
Proteome and Dihydrorhodamine Profiling of Bronchoalveolar Lavage in Patients with Chronic Pulmonary Aspergillosis
 , , and
, , and
Abstract
:1. Introduction
2. Aim
3. Methods
3.1. Study Design
3.2. Setting
3.3. Cases and Controls
3.4. DHR-Test for Neutrophil NADPH-Generated Oxidative Burst
3.5. Mass Spectrometry
3.6. Proteome Data Processing and Statistical Analysis
4. Results
4.1. Patient Population and Baseline Data
4.2. BAL Leukocyte Composition in Patients with CPA and ILD
4.3. DHR-Profiling of BAL-Fluid and Blood Neutrophils from CPA Patients
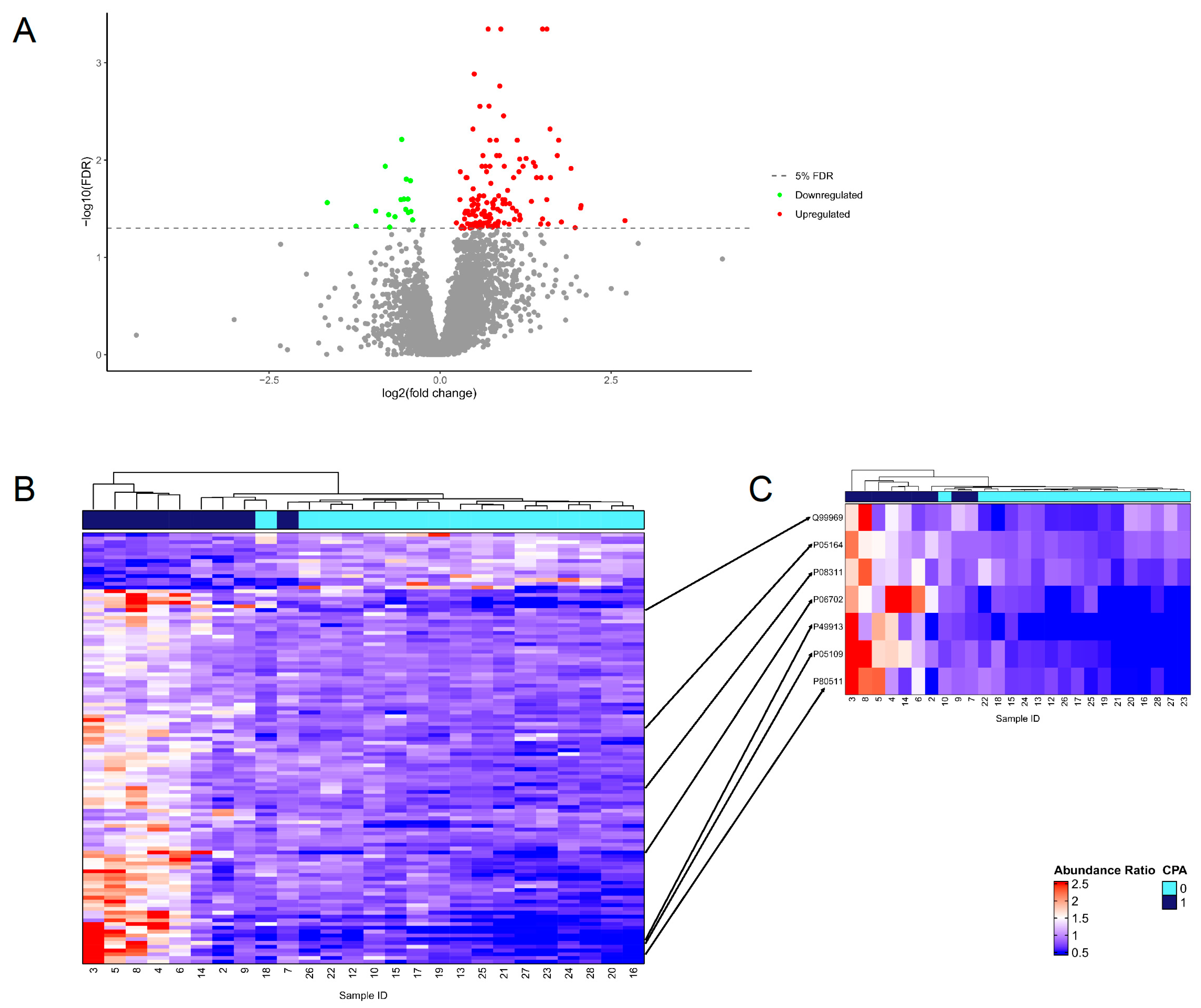
4.4. Proteome Analysis
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Denning, D.W. Global incidence and mortality of severe fungal disease. Lancet Infect. Dis. 2024. online first. [Google Scholar] [CrossRef]
- Mortensen, K.L.; Denning, D.W.; Arendrup, M.C. The burden of fungal disease in Denmark. Mycoses 2015, 58 (Suppl. S5), 15–21. [Google Scholar] [CrossRef]
- Kousha, M.; Tadi, R.; Soubani, A.O. Pulmonary aspergillosis: A clinical review. Eur. Respir. Rev. 2011, 20, 156–174. [Google Scholar] [CrossRef]
- Kosmidis, C.; Denning, D.W. The clinical spectrum of pulmonary aspergillosis. Thorax 2015, 70, 270–277. [Google Scholar] [CrossRef]
- Denning, D.W.; Cadranel, J.; Beigelman-Aubry, C.; Ader, F.; Chakrabarti, A.; Blot, S.; Ullmann, A.J.; Dimopoulos, G.; Lange, C. Chronic pulmonary aspergillosis: Rationale and clinical guidelines for diagnosis and management. Eur. Respir. J. 2016, 47, 45–68. [Google Scholar] [CrossRef]
- Delliere, S.; Aimanianda, V. Humoral Immunity Against Aspergillus fumigatus. Mycopathologia 2023, 188, 603–621. [Google Scholar] [CrossRef]
- Lionakis, M.S.; Drummond, R.A.; Hohl, T.M. Immune responses to human fungal pathogens and therapeutic prospects. Nat. Rev. Immunol. 2023, 23, 433–452. [Google Scholar] [CrossRef]
- Gonzales, D.A.; De Torre, C.; Wang, H.; Devor, C.B.; Munson, P.J.; Ying, S.X.; Kern, S.J.; Petraitiene, R.; Levens, D.L.; Walsh, T.J.; et al. Protein expression profiles distinguish between experimental invasive pulmonary aspergillosis and Pseudomonas pneumonia. Proteomics 2010, 10, 4270–4280. [Google Scholar] [CrossRef]
- Krel, M.; Petraitis, V.; Petraitiene, R.; Jain, M.R.; Zhao, Y.; Li, H.; Walsh, T.J.; Perlin, D.S. Host biomarkers of invasive pulmonary aspergillosis to monitor therapeutic response. Antimicrob. Agents Chemother. 2014, 58, 3373–3378. [Google Scholar] [CrossRef]
- Meyer, K.C.; Raghu, G.; Baughman, R.P.; Brown, K.K.; Costabel, U.; du Bois, R.M.; Drent, M.; Haslam, P.L.; Kim, D.S.; Nagai, S.; et al. An official American Thoracic Society clinical practice guideline: The clinical utility of bronchoalveolar lavage cellular analysis in interstitial lung disease. Am. J. Respir. Crit. Care Med. 2012, 185, 1004–1014. [Google Scholar] [CrossRef]
- Mulorz, J.; Spin, J.M.; Beck, H.C.; Tha Thi, M.L.; Wagenhauser, M.U.; Rasmussen, L.M.; Lindholt, J.S.; Tsao, P.S.C.; Steffensen, L.B. Hyperlipidemia does not affect development of elastase-induced abdominal aortic aneurysm in mice. Atherosclerosis 2020, 311, 73–83. [Google Scholar] [CrossRef]
- Palstrom, N.B.; Campbell, A.J.; Lindegaard, C.A.; Cakar, S.; Matthiesen, R.; Beck, H.C. Spectral library search for improved TMTpro labelled peptide assignment in human plasma proteomics. Proteomics 2024, 24, e2300236. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Allaire, J. RStudio: Integrated Development Environment for R; RStudio, PBC; RStudio Team: Boston, MA, USA, 2022. [Google Scholar]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Binns, D.; Dimmer, E.; Huntley, R.; Barrell, D.; O’Donovan, C.; Apweiler, R. QuickGO: A web-based tool for Gene Ontology searching. Bioinformatics 2009, 25, 3045–3046. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]
- Park, S.J.; Mehrad, B. Innate immunity to Aspergillus species. Clin. Microbiol. Rev. 2009, 22, 535–551. [Google Scholar] [CrossRef]
- Ok, M.; Einsele, H.; Loeffler, J. Genetic susceptibility to Aspergillus fumigatus infections. Int. J. Med. Microbiol. 2011, 301, 445–452. [Google Scholar] [CrossRef]
- Lambourne, J.; Agranoff, D.; Herbrecht, R.; Troke, P.F.; Buchbinder, A.; Willis, F.; Letscher-Bru, V.; Agrawal, S.; Doffman, S.; Johnson, E.; et al. Association of mannose-binding lectin deficiency with acute invasive aspergillosis in immunocompromised patients. Clin. Infect. Dis. 2009, 49, 1486–1491. [Google Scholar] [CrossRef]
- Palmer, L.B.; Greenberg, H.E.; Schiff, M.J. Corticosteroid treatment as a risk factor for invasive aspergillosis in patients with lung disease. Thorax 1991, 46, 15–20. [Google Scholar] [CrossRef]
- Caffrey, A.K.; Lehmann, M.M.; Zickovich, J.M.; Espinosa, V.; Shepardson, K.M.; Watschke, C.P.; Hilmer, K.M.; Thammahong, A.; Barker, B.M.; Rivera, A.; et al. IL-1alpha signaling is critical for leukocyte recruitment after pulmonary Aspergillus fumigatus challenge. PLoS Pathog. 2015, 11, e1004625. [Google Scholar] [CrossRef]
- Gresnigt, M.S.; Rekiki, A.; Rasid, O.; Savers, A.; Jouvion, G.; Dannaoui, E.; Parlato, M.; Fitting, C.; Brock, M.; Cavaillon, J.M.; et al. Reducing hypoxia and inflammation during invasive pulmonary aspergillosis by targeting the Interleukin-1 receptor. Sci. Rep. 2016, 6, 26490. [Google Scholar] [CrossRef]
- Zelante, T.; Bozza, S.; De Luca, A.; D’Angelo, C.; Bonifazi, P.; Moretti, S.; Giovannini, G.; Bistoni, F.; Romani, L. Th17 cells in the setting of Aspergillus infection and pathology. Med. Mycol. 2009, 47 (Suppl. S1), S162–S169. [Google Scholar] [CrossRef]
- Mircescu, M.M.; Lipuma, L.; van Rooijen, N.; Pamer, E.G.; Hohl, T.M. Essential role for neutrophils but not alveolar macrophages at early time points following Aspergillus fumigatus infection. J. Infect. Dis. 2009, 200, 647–656. [Google Scholar] [CrossRef]
- Gazendam, R.P.; van Hamme, J.L.; Tool, A.T.; Hoogenboezem, M.; van den Berg, J.M.; Prins, J.M.; Vitkov, L.; van de Veerdonk, F.L.; van den Berg, T.K.; Roos, D.; et al. Human Neutrophils Use Different Mechanisms To Kill Aspergillus fumigatus Conidia and Hyphae: Evidence from Phagocyte Defects. J. Immunol. 2016, 196, 1272–1283. [Google Scholar] [CrossRef]
- Brown, G.D. Innate antifungal immunity: The key role of phagocytes. Annu. Rev. Immunol. 2011, 29, 1–21. [Google Scholar] [CrossRef]
- Washburn, R.G.; Gallin, J.I.; Bennett, J.E. Oxidative killing of Aspergillus fumigatus proceeds by parallel myeloperoxidase-dependent and -independent pathways. Infect. Immun. 1987, 55, 2088–2092. [Google Scholar] [CrossRef]
- Yipp, B.G.; Kubes, P. NETosis: How vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef]
- Bagnato, G.; Harari, S. Cellular interactions in the pathogenesis of interstitial lung diseases. Eur. Respir. Rev. 2015, 24, 102–114. [Google Scholar] [CrossRef]
- Prufer, S.; Weber, M.; Stein, P.; Bosmann, M.; Stassen, M.; Kreft, A.; Schild, H.; Radsak, M.P. Oxidative burst and neutrophil elastase contribute to clearance of Aspergillus fumigatus pneumonia in mice. Immunobiology 2014, 219, 87–96. [Google Scholar] [CrossRef]
- Zarember, K.A.; Sugui, J.A.; Chang, Y.C.; Kwon-Chung, K.J.; Gallin, J.I. Human polymorphonuclear leukocytes inhibit Aspergillus fumigatus conidial growth by lactoferrin-mediated iron depletion. J. Immunol. 2007, 178, 6367–6373. [Google Scholar] [CrossRef]
- Xiao, X.; Yeoh, B.S.; Vijay-Kumar, M. Lipocalin 2: An Emerging Player in Iron Homeostasis and Inflammation. Annu. Rev. Nutr. 2017, 37, 103–130. [Google Scholar] [CrossRef]
- Happacher, I.; Aguiar, M.; Alilou, M.; Abt, B.; Baltussen, T.J.H.; Decristoforo, C.; Melchers, W.J.G.; Haas, H. The Siderophore Ferricrocin Mediates Iron Acquisition in Aspergillus fumigatus. Microbiol. Spectr. 2023, 11, e0049623. [Google Scholar] [CrossRef]
- Bellocchio, S.; Montagnoli, C.; Bozza, S.; Gaziano, R.; Rossi, G.; Mambula, S.S.; Vecchi, A.; Mantovani, A.; Levitz, S.M.; Romani, L. The contribution of the Toll-like/IL-1 receptor superfamily to innate and adaptive immunity to fungal pathogens in vivo. J. Immunol. 2004, 172, 3059–3069. [Google Scholar] [CrossRef]
- Meier, A.; Kirschning, C.J.; Nikolaus, T.; Wagner, H.; Heesemann, J.; Ebel, F. Toll-like receptor (TLR) 2 and TLR4 are essential for Aspergillus-induced activation of murine macrophages. Cell. Microbiol. 2003, 5, 561–570. [Google Scholar] [CrossRef]
- Chai, L.Y.; Kullberg, B.J.; Vonk, A.G.; Warris, A.; Cambi, A.; Latge, J.P.; Joosten, L.A.; van der Meer, J.W.; Netea, M.G. Modulation of Toll-like receptor 2 (TLR2) and TLR4 responses by Aspergillus fumigatus. Infect. Immun. 2009, 77, 2184–2192. [Google Scholar] [CrossRef]
- Vandamme, D.; Landuyt, B.; Luyten, W.; Schoofs, L. A comprehensive summary of LL-37, the factotum human cathelicidin peptide. Cell. Immunol. 2012, 280, 22–35. [Google Scholar] [CrossRef]
- Liu, P.T.; Stenger, S.; Li, H.; Wenzel, L.; Tan, B.H.; Krutzik, S.R.; Ochoa, M.T.; Schauber, J.; Wu, K.; Meinken, C.; et al. Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science 2006, 311, 1770–1773. [Google Scholar] [CrossRef]
- Luo, X.L.; Li, J.X.; Huang, H.R.; Duan, J.L.; Dai, R.X.; Tao, R.J.; Yang, L.; Hou, J.Y.; Jia, X.M.; Xu, J.F. LL37 Inhibits Aspergillus fumigatus Infection via Directly Binding to the Fungus and Preventing Excessive Inflammation. Front. Immunol. 2019, 10, 283. [Google Scholar] [CrossRef]
- Machata, S.; Muller, M.M.; Lehmann, R.; Sieber, P.; Panagiotou, G.; Carvalho, A.; Cunha, C.; Lagrou, K.; Maertens, J.; Slevogt, H.; et al. Proteome analysis of bronchoalveolar lavage fluids reveals host and fungal proteins highly expressed during invasive pulmonary aspergillosis in mice and humans. Virulence 2020, 11, 1337–1351. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CPA Patients | ILD Patients | |
|---|---|---|
| Subjects (n) | 10 | 18 |
| Male sex (n (%)) | 5 (50%) | 11 (61%) |
| Age—years (mean (min/max)) | 58 (21/83) | 72 (55/86) |
| Identifier | Reactome Pathway | Observed Count | Background Count | Strength | FDR | Gene Name |
|---|---|---|---|---|---|---|
| HSA-6798695 | Neutrophil degranulation | 26 | 476 | 0.87 | 6.73 × 10−12 | CTSG, MPO, AZU1, MMP8, SERPINB10, CEACAM8, BPI, RHOF, CAMP, FCAR, HP, MNDA, S100A8, S100A12, S100A9, MMP9, LCN2, CDA, NHLRC3, SERPINB1, MGAM, CD177, RAB44, SPTAN1, DDX3X, ITGAM |
| HSA-168249 | Innate immune system | 36 | 1041 | 0.67 | 7.07 × 10−12 | CTSG, MPO, AZU1, MMP8, SERPINB10, CEACAM8, BPI, RHOF, NCF1, CAMP, FGB, PSMC5, PRKDC, UBA7, FCAR, HP, MNDA, S100A8, S100A12, S100A9, SFTPD, MMP9, CNPY3, LCN2, CDA, NHLRC3, SERPINB1, DNM2, DHX36, MGAM, CD177, RAB44, SPTAN1, DDX3X, ACTB, ITGAM |
| HSA-166016 | Toll-like receptor 4 (TLR4) cascade | 8 | 139 | 0.9 | 0.0055 | BPI, FGB, S100A8, S100A12, S100A9, SFTPD, DNM2, ITGAM |
| HSA-6799990 | Metal sequestration by antimicrobial proteins | 3 | 6 | 1.84 | 0.0099 | S100A8, S100A9, LCN2 |
| HSA-6803157 | Antimicrobial peptides | 6 | 87 | 0.97 | 0.017 | CTSG, BPI, CAMP, S100A8, S100A9, LCN2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assing, K.; Laursen, C.B.; Campbell, A.J.; Beck, H.C.; Davidsen, J.R. Proteome and Dihydrorhodamine Profiling of Bronchoalveolar Lavage in Patients with Chronic Pulmonary Aspergillosis. J. Fungi 2024, 10, 314. https://doi.org/10.3390/jof10050314
Assing K, Laursen CB, Campbell AJ, Beck HC, Davidsen JR. Proteome and Dihydrorhodamine Profiling of Bronchoalveolar Lavage in Patients with Chronic Pulmonary Aspergillosis. Journal of Fungi. 2024; 10(5):314. https://doi.org/10.3390/jof10050314
Chicago/Turabian StyleAssing, Kristian, Christian B. Laursen, Amanda Jessica Campbell, Hans Christian Beck, and Jesper Rømhild Davidsen. 2024. "Proteome and Dihydrorhodamine Profiling of Bronchoalveolar Lavage in Patients with Chronic Pulmonary Aspergillosis" Journal of Fungi 10, no. 5: 314. https://doi.org/10.3390/jof10050314