Do Pentraxins Bind to Fungi in Invasive Human Gastrointestinal Candidiasis?

Abstract
:1. Introduction
2. Methods
2.1. Staining Human Tissue
2.2. Immunohistochemistry
2.3. Clinical Information
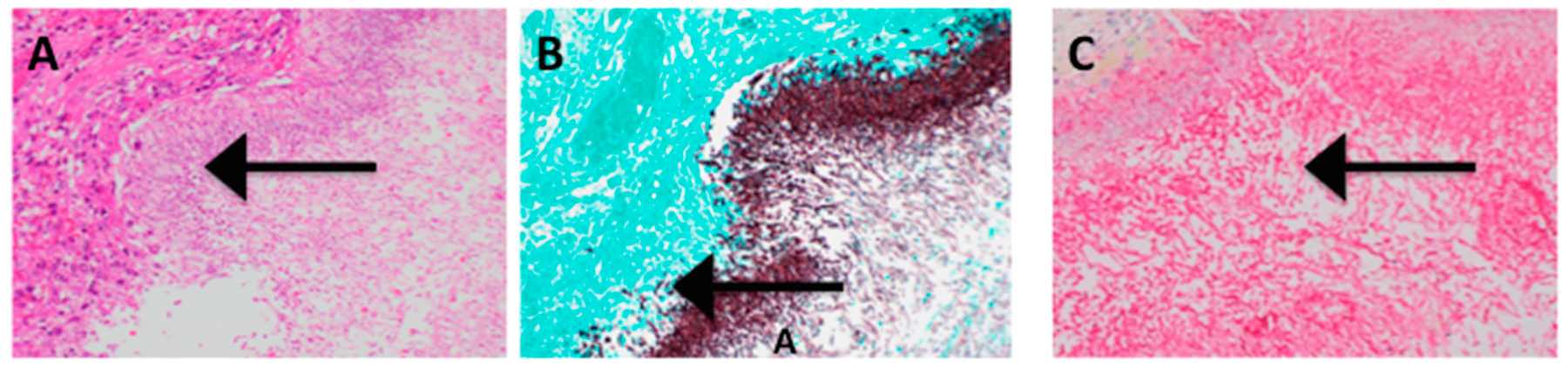
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Du Clos, T.W. Pentraxins: Structure, functions, and role in inflammation. ISRN Inflamm. 2013. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Singh, P.P.; Bottazzi, B.; Garlanda, C.; Mantovani, A. Pattern recognition by pentraxins. Adv. Exp. Med. Biol. 2009, 653, 98–116. [Google Scholar] [PubMed]
- Alsteen, D.; Garcia, M.C.; Lipke, P.N.; Dufrene, Y. Force-induced formation and propagation of adhesion nanodomains in living cells. Proc. Natl. Acad. Sci. USA 2010, 107, 20744–20749. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, K.B.; Garcia, M.C.; Sobonya, R.; Lipke, P.N.; Klotz, S.A. New features of invasive candidiasis in humans: Amyloid formation by fungi and deposition of serum amyloid P component by the host. J. Infect. Dis. 2012, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sherman, M.C.; Lysak, N.; Filonenko, A.; Richards, H.; Sobonya, R.E.; Klotz, S.A.; Lipke, P.N. Peptide detection of fungal functional amyloids in infected tissue. PLoS ONE 2014, 9, e86067. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sherman, M.C.; Lundberg, T.; Sobonya, R.E.; Lipke, P.N.; Klotz SAGarcia-Sherman, M.C.; Lundberg, T.; Sobonya, R.E.; Lipke, P.N.; Klotz, S.A. A unique biofilm in human deep mycoses: Fungal amyloid is bound by host serum amyloid P component. NPJ Biofilms Microbiomes 2015, 1, 15009. [Google Scholar] [CrossRef] [PubMed]
- Thorn, J.L.; Gilchrist, K.B.; Sobonya, R.E.; Gaur, N.K.; Lipke, P.N.; Klotz, S.A. Postmortem candidemia. A reliable marker of disseminated candidiasis. J. Clin. Pathol. 2010, 63, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Cox, N.; Pilling, D.; Gomer, R. Serum amyloid P: A systemic regulator of the innate immune response. J. Leukoc. Biol. 2014, 96, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Pepys, M.B. C-reactive protein fifty years on. Lancet 1981, 317, 653–657. [Google Scholar] [CrossRef]
- Thompson, D.; Pepys, M.B.; Wood, S.P. The physiological structure of human C-reactive protein and its complex with phosphocholine. Structure 1999, 7, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Garlanda, C. Non-redundant role of the long pentraxin PTX3 in anti-fungal innate immune response. Nature 2002, 420, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Lipke, P.; Klotz, S. Pathogenic microbial amyloids: Their function and host response. OA Microbiol. 2013, 1, 2. [Google Scholar] [PubMed]
- Klotz, S.A.; Sobonya, R.E.; Lipke, P.N.; Garcia-Sherman, M.C. Serum amyloid P component and systemic fungal infections. Does it protect the host or is a Trojan horse? Open Forum Infect. Dis. 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Pepys, M.B. Pathogenesis, diagnosis and treatment of systemic amyloidosis. Philos. Trans. R Soc. Lond. B Biol. Sci. 2001, 356, 203–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikolajek, H.; Kolstoe, S.E.; Pye, V.E.; Mangione, P.; Pepys, M.B.; Wood, S.P. Structural basis of ligand specificity in human pentaxins, C-reactive protein and serum amyloid P component. J. Mol. Recognit. 2011, 24, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Ballmann, G.E.; Caffin, W.L. Lipid synthesis during reinitiation of growth from stationary phase cultures of Candida albicans. Mycopathologica 1979, 30, 39–43. [Google Scholar] [CrossRef]
- Christner, R.B.; Mortensen, R.F. Binding of human serum amyloid P-component to phosphocholine. Arch. Biochem. Biophys. 1994, 14, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Orlean, P. Architecture and biosynthesis of the Saccharomyces cerevisiae cell wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef] [PubMed]
- Bottazzi, B.; Doni, A.; Garland, C.; Montovani, A. An integrated view of humoral innate immunity: Pentraxins as a paradigm. Ann. Rev. Immunol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.D.; Gray, C.A.; Shankland, G.S. Opsonic effect of C-reactive protein on phagocytosis and intracellular killing of virulent and attenuated strains of Candida albicans by human neutrophils. FEMS Microbiol. Immunol. 1991, 76, 341–344. [Google Scholar] [CrossRef]
- Caironi, P.; Masson, S.; Mauri, T.; Bottazzi, B.; Leone, R.; Magnoli, M.; Barlera, S.; Mamprin, F.; Fedele, A.; Mantovani, A.; et al. ALBIOS Biomarkers Study Investigators. Pentraxin 3 in patients with severre sepsis or shock: The ALBIOS trial. Eur. J. Clin. Investig. 2017, 47, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Quan, L.; Fujita, M.Q.; Zhu, B.-L.; Ishida, K.; Maeda, H. Immunohistochemical distribution of C-reactive protein in the hepatic tissue in forensic autopsy. Forensic Sci. Int. 2000, 113, 177–182. [Google Scholar] [CrossRef]
- Lgeng’o, J.A.; Cohen, D.L.; Sayi, J.G.; Matuja, W.B.; Chande, H.M.; Kitinya, J.N.; Kimani, J.K.; Friedland, R.P.; Mori, H.; Kalaria, R.N. Cerebral amyloid B protein depositis and other Alzheimer lesions in non-demented elderly East Africans. Brain Pathol. 1996, 6, 101–108. [Google Scholar]
- Savchenko, A.S.; Imamura, M.; Ohashi, R.; Jiang, S.; Kawasaki, T.; Hasegawa, G.; Emura, I.; Iwanari, H.; Sagara, M.; Tanaka, T.; et al. Expression of pentraxin 3 (PTX3) in human atherosclerotic lesions. J. Pathol. 2008, 215, 48–55. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Candidal Lesion(s) * | Reduced Cellular Immunity | Cause of Death; Associated Problem(s) | Microbiology Cultures ** | Serum Amyloid P (SAP) on Fungi | Pentraxin 3 (PTX 3) on Fungi | C-Reactive Protein (CRP) on Fungi | PMN in Lesion *** | Lymph cells in Lesion *** |
|---|---|---|---|---|---|---|---|---|
| Esophagitis; myocarditis encephalitis | No | Prematurity necrotizing enterocolitis; disseminated candididasis | C. albicans, E. coli in blood | + | − | − | 1+ | 1+ |
| Esophagitis | No | Ischemic heart disease | Group D Streptococcus in blood | + | − | − | 0 | 0 |
| Esophagitis, gastritis | No | Sepsis; non-Hodgkins | C. albicans, Enterococcus in blood | + | − | − | 1+ | 1+ |
| Esophagitis, gastritis | No | Myocardial infarction | Yeast in lung (only yeasts were found in tissue) | - | − | − | 2+ | 2+ |
| Esophagitis | No | Pulmonary embolism | + | − | − | 1+ | 1+ | |
| Gastric ulcer | Yes | Sepsis; exsanguination; treated acute myelomonocytic leukemia | Candida sp., Enterococcus, Xanthomonas maltophilia in blood | + | − | − | 0 | 1+ |
| Esophagitis | Yes | Disseminated aspergillosis; pneumonitis; pancytopenia; acute myeloid leukemia | + | − | − | 2+ | 1+ | |
| Esophagitis | Yes | Hemorrhagic enterocolitis; acute lymphocytic leukemia; graft versus host disease | Disseminated aspergillosis | + | − | − | 0 | 1+ |
| Colitis | No | Sepsis; pseudomembranous colitis | C. krusei, Group D Streptococcus in blood; Aspergillus in tissue | + | − | − | 0 | 1+ |
| Gastric ulcer | No | Gastrointestinal hemorrhage; gastric ulcer | + | − | − | 3+ | 1+ | |
| Esophagitis | No | Ischemic heart disease | + | − | − | 0 | 0 | |
| Duodenal ulcer | No | Exsanguination from peptic ulcer | C. albicans in blood; yeasts in lung | + | − | − | 1+ | 1+ |
| Esophagitis | Yes | Sepsis; neutropenia; post-operative cancer resection of colon | Escherichia coli; viridans Streptococcus in blood; yeasts in lung | + | − | − | 0 | 1+ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golconda, U.; Sobonya, R.E.; Klotz, S.A. Do Pentraxins Bind to Fungi in Invasive Human Gastrointestinal Candidiasis? J. Fungi 2018, 4, 111. https://doi.org/10.3390/jof4030111
Golconda U, Sobonya RE, Klotz SA. Do Pentraxins Bind to Fungi in Invasive Human Gastrointestinal Candidiasis? Journal of Fungi. 2018; 4(3):111. https://doi.org/10.3390/jof4030111
Chicago/Turabian StyleGolconda, Umamaheshwari, Richard E. Sobonya, and Stephen A. Klotz. 2018. "Do Pentraxins Bind to Fungi in Invasive Human Gastrointestinal Candidiasis?" Journal of Fungi 4, no. 3: 111. https://doi.org/10.3390/jof4030111