
Exploring Mechanisms of Antifungal Lipopeptide Iturin A from Bacillus against Aspergillus niger

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Microorganisms and Culture Conditions
2.3. Extraction of Iturin A
2.4. Determination of the Inhibitory Effect of Iturin A on A. niger
2.4.1. Determination of Minimum Inhibitory Concentrations
2.4.2. Effect of Iturin A on Spore Germination
2.4.3. Effect of Iturin A on Colony Growth
2.5. Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM) Observations
2.6. Determination of Cytomembrane Damage
2.7. Intracellular Reactive Oxygen Species Accumulation
2.8. Determination of Mitochondrial Membrane Potential
2.9. Determination of Malondialdehyde (MDA) Content
2.10. Determination of Mitochondrial ATPase and Dehydrogenase Activity
2.11. Proteomic Analysis
2.11.1. Protein Extraction
2.11.2. Enzymatic Desalination
2.11.3. LC-MS/MS and Bioinformatic Analysis
2.12. Statistical Analysis
3. Results
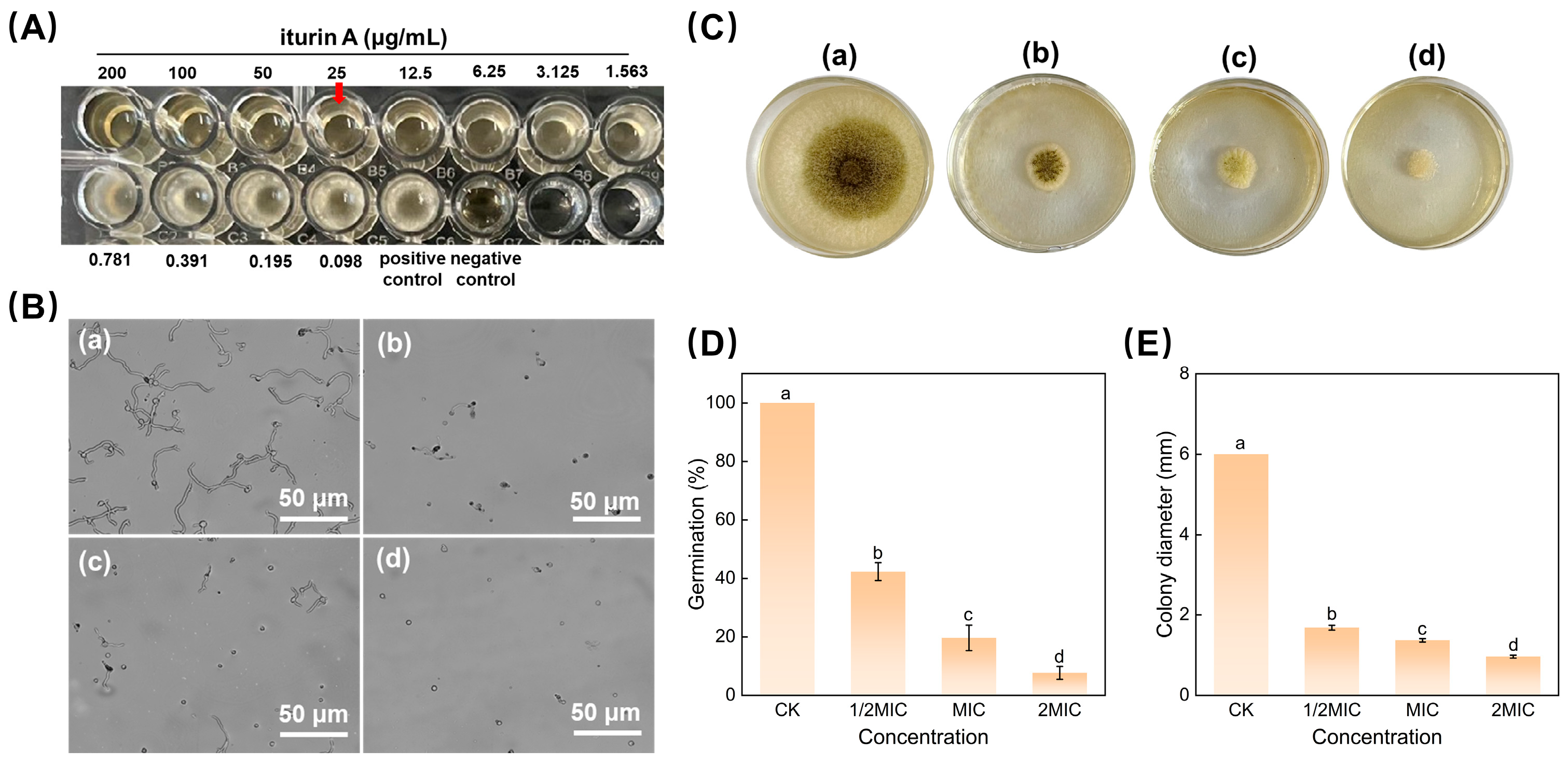
3.1. Inhibitory Effect of Iturin A on A. niger
3.1.1. Effect of iturin A at the Minimum Inhibitory Concentration against A. niger
3.1.2. Spore Germination and Mycelial Growth Assay
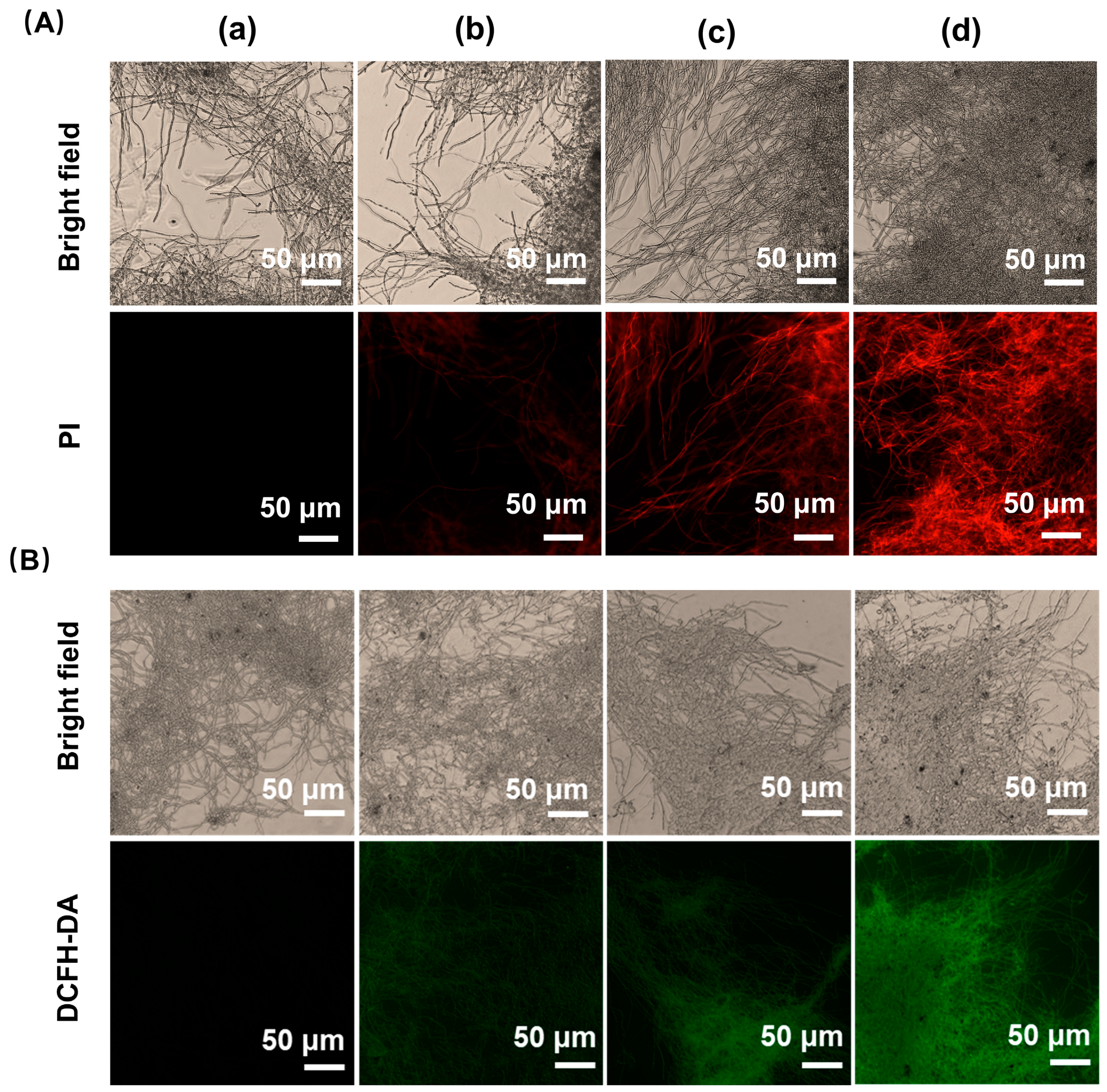
3.2. Effect of Iturin A on the Morphology and Cellular Structure of A. niger
3.3. Effect of Iturin A on Cytomembrane Integrity
3.4. Effect of Iturin A on Intracellular ROS Generation
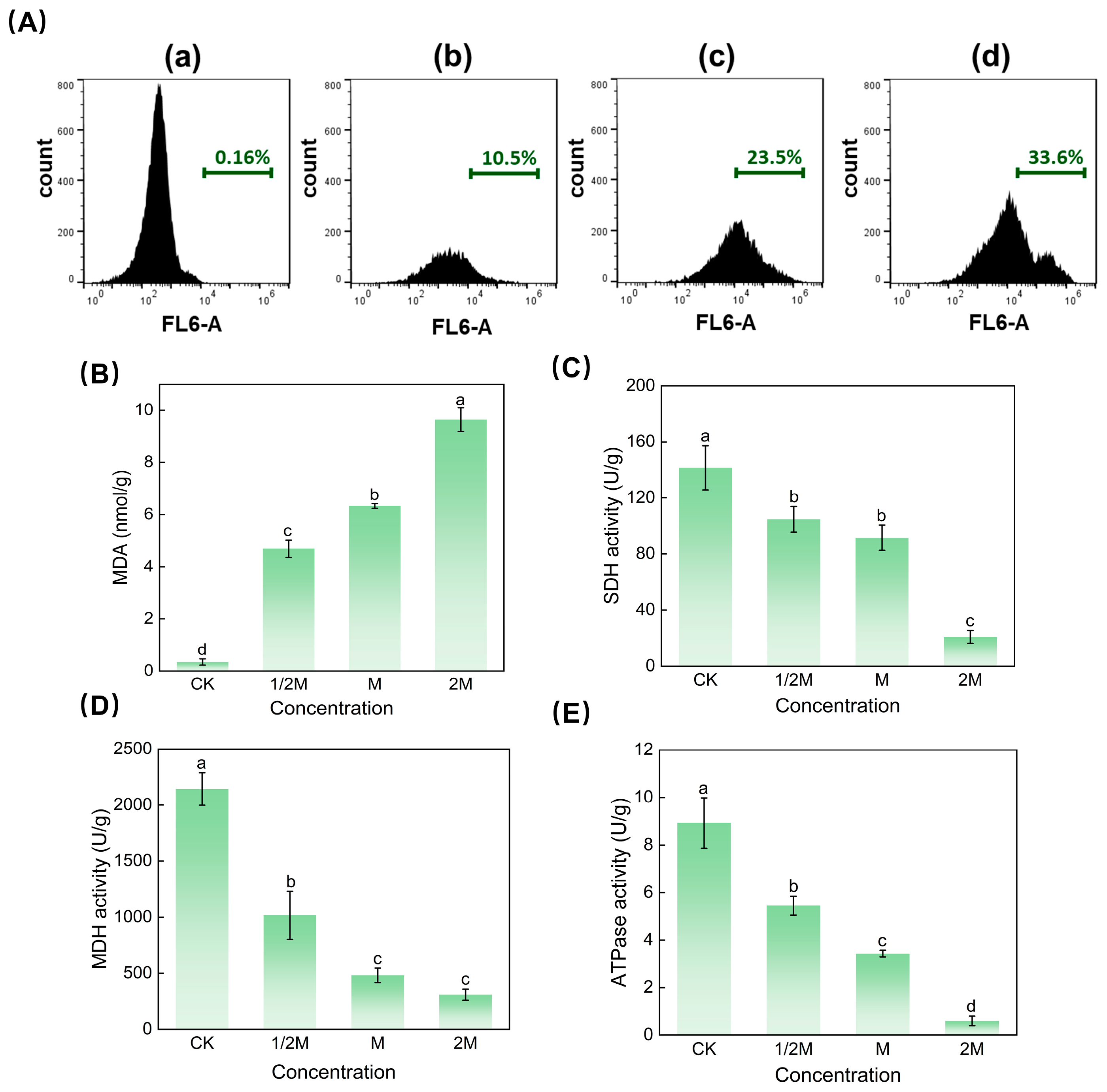
3.5. Effect of Iturin A on the Mitochondrial Membrane Potential
3.6. Effect of Iturin A on MDA Content
3.7. Determination of Mitochondrial Dehydrogenase and ATPase Activity
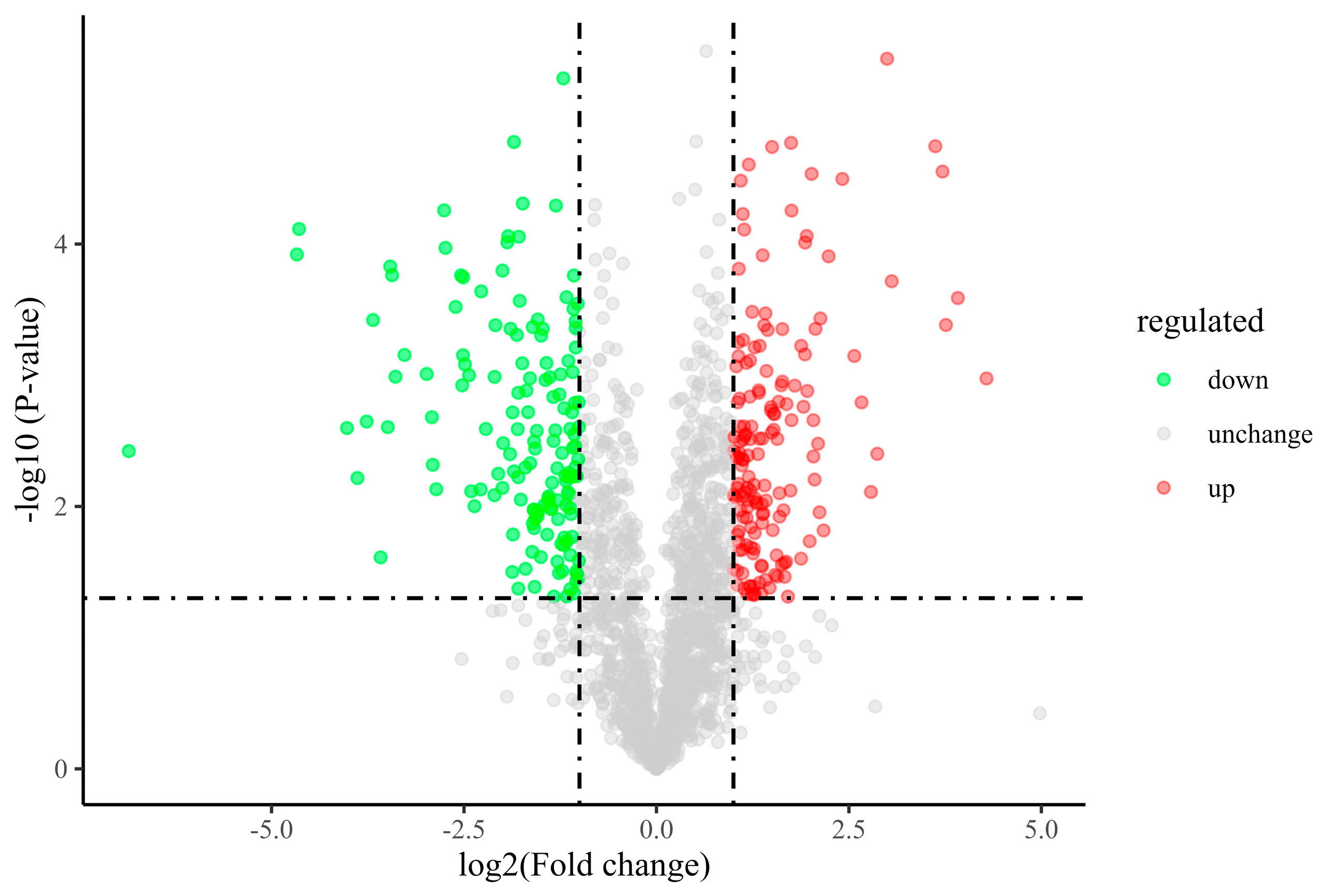
3.8. Comparative Proteomic Analysis of A. niger
3.8.1. Proteomic Profiles
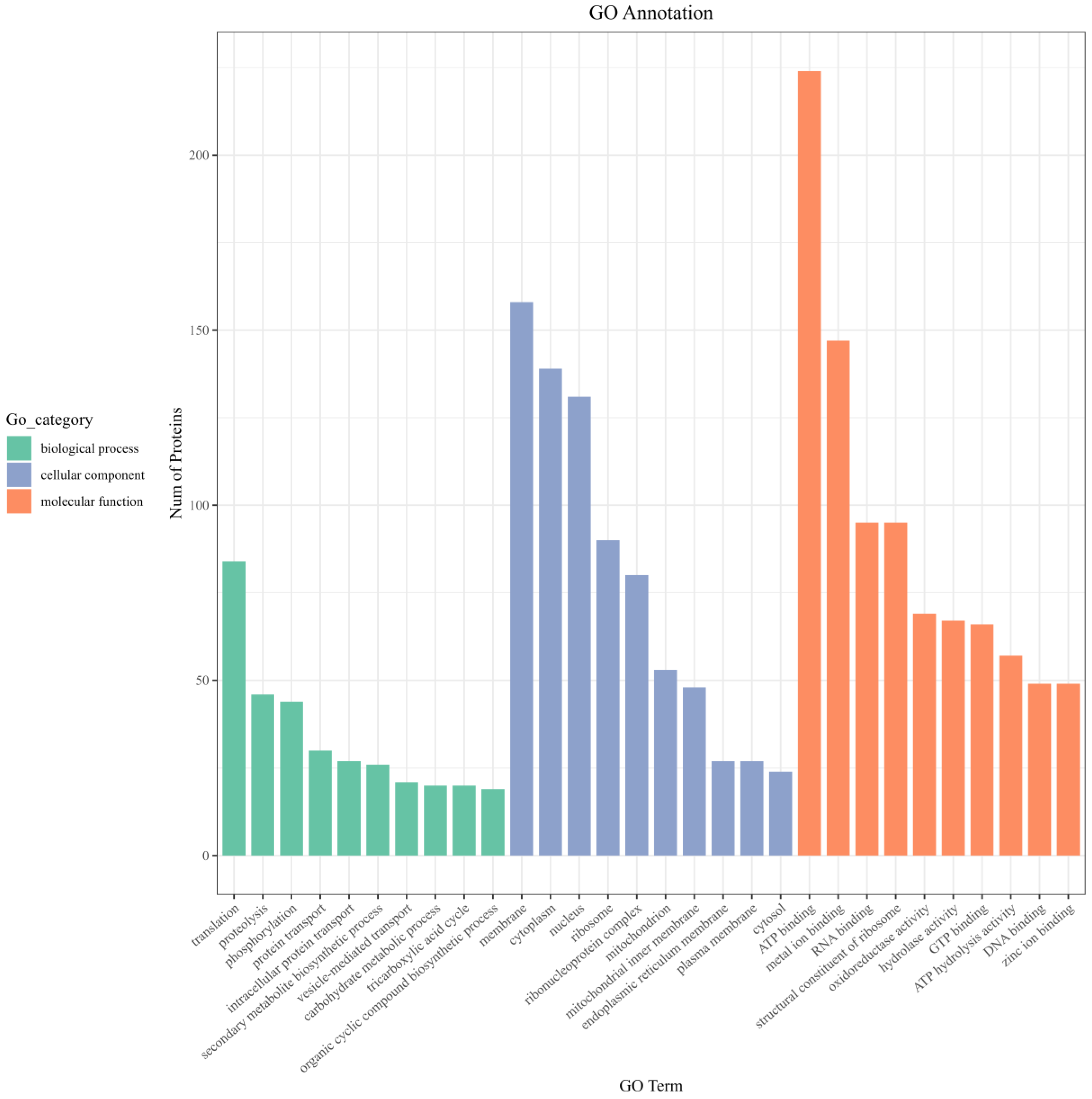
3.8.2. GO Function Enrichment of Differentially Expressed Proteins
3.8.3. KEGG Function Enrichment of Differentially Expressed Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, H.N.; Chaves-Lopez, C.; Oliveira, R.C.; Paparella, A.; Rodrigues, D.F. Cellular and metabolic approaches to investigate the effects of graphene and graphene oxide in the fungi Aspergillus flavus and Aspergillus niger. Carbon 2019, 143, 419–429. [Google Scholar] [CrossRef]
- Ju, J.; Xie, Y.; Yu, H.; Guo, Y.; Cheng, Y.; Zhang, R.; Yao, W. Synergistic inhibition effect of citral and eugenol against Aspergillus niger and their application in bread preservation. Food Chem. 2020, 310, 125974. [Google Scholar] [CrossRef]
- Ju, J.; Xu, X.; Xie, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Inhibitory effects of cinnamon and clove essential oils on mold growth on baked foods. Food Chem. 2018, 240, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.; Lei, Y.; Guo, Y.; Yu, H.; Cheng, Y.; Yao, W. Eugenol and citral kills Aspergillus niger through the tricarboxylic acid cycle and its application in food preservation. LWT 2023, 173, 114226. [Google Scholar] [CrossRef]
- Kumar, P.; Mishra, S.; Kumar, A.; Kumar, S.; Prasad, C.S. In Vivo and In Vitro control activity of plant essential oils against three strains of Aspergillus niger. Environ. Sci. Pollut. Res. Int. 2017, 24, 21948–21959. [Google Scholar] [CrossRef]
- Veras, F.F.; Silveira, R.D.; Welke, J.E. Bacillus spp. as a strategy to control fungi and mycotoxins in food. Curr. Opin. Food Sci. 2023, 52, 101068. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Fan, X.; Lou, Z.; Wang, H.; Olatunde, A.; Rengasamy, K.R.R. Iturin: Cyclic lipopeptide with multifunction biological potential. Crit. Rev. Food Sci. Nutr. 2022, 62, 7976–7988. [Google Scholar] [CrossRef] [PubMed]
- Ambrico, A.; Trupo, M. Efficacy of cell free supernatant from Bacillus subtilis ET-1, an Iturin A producer strain, on biocontrol of green and gray mold. Postharvest Biol. Technol. 2017, 134, 5–10. [Google Scholar] [CrossRef]
- Yan, F.; Li, C.; Ye, X.; Lian, Y.; Wu, Y.; Wang, X. Antifungal activity of lipopeptides from Bacillus amyloliquefaciens MG3 against Colletotrichum gloeosporioides in loquat fruits. Biol. Control 2020, 146, 104281. [Google Scholar] [CrossRef]
- Arrebola, E.; Jacobs, R.; Korsten, L. Iturin A is the principal inhibitor in the biocontrol activity of Bacillus amyloliquefaciens PPCB004 against postharvest fungal pathogens. J. Appl. Microbiol. 2010, 108, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhou, Z.-J.; Han, Y.; Wang, Z.-Z.; Fan, J.; Xiao, H.-Z. Isolation and identification of antifungal peptides from Bacillus BH072, a novel bacterium isolated from honey. Microbiol. Res. 2013, 168, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Li, J.; Sun, Y.; Chen, Q.; Zhang, L.; Le, T. The antifungal effects of cinnamaldehyde against Aspergillus niger and its application in bread preservation. Food Chem. 2020, 317, 126405. [Google Scholar] [CrossRef]
- Ma, Y.-N.; Xu, F.-R.; Chen, C.-J.; Li, Q.-Q.; Wang, M.-Z.; Cheng, Y.-X.; Dong, X. The beneficial use of essential oils from buds and fruit of Syzygium aromaticum to combat pathogenic fungi of Panax notoginseng. Ind. Crops Prod. 2019, 133, 185–192. [Google Scholar] [CrossRef]
- Wang, C.; Yuan, S.; Zhang, W.; Ng, T.B.; Ye, X. Buckwheat Antifungal Protein with Biocontrol Potential To Inhibit Fungal (Botrytis cinerea) Infection of Cherry Tomato. J. Agric. Food Chem. 2019, 67, 6748–6756. [Google Scholar] [CrossRef]
- Li, S.-F.; Zhang, S.-B.; Lv, Y.-Y.; Zhai, H.-C.; Hu, Y.-S.; Cai, J.-P. Heptanal inhibits the growth of Aspergillus flavus through disturbance of plasma membrane integrity, mitochondrial function and antioxidant enzyme activity. LWT 2022, 154, 112655. [Google Scholar] [CrossRef]
- Ke, Y.; Ding, B.; Zhang, M.; Dong, T.; Fu, Y.; Lv, Q.; Ding, W.; Wang, X. Study on inhibitory activity and mechanism of chitosan oligosaccharides on Aspergillus Flavus and Aspergillus Fumigatus. Carbohydr. Polym. 2022, 275, 118673. [Google Scholar] [CrossRef]
- Feng, L.; Li, Y.; Wang, Z.; Qi, L.; Mo, H. Antifungal Actions of Glycinin Basic Peptide against Aspergillus niger through the Collaborative Damage to Cell Membrane and Mitochondria. Food Biophys. 2018, 14, 97–107. [Google Scholar] [CrossRef]
- Chen, J.; Wang, H.; Chen, Y.; Zhu, Q.; Wan, J. Inhibitive effect and mechanism of cinnamaldehyde on growth and OTA production of Aspergillus niger in vitro and in dried red chilies. Food Res. Int. 2023, 168, 112794. [Google Scholar] [CrossRef]
- Yue, Y.; Wang, Z.; Zhong, T.; Guo, M.; Huang, L.; Yang, L.; Kan, J.; Zalán, Z.; Hegyi, F.; Takács, K.; et al. Antifungal mechanisms of volatile organic compounds produced by Pseudomonas fluorescens ZX as biological fumigants against Botrytis cinerea. Microbiol. Res. 2023, 267, 127253. [Google Scholar] [CrossRef]
- Yan, H.; Meng, X.; Lin, X.; Duan, N.; Wang, Z.; Wu, S. Antifungal activity and inhibitory mechanisms of ferulic acid against the growth of Fusarium graminearum. Food Biosci. 2023, 52, 102414. [Google Scholar] [CrossRef]
- Yu, H.; Lin, Z.-X.; Xiang, W.-L.; Huang, M.; Tang, J.; Lu, Y.; Zhao, Q.-H.; Zhang, Q.; Rao, Y.; Liu, L. Antifungal activity and mechanism of D-limonene against foodborne opportunistic pathogen Candida tropicalis. LWT—Food Sci. Technol. 2022, 159, 113144. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef]
- Zaccaria, M.; Ludovici, M.; Sanzani, S.M.; Ippolito, A.; Cigliano, R.A.; Sanseverino, W.; Scarpari, M.; Scala, V.; Fanelli, C.; Reverberi, M. Menadione-Induced Oxidative Stress Re-Shapes the Oxylipin Profile of Aspergillus flavus and Its Lifestyle. Toxins 2015, 7, 4315–4329. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, L.; Du, F.; Sun, L.; Shi, J.; Long, M.; Chen, Z. Activity and Mechanism of Action of Antifungal Peptides from Microorganisms: A Review. Molecules 2021, 26, 3438. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Hammer, J.; Pate, M.; Zhang, Y.; Zhu, F.; Zmuda, E.; Blazyk, J. Antimicrobial activities and structures of two linear cationic peptide families with various amphipathic beta-sheet and alpha-helical potentials. Antimicrob. Agents Chemother. 2005, 49, 4957–4964. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Li, M.; Cao, Y.; Gao, P.; Wang, Y.; Jiang, Y. The alternative oxidase of Candida albicans causes reduced fluconazole susceptibility. J. Antimicrob. Chemother. 2009, 64, 764–773. [Google Scholar] [CrossRef]
- Yaakoub, H.; Mina, S.; Calenda, A.; Bouchara, J.-P.; Papon, N. Oxidative stress response pathways in fungi. Cell. Mol. Life Sci. 2022, 79, 333. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Šileikytė, J.; Forte, M. The Mitochondrial Permeability Transition in Mitochondrial Disorders. Oxidative Med. Cell. Longev. 2019, 2019, 3403075. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Zhang, Y.; Ju, J.; Xie, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Quek, S.Y.; Yao, W. Antifungal effects of thymol and salicylic acid on cell membrane and mitochondria of Rhizopus stolonifer and their application in postharvest preservation of tomatoes. Food Chem. 2019, 285, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Calderone, R. Exploiting mitochondria as targets for the development of new antifungals. Virulence 2016, 8, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Zhao, L.; Zhao, W.; Xie, Y. (E)-2-Hexenal, as a Potential Natural Antifungal Compound, Inhibits Aspergillus flavus Spore Germination by Disrupting Mitochondrial Energy Metabolism. J. Agric. Food Chem. 2019, 67, 1138–1145. [Google Scholar] [CrossRef]
- Deutscher, J.; Aké, F.M.D.; Derkaoui, M.; Zébré, A.C.; Cao, T.N.; Bouraoui, H.; Kentache, T.; Mokhtari, A.; Milohanic, E.; Joyet, P. The Bacterial Phosphoenolpyruvate:Carbohydrate Phosphotransferase System: Regulation by Protein Phosphorylation and Phosphorylation-Dependent Protein-Protein Interactions. Microbiol. Mol. Biol. Rev. 2014, 78, 231–256. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Xu, M.; Han, Y.; Zhou, Z. Exploring Mechanisms of Antifungal Lipopeptide Iturin A from Bacillus against Aspergillus niger. J. Fungi 2024, 10, 172. https://doi.org/10.3390/jof10030172
Wang S, Xu M, Han Y, Zhou Z. Exploring Mechanisms of Antifungal Lipopeptide Iturin A from Bacillus against Aspergillus niger. Journal of Fungi. 2024; 10(3):172. https://doi.org/10.3390/jof10030172
Chicago/Turabian StyleWang, Shiyi, Min Xu, Ye Han, and Zhijiang Zhou. 2024. "Exploring Mechanisms of Antifungal Lipopeptide Iturin A from Bacillus against Aspergillus niger" Journal of Fungi 10, no. 3: 172. https://doi.org/10.3390/jof10030172