
Myco–Phycobiont Interactions within the “Ramalina farinacea Group”: A Geographical Survey over Europe and Macaronesia

 , , , ,
, , , , 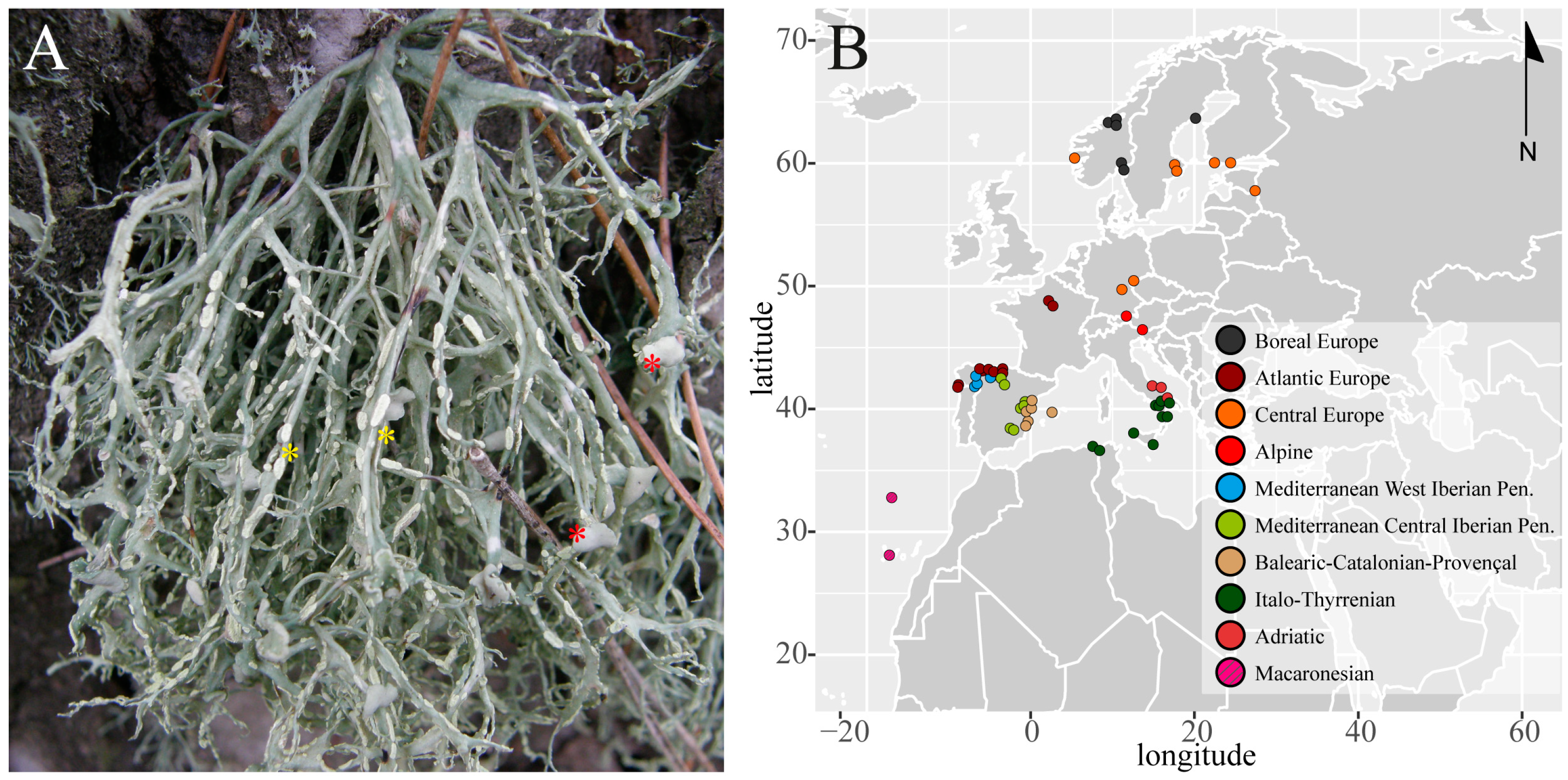{kind=link}
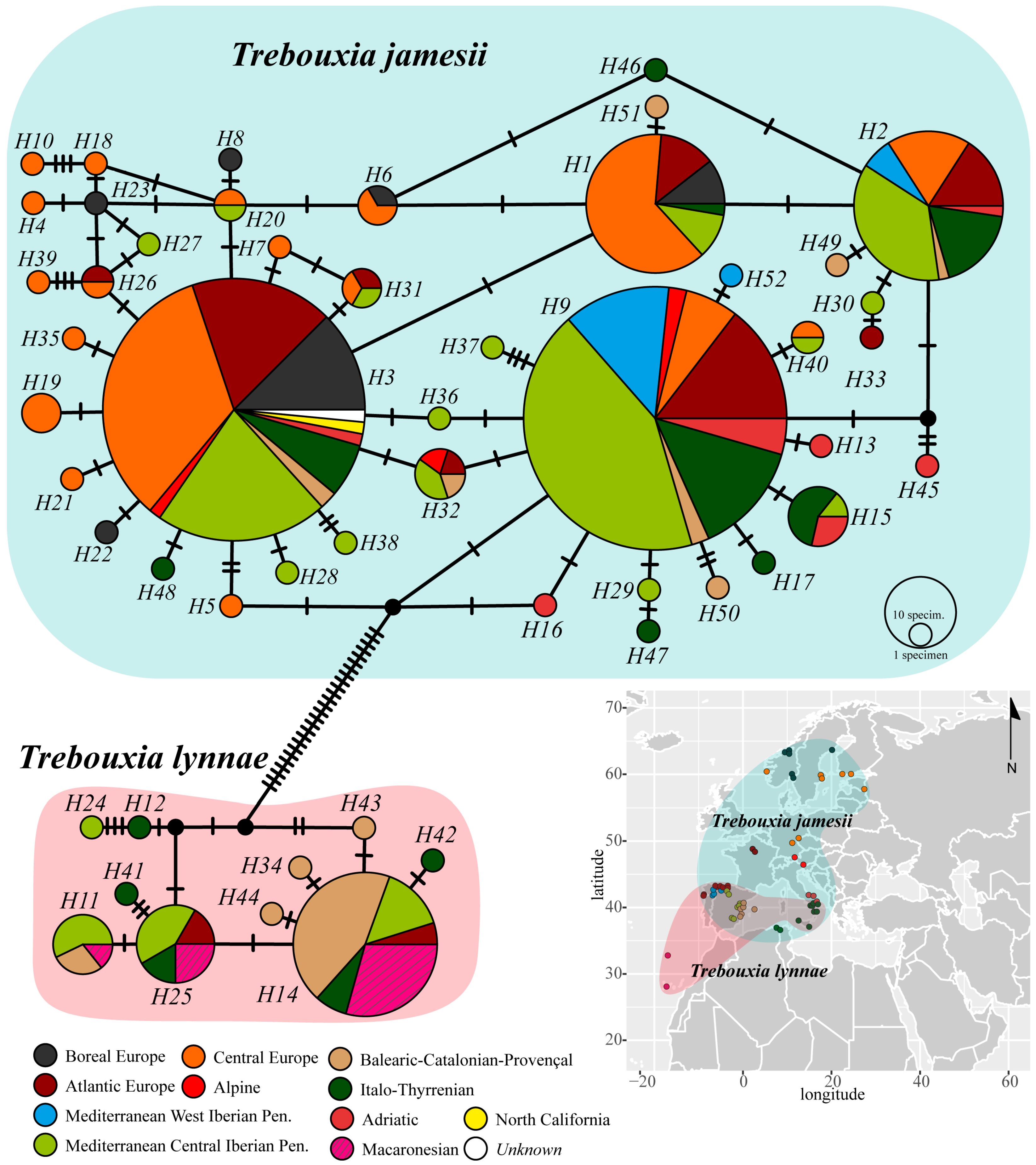{kind=link}
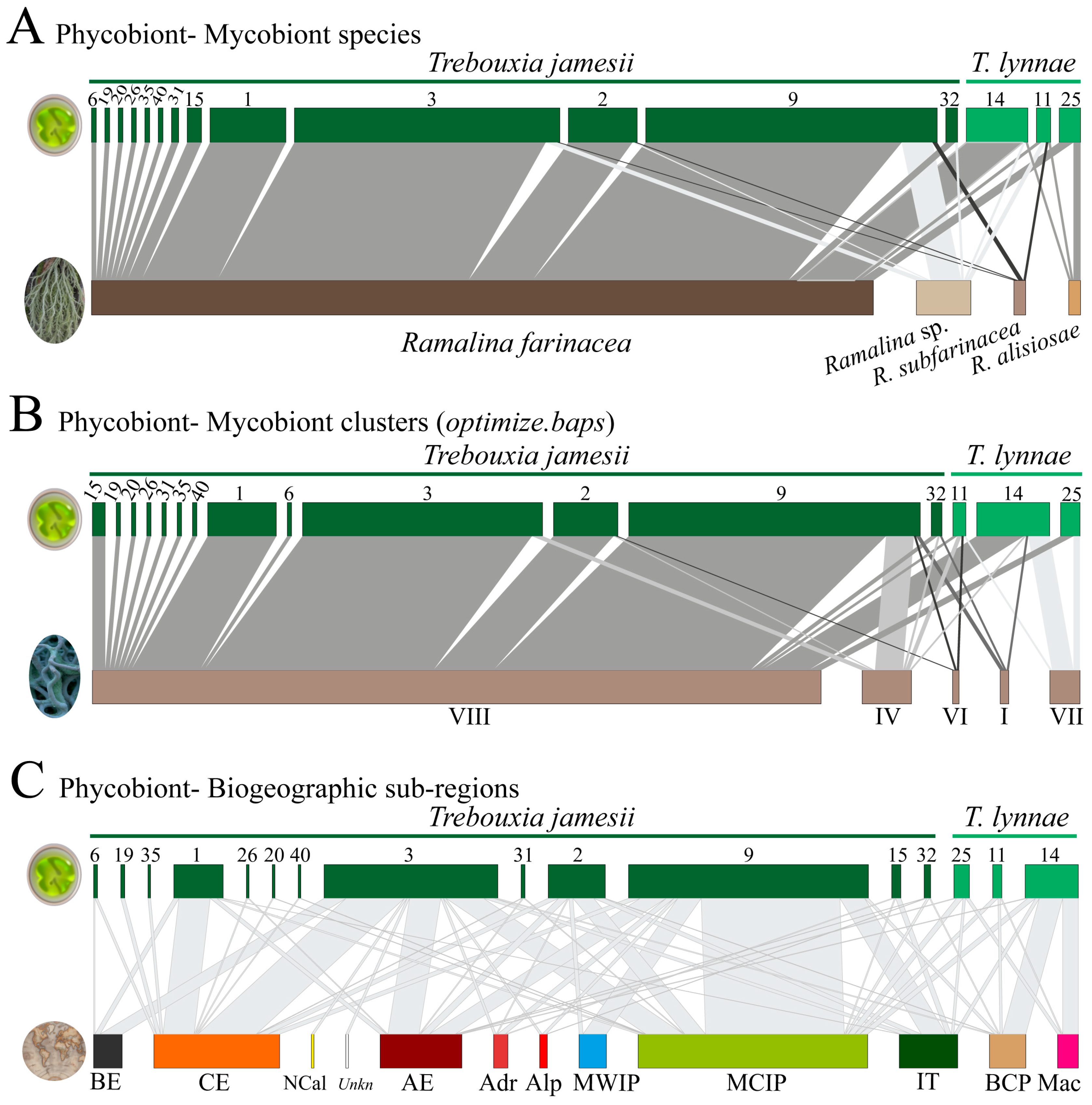{kind=link}
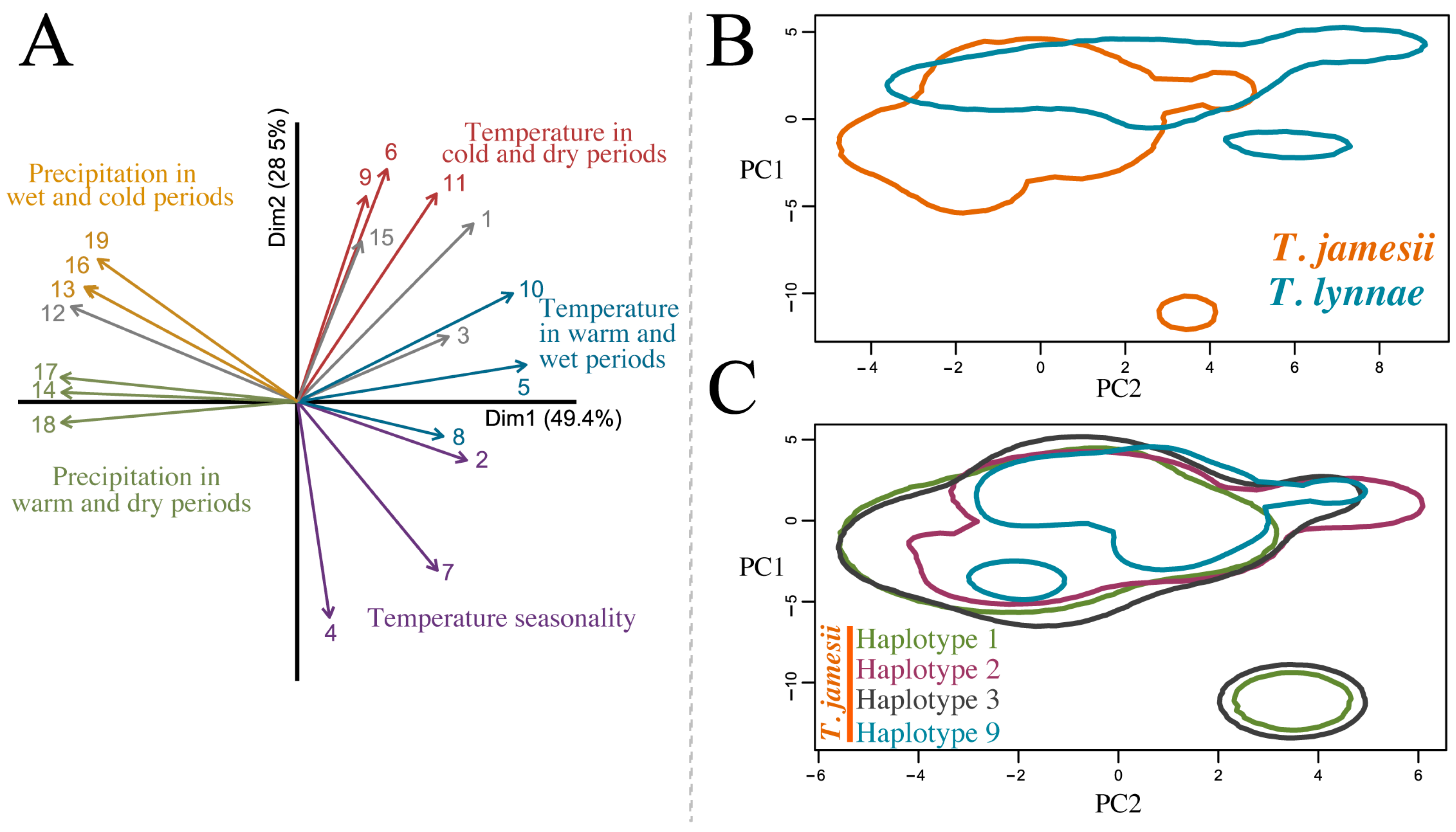{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling, Pretreatment of the Samples, and DNA Extraction
2.2. PCR Amplification and Sequencing
2.3. Inference of Genealogical Relationships among Phycobiont Haplotypes
2.4. Interaction Networks
2.5. Niche Hypervolumes
2.6. Trebouxia jamesii Geographical Occurrence
3. Results
3.1. Phycobiont Haplotype Network
3.2. fastbaps Cluster Assignment and Interaction Networks
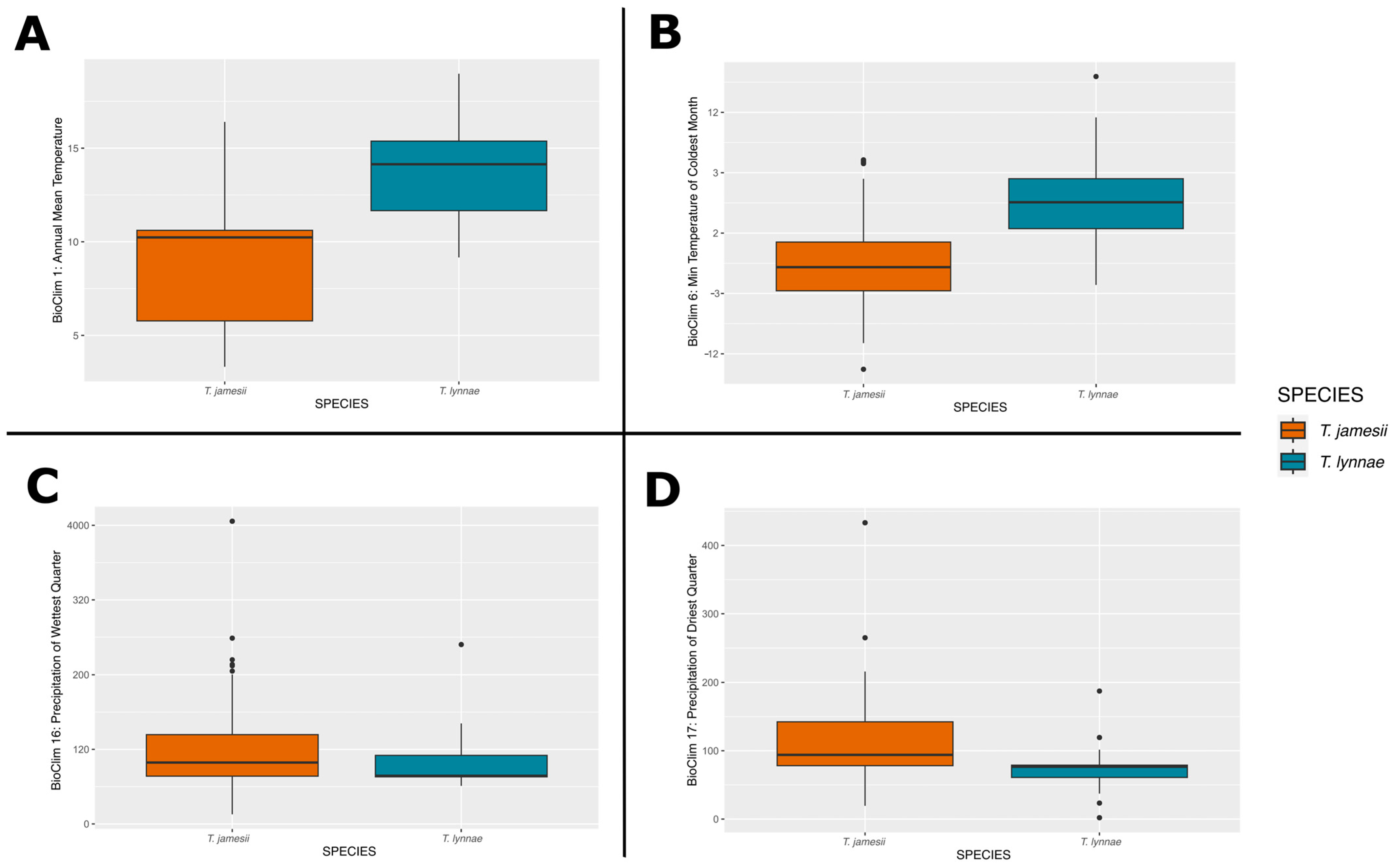
3.3. Climatic Niches
3.4. Phycobiont Geographical Occurrence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Acharius, E. Lichenographia Universalis; Apud Iust. Frid. Danckwerts: Göttingen, Germany, 1810. [Google Scholar]
- Armstrong, R.A. Dispersal of soredia from individual soralia of the lichen Hypogymnia physodes (L.) Nyl. in a simple wind tunnel. Environ. Exp. Bot. 1994, 34, 39–45. [Google Scholar] [CrossRef]
- Walser, J.C.; Zoller, S.; Büchler, U.; Scheidegger, C. Species-specific detection of Lobaria pulmonaria (lichenized ascomycete) diaspores in litter samples trapped in snow cover. Mol. Ecol. 2001, 10, 2129–2138. [Google Scholar] [CrossRef]
- Moya, P.; Garrido-Benavent, I.; Chiva, S.; Pérez-Ortega, S.; Blázquez, M.; Pazos, T.; Hamel, T.; Myllys, L.; Tønsberg, T.; Esseen, P.-A.; et al. Phylogeography of Ramalina farinacea (Lichenized Fungi, Ascomycota) in the Mediterranean Basin, Europe, and Macaronesia. Diversity 2023, 15, 310. [Google Scholar] [CrossRef]
- del Campo, E.; Gimeno, J.; Casano, L.M.; Gasulla, F.; García-Breijo, F.; Reig-Armiñana, J.; Barreno, E. South European populations of Ramalina farinacea (L.) Ach. share different Trebouxia algae. Bibl. Lichenol. 2010, 105, 247–256. [Google Scholar]
- Stocker-Wörgötter, E.; Elix, J.A.; Grube, M. Secondary chemistry of lichen-forming fungi: Chemosyndromic variation and DNA-analyses of cultures and chemotypes in the Ramalina farinacea complex. Bryologist 2004, 107, 152–162. [Google Scholar] [CrossRef]
- Nylander, W. Observata lichenologia in Pyranaeis orientalibus. Bull. Soc. Linnéenne Normandie 1872, 6, 256–328. [Google Scholar]
- Pérez-Vargas, I.; Pérez-Ortega, S. A new endemic Ramalina species from the Canary Islands (Ascomycota, Lecanorales). Phytotaxa 2014, 159, 269–278. [Google Scholar] [CrossRef]
- Krog, H.; Østhagen, H. The genus Ramalina in the Canary Islands. Norw. J. Bot. 1980, 27, 255–296. [Google Scholar]
- Aptroot, A.; Schumm, F. Key to Ramalina species known from Atlantic islands, with two new species from the Azores. Sauteria 2008, 15, 21–57. [Google Scholar]
- del Campo, E.M.; Català, S.; Casano, L.M.; Gimeno, J.; del Hoyo, A.; Martínez-Alberola, F.; Casano, L.M.; Grube, M.; Barreno, E. The genetic structure of the cosmopolitan three-partner lichen Ramalina farinacea evidences the concerted diversification of symbionts. FEMS Microbiol. Ecol. 2013, 83, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Molins, A.; Moya, P.; Muggia, L.; Barreno, E. Thallus growth stage and geographic origin shape microalgal diversity in Ramalina farinacea lichen holobionts. J. Phycol. 2021, 57, 975–987. [Google Scholar] [CrossRef]
- Moya, P.; Molins, A.; Martínez-Alberola, F.; Muggia, L.; Barreno, E. Unexpected associated microalgal diversity in the lichen Ramalina farinacea is uncovered by pyrosequencing analyses. PLoS ONE 2017, 12, e0175091. [Google Scholar] [CrossRef]
- Casano, L.M.; del Campo, E.M.; García-Breijo, F.J.; Reig-Armiñana, J.; Gasulla, F.; Del Hoyo, A.; Guéra, A.; Barreno, E. Two Trebouxia algae with different physiological performances are ever-present in lichen thalli of Ramalina farinacea. Coexistence versus competition? Environ. Microbiol. 2011, 13, 806–818. [Google Scholar] [CrossRef]
- del Hoyo, A.; Álvarez, R.; del Campo, E.; Gasulla, F.; Barreno, E.; Casano, L.M. Oxidative stress induces distinct physiological responses in the two Trebouxia phycobionts of the lichen Ramalina farinacea. Ann. Bot. 2011, 107, 109–118. [Google Scholar] [CrossRef]
- Muggia, L.; Vancurová, L.; Škaloud, P.; Peksa, O.; Wedin, M.; Grube, M. The symbiotic playground of lichen thalli–a highly flexible photobiont association in rock-inhabiting lichens. FEMS Microbiol. Ecol. 2013, 85, 313–323. [Google Scholar] [CrossRef]
- Ohmura, Y.; Takeshita, S.; Kawachi, M. Photobiont diversity within populations of a vegetatively reproducing lichen, Parmotrema tinctorum, can be generated by photobiont switching. Symbiosis 2019, 77, 59–72. [Google Scholar] [CrossRef]
- Peksa, O.; Škaloud, P. Do photobionts influence the ecology of lichens? A case study of environmental preferences in symbiotic green alga Asterochloris (Trebouxiophyceae). Mol. Ecol. 2011, 20, 3936–3948. [Google Scholar] [CrossRef]
- Muggia, L.; Pérez-Ortega, S.; Fryday, A.; Spribille, T.; Grube, M. Global assessment of genetic variation and phenotypic plasticity in the lichen-forming species Tephromela atra. Fungal Divers. 2014, 64, 233–251. [Google Scholar] [CrossRef]
- Dal Grande, F.; Rolshausen, G.; Divakar, P.K.; Crespo, A.; Otte, J.; Schleuning, M.; Schmitt, I. Environment and host identity structure communities of green algal symbionts in lichens. New Phytol. 2018, 217, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Vančurová, L.; Muggia, L.; Peksa, O.; Řídká, T.; Škaloud, P. The complexity of symbiotic interactions influences the ecological amplitude of the host: A case study in Stereocaulon (lichenized ascomycota). Mol. Ecol. 2018, 27, 3016–3033. [Google Scholar] [CrossRef] [PubMed]
- Blázquez, M.; Hernández-Moreno, L.S.; Gasulla, F.; Pérez-Vargas, I.; Pérez-Ortega, S. The role of photobionts as drivers of diversification in an island radiation of lichen-forming fungi. Front. Microbiol. 2022, 12, 784182. [Google Scholar] [CrossRef] [PubMed]
- Škvorová, Z.; Černajová, I.; Steinová, J.; Peksa, O.; Moya, P.; Škaloud, P. Promiscuity in lichens follows clear rules: Partner switching in Cladonia is regulated by climatic factors and soil chemistry. Front. Microbiol. 2022, 12, 781585. [Google Scholar] [CrossRef] [PubMed]
- Steinová, J.; Holien, H.; Košuthová, A.; Škaloud, P. An exception to the rule? Could photobiont identity be a better predictor of lichen phenotype than mycobiont identity? J. Fungi 2022, 8, 275. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Benavent, I.; Mora-Rodríguez, M.R.; Chiva, S.; Fos, S.; Barreno, E. Punctelia borreri and P. subrudecta (Parmeliaceae) associate with a partially overlapping pool of Trebouxia gelatinosa lineages. Lichenologist 2023, 55, 389–399. [Google Scholar] [CrossRef]
- Rodríguez-Arribas, C.; Martínez, I.; Aragón, G.; Zamorano-Elgueta, C.; Cavieres, L.; Prieto, M. Specialization patterns in symbiotic associations: A community perspective over spatial scales. Ecol. Evol. 2023, 13, e10296. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Martínez, S.; Penas, Á.; del Río, S.; Díaz González, T.E.; Rivas-Sáenz, S. Bioclimatology of the Iberian Peninsula and the Balearic Islands. In The Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Springer: Cham, Switzerland, 2017; Volume I, pp. 29–80. [Google Scholar]
- Wickham, H. Data analysis. In ggplot2; Springer: Cham, Switzerland, 2016; pp. 189–201. [Google Scholar]
- Piercey-Normore, M.D.; DePriest, P.T. Algal switching among lichen symbioses. Am. J. Bot. 2001, 88, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Kroken, S.; Taylor, J.W. Phylogenetic species, reproductive mode, and specificity of the green alga Trebouxia forming lichens with the fungal genus Letharia. Bryologist 2000, 103, 645–660. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Larena, I.; Salazar, O.; González, V.; Julián, M.C.; Rubio, V. Design of a primer for ribosomal DNA internal transcribed spacer with enhanced specificity for ascomycetes. J. Biotechnol. 1999, 75, 187–194. [Google Scholar] [CrossRef]
- Werth, S.; Sork, V.L. Ecological specialization in Trebouxia (Trebouxiophyceae) photobionts of Ramalina menziesii (Ramalinaceae) across six range-covering ecoregions of western North America. Am. J. Bot. 2014, 101, 1127–1140. [Google Scholar] [CrossRef]
- Álvarez, R.; Del Hoyo, A.; Díaz-Rodríguez, C.; Coello, A.J.; del Campo, E.M.; Barreno, E.; Casano, L.M. Lichen rehydration in heavy metal-polluted environments: Pb modulates the oxidative response of both Ramalina farinacea thalli and its isolated microalgae. Microbial. Ecol. 2015, 69, 698–709. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Kukwa, M.; Dal Grande, F.; Łubek, A.; Otte, J.; Schmitt, I. A glimpse into genetic diversity and symbiont interaction patterns in lichen communities from areas with different disturbance histories in Białowieża forest, Poland. Microorganisms 2019, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. Mafft multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. PopART: Full feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [CrossRef]
- Clement, M.; Snell, Q.; Walker, P.; Posada, D.; Crandall, K. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1660. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Zhu, C. Coding and applying gaps in phylogenetic analyses. Acta Zootax Sin. 2007, 32, 1–5. [Google Scholar]
- Joly, S.; Stevens, M.I.; van Vuuren, B.J. Haplotype Networks Can Be Misleading in the Presence of Missing Data. Syst. Biol. 2007, 56, 857–862. [Google Scholar] [CrossRef]
- Tonkin-Hill, G.; Lees, J.A.; Bentley, S.D.; Frost, S.D.; Corander, J. Fast hierarchical Bayesian analysis of population structure. Nucleic Acids Res. 2019, 47, 5539–5549. [Google Scholar] [CrossRef]
- R Studio Team. R Studio: Integrated Development for R; R Studio; PBC: Boston, MA, USA, 2023; Available online: http://www.rstudio.com/ (accessed on 17 January 2024).
- Heller, K.A.; Ghahramani, Z. Bayesian Hierarchical Clustering. In Proceedings of the 22nd International Conference on Machine Learning, ICML ’05, Bonn, Germany, 7–11 August 2005; ACM: New York, NY, USA, 2005; pp. 297–304. [Google Scholar]
- Cheng, L.; Connor, T.R.; Sirén, J.; Aanensen, D.M.; Corander, J. Hierarchical and spatially explicit clustering of DNA sequences with BAPS software. Mol. Biol. Ecol. 2013, 30, 1224–1228. [Google Scholar] [CrossRef] [PubMed]
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the bipartite package: Analysing ecological networks. R News 2008, 8, 8–11. [Google Scholar]
- Hutchinson, G.E. Concluding remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Blonder, B.; Lamanna, C.; Violle, C.; Enquist, B.J. The n-dimensional hypervolume. Glob. Ecol. Biogeogr. 2014, 23, 595–609. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef] [PubMed]
- Pateiro-Lopez, B.; Rodriguez-Casal, A. Alphahull: Generalization of the Convex Hull of a Sample of Points in the Plane Manual. 2016. Available online: https://cran.r-project.org/package=alphahull (accessed on 13 January 2024).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid boostrap algorithm for the RAxML web server. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: A community resource for phylogenetic analyses. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Helms, G.; Friedl, T.; Rambold, G.; Mayrhofer, H. Identification of photobionts from the lichen family Physciaceae using algal-specific ITS rDNA sequencing. Lichenologist 2001, 33, 73–86. [Google Scholar] [CrossRef]
- Cordeiro, L.M.; Reis, R.A.; Cruz, L.M.; Stocker-Wörgötter, E.; Grube, M.; Iacomini, M. Molecular studies of photobionts of selected lichens from the coastal vegetation of Brazil. FEMS Microbiol. Ecol. 2005, 54, 381–390. [Google Scholar] [CrossRef]
- Blaha, J.; Baloch, E.; Grube, M. High photobiont diversity associated with the euryoecious lichen-forming ascomycete Lecanora rupicola (Lecanoraceae, Ascomycota). Biol. J. Linnean Soc. 2006, 88, 283–293. [Google Scholar] [CrossRef]
- Schmull, M.; Miadlikowska, J.; Pelzer, M.; Stocker-Wörgötter, E.; Hofstetter, V.; Fraker, E.; Hodkinson, B.P.; Reeb, V.; Kukwa, M.; Lumbsch, H.T.; et al. Phylogenetic affiliations of members of the heterogeneous lichen-forming fungi of the genus Lecidea sensu Zahlbruckner (Lecanoromycetes, Ascomycota). Mycologia 2011, 103, 983–1003. [Google Scholar] [CrossRef] [PubMed]
- Muggia, L.; Pérez-Ortega, S.; Kopun, T.; Zellnig, G.; Grube, M. Photobiont selectivity leads to ecological tolerance and evolutionary divergence in a polymorphic complex of lichenized fungi. Ann. Bot. 2014, 114, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Catalá, S.; Del Campo, E.M.; Barreno, E.; García-Breijo, F.J.; Reig-Armiñana, J.; Casano, L.M. Coordinated ultrastructural and phylogenomic analyses shed light on the hidden phycobiont diversity of Trebouxia microalgae in Ramalina fraxinea. Mol. Phyl. Evol. 2016, 94, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Voytsekhovich, A.; Beck, A. Lichen photobionts of the rocky outcrops of Karadag massif (Crimean Peninsula). Symbiosis 2016, 68, 9–24. [Google Scholar] [CrossRef]
- Singh, G.; Dal Grande, F.; Divakar, P.K.; Otte, J.; Crespo, A.; Schmitt, I. Fungal–algal association patterns in lichen symbiosis linked to macroclimate. New Phytol. 2017, 214, 317–329. [Google Scholar] [CrossRef]
- Ertz, D.; Guzow-Krzemińska, B.; Thor, G.; Łubek, A.; Kukwa, M. Photobiont switching causes changes in the reproduction strategy and phenotypic dimorphism in the Arthoniomycetes. Sci. Rep. 2018, 8, 4952. [Google Scholar] [CrossRef]
- Mark, K.; Laanisto, L.; Bueno, C.G.; Niinemets, Ü.; Keller, C.; Scheidegger, C. Contrasting co-occurrence patterns of photobiont and cystobasidiomycete yeast associated with common epiphytic lichen species. New Phytol. 2020, 227, 1362–1375. [Google Scholar] [CrossRef]
- Bordenave, C.D.; Muggia, L.; Chiva, S.; Leavitt, S.D.; Carrasco, P.; Barreno, E. Chloroplast morphology and pyrenoid ultrastructural analyses reappraise the diversity of the lichen phycobiont genus Trebouxia (Chlorophyta). Algal. Res. 2022, 61, 102561. [Google Scholar] [CrossRef]
- Barreno, E.; Muggia, L.; Chiva, S.; Molins, A.; Bordenave, C.; García-Breijo, F.; Moya, P. Trebouxia lynnae sp. nov. (Former Trebouxia sp. TR9): Biology and Biogeography of an Epitome Lichen Symbiotic Microalga. Biology 2022, 11, 1196. [Google Scholar] [CrossRef]
- Guerrero, R.; Margulis, L.; Berlanga, M. Symbiogenesis: The Holobiont as a Unit of Evolution. Int. Microbiol. 2013, 16, 133–143. [Google Scholar]
- Hawksworth, D.L.; Grube, M. Lichens redefined as complex ecosystems. New Phytol. 2020, 227, 1281–1283. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.L.; Lendemer, J.C. A call to reconceptualize lichen symbioses. Trends Ecol. Evol. 2022, 37, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Kranner, I.; Pichler, G.; Grube, M. The lichen market place. New Phytol. 2022, 234, 541–543. [Google Scholar] [CrossRef]
- Grube, M. Lichens. In Fungal Associations. The Mycota; Hsueh, Y.P., Blackwell, M., Eds.; Springer: Cham, Switzerlands, 2024; Volume 9. [Google Scholar]
- Werth, S. Fungal-algal interactions in Ramalina menziesii and its associated epiphytic lichen community. Lichenologist 2012, 44, 543–560. [Google Scholar] [CrossRef]
- Moya, P.; Škaloud, P.; Chiva, S.; García-Breijo, F.J.; Reig-Arminana, J.; Vančurová, L.; Barreno, E. Molecular phylogeny and ultrastructure of the lichen microalga Asterochloris mediterranea sp. nov. from Mediterranean and Canary Islands ecosystems. Int. J. Syst. Evol. Microbiol. 2015, 65, 1838–1854. [Google Scholar] [CrossRef] [PubMed]
- Moya, P.; Chiva, S.; Molins, A.; Jadrná, I.; Škaloud, P.; Peksa, O.; Barreno, E. Myrmecia israeliensis as the primary symbiotic microalga in squamulose lichens growing in European and Canary Island terricolous communities. Fottea 2018, 18, 72–85. [Google Scholar] [CrossRef]
- Moya, P.; Molins, A.; Chiva, S.; Bastida, J.; Barreno, E. Symbiotic microalgal diversity within lichenicolous lichens and crustose hosts on Iberian Peninsula gypsum biocrusts. Sci. Rep. 2020, 10, 14060. [Google Scholar] [CrossRef]
- Moya, P.; Molins, A.; Škaloud, P.; Divakar, P.K.; Chiva, S.; Dumitru, C.; Molina, M.C.; Crespo, A.; Barreno, E. Biodiversity patterns and ecological preferences of the photobionts associated with the lichen-forming genus Parmelia. Front. Microbiol. 2021, 12, 765310. [Google Scholar] [CrossRef]
- Chrismas, N.; Tindall-Jones, B.; Jenkins, H.; Harley, J.; Bird, K.; Cunliffe, M. Metatranscriptomics reveals diversity of symbiotic interaction and mechanisms of carbon exchange in the marine cyanolichen Lichina pygmaea. New Phytol. 2023, 241, 2243–2257. [Google Scholar] [CrossRef]
- Helms, G. Taxonomy and Symbiosis in Associations of Physciaceae and Trebouxia. Ph.D. Thesis, University of Göttingen, Göttingen, Germany, 2003. [Google Scholar]
- Yahr, R.; Vilgalys, R.; Depriest, P.T. Strong fungal specificity and selectivity for algal symbionts in Florida scrub Cladonia lichens. Mol. Ecol. 2004, 13, 3367–3378. [Google Scholar] [CrossRef]
- Castillo, R.V.; Beck, A. Photobiont selectivity and specificity in Caloplaca species in a fog-induced community in the Atacama Desert, northern Chile. Fung Biol. 2012, 116, 665–676. [Google Scholar] [CrossRef]
- Nyati, S.; Scherrer, S.; Werth, S.; Honegger, R. Green-algal photobiont diversity (Trebouxia spp.) in representatives of Teloschistaceae (Lecanoromycetes, lichen-forming ascomycetes). Lichenologist 2014, 46, 189–212. [Google Scholar] [CrossRef]
- Garrido-Benavent, I.; Pérez-Ortega, S.; de los Ríos, A.; Fernández-Mendoza, F. Amphitropical variation of the algal partners of Pseudephebe (Parmeliaceae, lichenized fungi). Symbiosis 2020, 82, 35–48. [Google Scholar] [CrossRef]
- Garrido-Benavent, I.; Molins, A.; Barreno, E. Genetic variation in the symbiont partners in the endangered macrolichen Seirophora villosa (Teloschistaceae: Ascomycota). Bot. J. Linnean Soc. 2022, 199, 816–829. [Google Scholar] [CrossRef]
- Nelsen, M.P.; Gargas, A. Dissociation and horizontal transmission of codispersing lichen symbionts in the genus Lepraria (Lecanorales: Stereocaulaceae). New Phytol. 2008, 177, 264–275. [Google Scholar] [CrossRef]
- Pino-Bodas, R.; Stenroos, S. Global biodiversity patterns of the photobionts associated with the genus Cladonia (Lecanorales, Ascomycota). Microb. Ecol. 2021, 82, 173–187. [Google Scholar] [CrossRef]
- Beck, A. Photobiont inventory of a lichen community growing on heavy-metal-rich rock. Lichenologist 1999, 31, 501–510. [Google Scholar] [CrossRef]
- Beck, A. Selektivität der Symbionten Schwermetalltoleranter Flechten. Ph.D. Thesis, Ludwig-Maximilians-Universität München, Munich, Germany, 2002. [Google Scholar]
- Doering, M.; Piercey-Normore, M.D. Genetically divergent algae shape an epiphytic lichen community on Jack Pine in Manitoba. Lichenologist 2009, 41, 69–80. [Google Scholar] [CrossRef]
- Berlinches de Gea, A.; Verdú, M.; Villar-de Pablo, M.; Pérez-Ortega, S. The combined effects of habitat fragmentation and life history traits on specialisation in lichen symbioses. J. Ecol. 2023, 112, 200–216. [Google Scholar] [CrossRef]
- Hinojosa-Vidal, E.; Marco, F.; Martínez-Alberola, F.; Escaray, F.J.; García-Breijo, F.J.; Reig-Armiñana, J.; Carrasco, P.; Barreno, E. Characterization of the responses to saline stress in the symbiotic green microalga Trebouxia sp. TR9. Planta 2018, 248, 1473–1486. [Google Scholar] [CrossRef]
- Pérez-Rodrigo, M.; Moya, P.; Marco, F.; Carrasco, P.; Barreno, E.A. Symbiotic Trebouxia sp. TR9, Asterochloris erici and free-living Chlorella vulgaris green microalgae respond differentially to osmotic and saline stresses. In Proceedings of the IAL9 Program & Abstracts Book, Universidade Federal de Sergipe, Brasil, International Association for Lichenology 9th Symposium “Unlocking the Inner Lichen” Virtual-IAL9, Bonito, Brazil, 1–6 August 2021. [Google Scholar]
- Fernández-Mendoza, F.; Domaschke, S.; García, M.A.; Jordan, P.; Martín, M.P.; Printzen, C. Population structure of mycobionts and photobionts of the widespread lichen Cetraria aculeata. Mol. Ecol. 2011, 20, 1208–1232. [Google Scholar] [CrossRef]
- Yahr, R.; Vilgalys, R.; DePriest, P.T. Geographic variation in algal partners of Cladonia subtenuis (Cladoniaceae) highlights the dynamic nature of a lichen symbiosis. New Phytol. 2006, 171, 847–860. [Google Scholar] [CrossRef]
- Piercey-Normore, M.D. The lichen-forming ascomycete Evernia mesomorpha associates with multiple genotypes of Trebouxia jamesii. Phytologist 2006, 169, 331–344. [Google Scholar] [CrossRef]
- Dal Grande, F.; Alors, D.; Divakar, P.K.; Bálint, M.; Crespo, A.; Schmitt, I. Insights into intrathalline genetic diversity of the cosmopolitan lichen symbiotic green alga Trebouxia decolorans Ahmadjian using microsatellite markers. Mol. Phyl. Evol. 2014, 72, 54–60. [Google Scholar] [CrossRef]
- Sanders, W.B.; Masumoto, H. Lichen algae: The photosynthetic partners in lichen symbioses. Lichenologist 2021, 53, 347–393. [Google Scholar] [CrossRef]
- Friedl, T. Comparative ultrastructure of pyrenoids in Trebouxia (Microthamniales, Chlorophyta). Plant Syst. Evol. 1989, 164, 145–159. [Google Scholar] [CrossRef]
- Peršoh, D.; Beck, A.; Rambold, G. The distribution of ascus types and photobiontal selection in Lecanoromycetes (Ascomycota) against the background of a revised SSU nrDNA phylogeny. Mycol. Prog. 2004, 3, 103–121. [Google Scholar] [CrossRef]
- Opanowicz, M.; Grube, M. Photobiont genetic variation in Flavocetraria nivalis from Poland (Parmeliaceae, lichenized Ascomycota). Lichenologist 2004, 36, 125–131. [Google Scholar] [CrossRef]
- Muggia, L.; Nelsen, M.P.; Kirika, P.M.; Barreno, E.; Beck, A.; Lindgren, H.; Lumbsch, H.T.; Leavitt, S.D.; Trebouxia Working Group. Formally described species woefully underrepresent phylogenetic diversity in the common lichen photobiont genus Trebouxia (Trebouxiophyceae, Chlorophyta): An impetus for developing an integrated taxonomy. Mol. Phyl. Evol. 2020, 149, 106821. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2024. Available online: https://www.algaebase.org (accessed on 17 January 2024).





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moya, P.; Chiva, S.; Pazos, T.; Barreno, E.; Carrasco, P.; Muggia, L.; Garrido-Benavent, I. Myco–Phycobiont Interactions within the “Ramalina farinacea Group”: A Geographical Survey over Europe and Macaronesia. J. Fungi 2024, 10, 206. https://doi.org/10.3390/jof10030206
Moya P, Chiva S, Pazos T, Barreno E, Carrasco P, Muggia L, Garrido-Benavent I. Myco–Phycobiont Interactions within the “Ramalina farinacea Group”: A Geographical Survey over Europe and Macaronesia. Journal of Fungi. 2024; 10(3):206. https://doi.org/10.3390/jof10030206
Chicago/Turabian StyleMoya, Patricia, Salvador Chiva, Tamara Pazos, Eva Barreno, Pedro Carrasco, Lucia Muggia, and Isaac Garrido-Benavent. 2024. "Myco–Phycobiont Interactions within the “Ramalina farinacea Group”: A Geographical Survey over Europe and Macaronesia" Journal of Fungi 10, no. 3: 206. https://doi.org/10.3390/jof10030206