solution Treatments Trigger Antioxidant Defense Response against Gray Mold Disease in Table Grapes. Journal of Fungi. 2020; 6(3):179. https://doi.org/10.3390/jof6030179
Chicago/Turabian StyleYoussef, Khamis, Sergio Ruffo Roberto, Angélica Nunes Tiepo, Leonel Vinicius Constantino, Juliano Tadeu Vilela de Resende, and Kamal A.M. Abo-Elyousr. 2020. "Salt Solution Treatments Trigger Antioxidant Defense Response against Gray Mold Disease in Table Grapes" Journal of Fungi 6, no. 3: 179. https://doi.org/10.3390/jof6030179