
Biodegradable Mulch Films and Bioformulations Based on Trichoderma sp. and Seaweed Extract Differentially Affect the Metabolome of Industrial Tomato Plants

 , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Pre-Transplant Processing
2.2. Biostimulants
- (i)
- Phylgreen® (Trade Corporation International, Madrid, Spain), a liquid formulation consisting of the pure extract of the seaweed Ascophyllum nodosum. The formulation is soluble and has a high content of alginates, vitamins, natural antioxidants and noble amino acids; it is used to improve flowering and fruit setting, acts as a promoter of photosynthesis and root development, and increases plant tolerance to environmental stress.
- (ii)
- Trianum-P® (Koppert B. V., Berkel en Rodenrijs, The Netherlands), containing the microorganism Trichoderma afroharzianum (ex T. harzianum) Rifai strain KRL-AG2 (T-22). The bioformulation contains a minimum concentration of 1 × 109 CFU g−1. It has been registered as a Plant Protection Product (PPP), but it is also able to stimulate plant growth and development.
2.3. Experimental Design and Field Management
2.4. Plant Sampling
2.5. Metabolites Extraction
2.6. LC-MS Analysis
2.7. Statistical Analysis
3. Results
3.1. Metabolites Identification
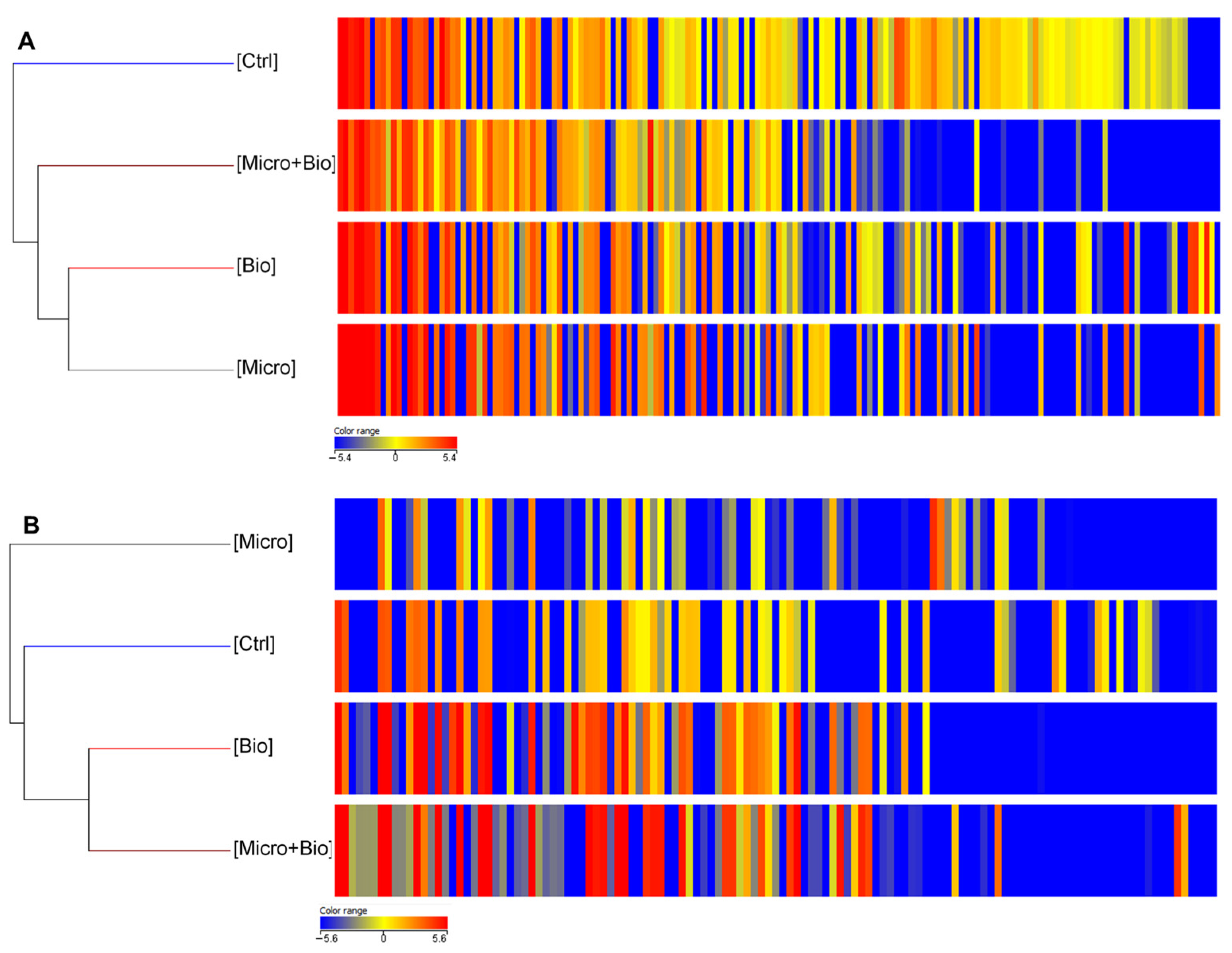
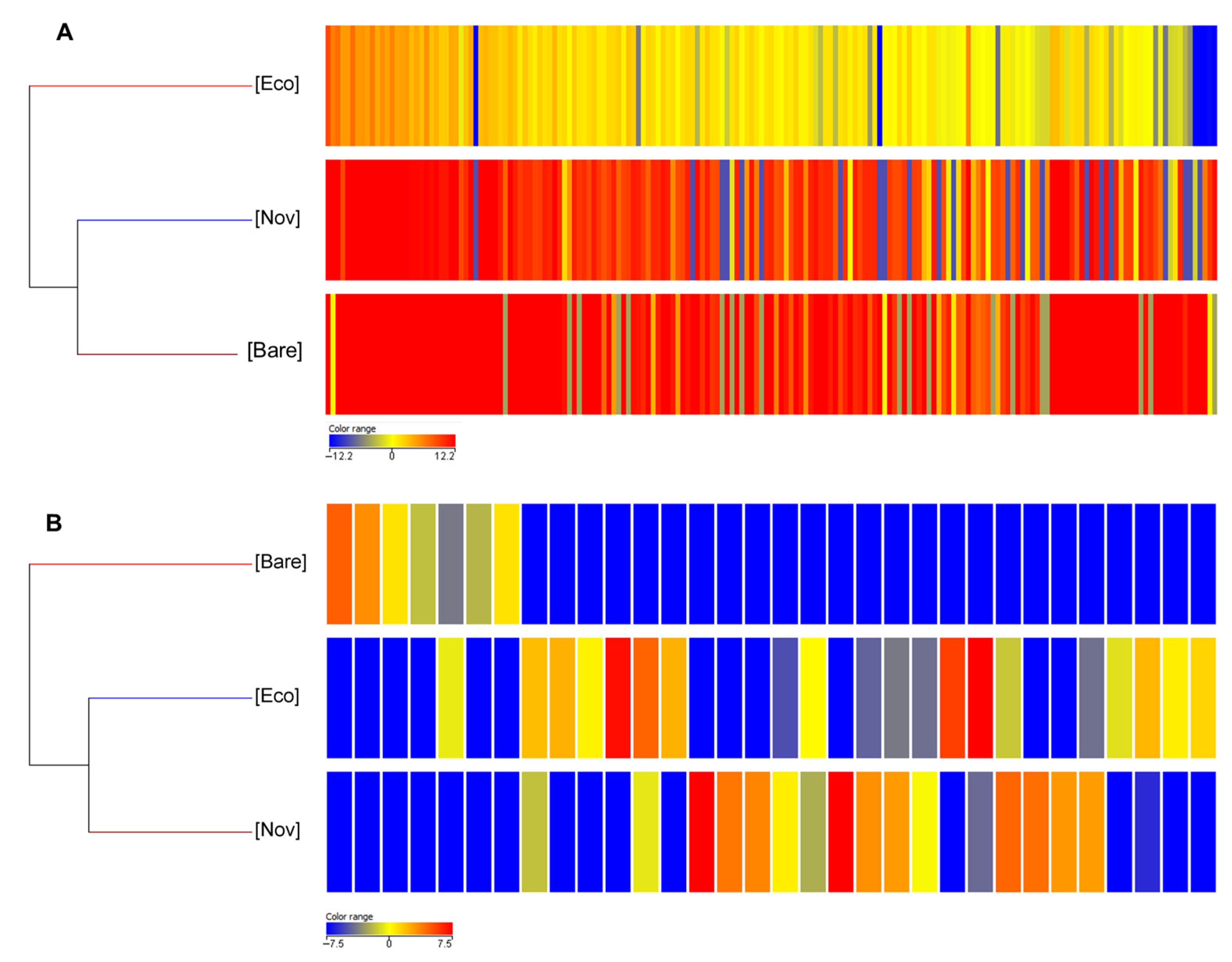
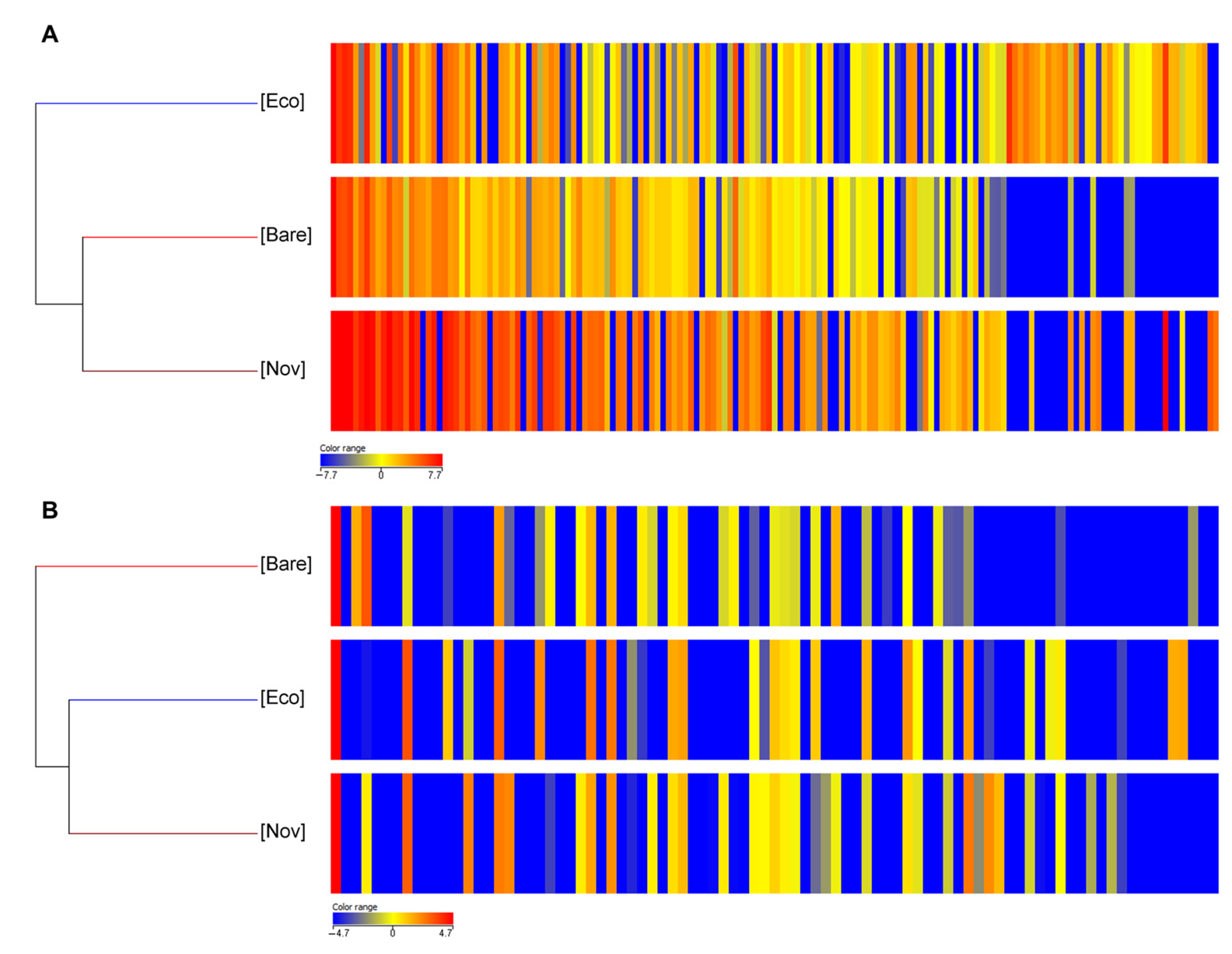
3.2. Untargeted Metabolomic Analysis
3.2.1. Leaves
3.2.2. Berries
3.3. Targeted Metabolomic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Processing Tomato Council (WPTC). WPTC Crop Update and World Production Estimate as of 23 October 2019. Available online: https://www.wptc.to/release-wptc.php (accessed on 24 February 2020).
- Frusciante, L.; Carli, P.; Ercolano, M.R.; Pernice, R.; Di Matteo, A.; Fogliano, V.; Pellegrini, N. Antioxidant nutritional quality of tomato. Mol. Nutr. Food Res. 2007, 51, 609–617. [Google Scholar] [CrossRef]
- Cristofano, F.; El-Nakhel, C.; Rouphael, Y. Biostimulant substances for sustainable agriculture: Origin, operating mechanisms and effects on cucurbits, leafy greens, and nightshade vegetables species. Biomolecules 2021, 11, 1103. [Google Scholar] [CrossRef] [PubMed]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- du Jardin, P. Plants Biostimulants: Definition, Concept, Main Categories and Regulations. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory Action: Designing the Next Generation of Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 13, 1655. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Pepe, O. Microbial Consortia: Promising Probiotics as Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1801. [Google Scholar] [CrossRef] [PubMed]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Weindling, R. Trichoderma lignorum as a Parasite of Other Soil Fungi. Phytopathology 1932, 22, 837–845. [Google Scholar]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: A multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef]
- Cai, F.; Druzhinina, I.S. In honor of John Bissett: Authoritative guidelines on molecular identification of Trichoderma. Fungal Divers. 2021, 107, 1–69. [Google Scholar] [CrossRef]
- Innocenti, G.; Roberti, R.; Piattoni, F. Biocontrol ability of Trichoderma harzianum strain T22 against Fusarium wilt disease on water-stressed lettuce plants. BioControl 2015, 60, 573–581. [Google Scholar] [CrossRef]
- Pocurull, M.; Fullana, A.M.; Ferro, M.; Valero, P.; Escudero, N.; Saus, E.; Gabaldón, T.; Sorribas, F.J. Commercial formulates of Trichoderma induce systemic plant resistance to Meloidogyne incognita in tomato and the effect is additive to that of the mi-1.2 resistance gene. Front. Microbiol. 2020, 10, 3042. [Google Scholar] [CrossRef]
- Vitti, A.; Pellegrini, E.; Nali, C.; Lovelli, S.; Sofo, A.; Valerio, M.; Scopa, A.; Nuzzaci, M. Trichoderma harzianum T-22 induces systemic resistance in tomato infected by Cucumber mosaic virus. Front. Plant. Sci. 2016, 7, 1520. [Google Scholar] [CrossRef]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Marra, R.; Vitale, S.; Pironti, A.; Fiorentino, N.; Mori, M. Yield and quality of processing tomato as improved by biostimulants based on Trichoderma sp. and Ascophyllum nodosum and biodegradable mulching films. Agronomy 2023, 13, 901. [Google Scholar] [CrossRef]
- Staropoli, A.; Vassetti, A.; Salvatore, M.M.; Andolfi, A.; Prigigallo, M.I.; Bubici, G.; Scagliola, M.; Salerno, P.; Vinale, F. Improvement of nutraceutical value of parsley leaves (Petroselinum crispum) upon field applications of beneficial microorganisms. Horticulturae 2021, 7, 281. [Google Scholar] [CrossRef]
- Kasirajan, S.; Ngouajio, M. Polyethylene and biodegradable mulches for agricultural applications: A review. Agron. Sustain. Dev. 2012, 32, 501–529. [Google Scholar] [CrossRef]
- Kyrikou, I.; Briassoulis, D. Biodegradation of Agricultural Plastic Films: A Critical Review. J. Polym. Environ. 2007, 15, 125–150. [Google Scholar] [CrossRef]
- Muroi, F.; Tachibana, Y.; Kobayashi, Y.; Sakurai, T.; Kasuya, K.I. Influences of poly (butylene adipate-co-terephthalate) on soil microbiota and plant growth. Polym. Degrad. Stab. 2016, 129, 338–346. [Google Scholar] [CrossRef]
- Taromi Aliabadi, B.; Hassandokht, M.R.; Etesami, H.; Alikhani, H.A.; Dehghanisanij, H. Effect of mulching on some characteristics of tomato (Lycopersicon esculentum Mill.) under deficit irrigation. J. Agric. Sci. Technol. 2019, 21, 927–941. [Google Scholar]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Sacco, A.; El-Nakhel, C.; Rouphael, Y.; Mori, M. Trichoderma spp. and mulching films differentially boost qualitative and quantitative aspects of greenhouse lettuce under diverse N conditions. Horticulturae 2020, 6, 55. [Google Scholar] [CrossRef]
- Marra, R.; Lombardi, N.; Piccolo, A.; Bazghaleh, N.; Prashar, P.; Vandenberg, A.; Woo, S. Mineral biofortification and growth stimulation of lentil plants inoculated with Trichoderma strains and metabolites. Microorganisms 2022, 10, 87. [Google Scholar] [CrossRef]
- Marra, R.; Lombardi, N.; d’Errico, G.; Troisi, J.; Scala, G.; Vinale, F.; Woo, S.L.; Bonanomi, G.; Lorito, M. Application of Trichoderma strains and metabolites enhances soybean productivity and nutrient content. J. Agric. Food Chem. 2019, 67, 1814–1822. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Sivasithamparam, K. Beneficial effects of Trichoderma secondary metabolites on crops. Phytother. Res. 2020, 34, 2835–2842. [Google Scholar] [CrossRef]
- Du Jardin, P.; Xu, L.; Geelen, D. Agricultural functions and action mechanisms of plant biostimulants (PBs). In The Chemical Biology of Plant Biostimulants; Geelen, D., Xu, L., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 1–30. [Google Scholar]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Fleming, T.R.; Fleming, C.C.; Levy, C.C.B.; Repiso, C.; Hennequart, F.; Nolasco, J.B.; Liu, F. Biostimulants enhance growth and drought tolerance in Arabidopsis thaliana and exhibit chemical priming action. Ann. Appl. Biol. 2019, 174, 153–165. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Labuschagne, N.; Dubery, I.A. Metabolic profiling of PGPR-treated tomato plants reveal priming-related adaptations of secondary metabolites and aromatic amino acids. Metabolites 2020, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Diretto, G.; Digilio, M.C.; Woo, S.L.; Giuliano, G.; Molisso, D.; Pennacchio, F.; Lorito, M.; Rao, R. Transcriptome and Metabolome Reprogramming in Tomato Plants by Trichoderma harzianum strain T22 Primes and Enhances Defense Responses Against Aphids. Front. Physiol. 2019, 10, 745. [Google Scholar] [CrossRef] [PubMed]
- Salvi, L.; Brunetti, C.; Cataldo, E.; Niccolai, A.; Centritto, M.; Ferrini, F.; Mattii, G.B. Effects of Ascophyllum nodosum extract on Vitis vinifera: Consequences on plant physiology, grape quality and secondary metabolism. Plant Physiol. Biochem. 2019, 139, 21–32. [Google Scholar] [CrossRef]
- Wu, C.; Ma, Y.; Wang, D.; Shan, Y.; Song, X.; Hu, H.; Ren, X.; Ma, X.; Cui, J.; Ma, Y. Integrated microbiology and metabolomics analysis reveal plastic mulch film residue affects soil microorganisms and their metabolic functions. J. Hazard. Mater. 2022, 423, 127258. [Google Scholar] [CrossRef]
- Bandopadhyay, S.; Martin-Closas, L.; Pelacho, A.M.; DeBruyn, J.M. Biodegradable plastic mulch films: Impacts on soil microbial communities and ecosystem functions. Front. Microbiol. 2018, 9, 819. [Google Scholar] [CrossRef]
- Bandopadhyay, S.; Sintim, H.Y.; DeBruyn, J.M. Effects of biodegradable plastic film mulching on soil microbial communities in two agroecosystems. Peer J. 2020, 8, e9015. [Google Scholar] [CrossRef]
- Kozukue, N.; Han, J.S.; Lee, K.R.; Friedman, M. Dehydrotomatine and alpha-tomatine content in tomato fruits and vegetative plant tissues. J. Agric. Food Chem. 2004, 52, 2079–2083. [Google Scholar] [CrossRef]
- Fontaine, T.D.; Irving, G.W., Jr. Isolation and partial characterization of crystalline tomatine, an antibiotic agent from the tomato plant. Arch. Biochem. 1948, 18, 467–475. [Google Scholar]
- Leonardi, C.; Ambrosino, P.; Esposito, F.; Fogliano, V. Antioxidative activity and carotenoid and tomatine contents in different typologies of fresh consumption tomatoes. J. Agric. Food Chem. 2000, 48, 4723–4727. [Google Scholar] [CrossRef]
- Kozukue, N.; Friedman, M. Tomatine, chlorophyll, β-carotene and lycopene content in tomatoes during growth and maturation. J. Sci. Food Agric. 2003, 83, 195–200. [Google Scholar] [CrossRef]
- Lanzuise, S.; Manganiello, G.; Guastaferro, V.M.; Vincenzo, C.; Vitaglione, P.; Ferracane, R.; Vecchi, A.; Vinale, F.; Kamau, S.; Lorito, M.; et al. Combined biostimulant applications of Trichoderma spp. with fatty acid mixtures improve biocontrol activity, horticultural crop yield and nutritional quality. Agronomy 2022, 12, 275. [Google Scholar] [CrossRef]
- Bailly, C. The steroidal alkaloids α-tomatine and tomatidine: Panorama of their mode of action and pharmacological properties. Steroids 2021, 176, 108933. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Haslam, E. Practical Polyphenolics. From Structure to Molecular Recognition and Physiological Action; Cambridge University Press: New York, NY, USA, 1998; 422p. [Google Scholar]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Lev-Yadun, S. Plants are not sitting ducks waiting for herbivores to eat them. Plant Signal. Behav. 2016, 11, e1179419. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, M.; Hara-Nishimura, I. Specialized Vacuoles of Myrosin Cells: Chemical Defense Strategy in Brassicales Plants. Plant Cell Physiol. 2018, 59, 1309–1316. [Google Scholar] [CrossRef]
- Yactayo-Chang, J.P.; Tang, H.V.; Mendoza, J.; Christensen, S.A.; Block, A.K. Plant Defense Chemicals against Insect Pests. Agronomy 2020, 10, 1156. [Google Scholar] [CrossRef]
- Chavan, S.B.; Koche, K. Correlating medicarpin content of chickpea cultivars as a key defense compound against Fusarium wilt. Res. Crops 2021, 22, 178–182. [Google Scholar] [CrossRef]
- Kale, M.C.; Choudhary, A.D. Induction of phenylalanine ammonia lyase, in groundnut cultivars in response to biotic and abiotic stress. Indian Phytopathol. 2001, 54, 288–292. [Google Scholar]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Enhancement of Phenolic and Flavonoid Compounds in Cabbage (Brassica oleraceae) Following Application of Commercial Seaweed Extracts of the Brown Seaweed, (Ascophyllum nodosum). Agric. Food Sci. 2013, 22, 288–295. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.M. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Marra, R.; Coppola, M.; Pironti, A.; Grasso, F.; Lombardi, N.; d’Errico, G.; Sicari, A.; Bolletti Censi, S.; Woo, S.L.; Rao, R.; et al. The application of Trichoderma strains or metabolites alters the olive leaf metabolome and the expression of defense-related genes. J. Fungi 2020, 6, 369. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Pascale, M.; Staropoli, A.; Marra, R.; Vinale, F. Effect of Selected Trichoderma Strains and Metabolites on Olive Drupes. Appl. Sci. 2021, 11, 8710. [Google Scholar] [CrossRef]
- Mazzei, P.; Vinale, F.; Woo, S.L.; Pascale, A.; Lorito, M.; Piccolo, A. Metabolomics by Proton High-Resolution Magic-Angle-Spinning Nuclear Magnetic Resonance of Tomato Plants Treated with Two Secondary Metabolites Isolated from Trichoderma J. Agric. Food Chem. 2016, 64, 3538–3545. [Google Scholar] [CrossRef] [PubMed]
- Pakora, G.A.; Mpika, J.; Kone, D.; Ducamp, M.; Kebe, I.; Nay, B.; Buisson, D. Inhibition of Phytophthora species, agents of cocoa black pod disease, by secondary metabolites of Trichoderma species Environ. Sci. Pollut. Res. Int. 2018, 25, 29901–29909. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| N. | Origin | Ionization | RT | Compound | Class | Formula | Experimental Mass (Da) | Theoretical Mass (Da) |
|---|---|---|---|---|---|---|---|---|
| 1 | L | − | 1.257 | p-Coumaroylquinic acid | Phenol | C16H18O8 | 338.0854 | 338.10016753 |
| 2 | L | + | 1.786 | Phenethylamine | Alkaloid | C8H11N | 121.0895 | 121.089149355 |
| 3 | L | − | 1.831 | Lupinisoflavone E | Flavonoid | C25H26O7 | 438.1655 | 438.16785316 |
| 4 | L | − | 4.578 | 4-Caffeoylquinic acid | Phenol | C16H18O9 | 354.0938 | 354.09508215 |
| 5 | L | − | 4.619 | Kaempferol 3-glucosyl-(1->2)-galactoside-7-glucoside | Flavonoid glycoside | C33H40O21 | 772.2061 | 772.20620828 |
| 6 | L | − | 4.703 | 2′-Hydroxyisolupalbigenin | Flavonoid | C25H26O6 | 422.1707 | 422.17293854 |
| 7 | L | − | 4.705 | Icariside B2 | Glycoside | C19H30O8 | 386.1939 | 386.19406791 |
| 8 | L | − | 4.714 | Manghaslin | Flavonoid glycoside | C33H40O20 | 756.2113 | 756.21129366 |
| 9 | L | − | 4.857 | Quercetin 3-glucosyl-(1->6)-galactoside | Flavonoid glycoside | C27H30O17 | 626.148 | 626.14829948 |
| 10 | L | +/− | 4.895 | Cucurbitacin K 2-O-beta-D-glucopyranoside | Steroidal glycoside | C36H54O13 | 694.3571 | 694.35644177 |
| 11 | L | + | 5.012 | Hyperin | Flavonoid glycoside | C21H19O12 | 464.0943 | 464.09547607 |
| 12 | L | + | 5.020 | Hydroxytomatine | Steroidal glycoalkaloid | C50H85NO22 | 1052.5536 | 1052.56260009 |
| 13 | L | − | 5.051 | Phaseolic acid | Phenol | C13H12O8 | 296.0532 | 296.05321734 |
| 14 | L | + | 5.145 | Fisetin | Flavonoid | C15H10O6 | 286.0457 | 286.04773803 |
| 15 | L | +/− | 5.146 | Kaempferol 3-galactoside-7-rhamnoside | Flavonoid glycoside | C27H30O15 | 594.1581 | 594.15847025 |
| 16 | L | − | 5.146 | Myricitrin | Flavonoid glycoside | C21H20O12 | 464.0954 | 464.09547607 |
| 17 | L | − | 5.192 | Uttroside B | Saponin | C56H94O28 | 1214.5933 | 1214.59316234 |
| 18 | L | + | 5.228 | Lycoperoside C | Steroidal saponin | C52H85NO23 | 1091.551 | 1091.55123796 |
| 19 | L | + | 5.355 | β-2-Tomatine | Steroidal glycoalkaloid | C44H73NO16 | 871.4934 | 871.49293524 |
| 20 | L | +/− | 5.383 | γ-Tomatine | Steroidal glycoalkaloid | C39H65NO12 | 739.4534 | 739.45067651 |
| 21 | L | − | 5.388 | N-Malonyltryptophan | Aminoacid | C14H14N2O5 | 290.09 | 290.09027155 |
| 22 | L | − | 5.487 | Acuminoside | Terpene glycoside | C21H36O10 | 448.2299 | 448.23084734 |
| 23 | L | − | 5.581 | N-cis-Feruloyloctopamine | Phenol | C18H19NO5 | 329.1261 | 329.12632271 |
| 24 | L | − | 5.625 | 2-Methoxymedicarpin | Isoflavonoid | C17H16O5 | 300.1002 | 300.09977361 |
| 25 | L | + | 5.638 | Colnelenic acid | Fatty acid | C18H28O3 | 292.2037 | 292.20384475 |
| 26 | L | + | 5.647 | Apo-13-zeaxanthinone | Terpene | C18H26O2 | 274.1922 | 274.193280068 |
| 27 | L | − | 5.686 | (+)-Medicarpin | Isoflavonoid | C16H14O4 | 270.0891 | 270.08920892 |
| 28 | L | +/− | 5.864 | N-trans-Feruloyltyramine | Phenol | C18H19NO4 | 313.1315 | 313.13140809 |
| 29 | B | + | 1.342 | Lotaustralin | Cyanogenic glycoside | C11H19NO6 | 261.1209 | 261.12123733 |
| 30 | B | + | 1.499 | Linamarin | Cyanogenic glycoside | C10H17NO6 | 247.1054 | 247.10558726 |
| 31 | B | + | 1.551 | Dihydrozeatin | 6-alkylaminopurines | C10H15N5O | 243.1199 | 221.12766012 |
| 32 | B | + | 1.788 | (+)-Ligballinol | Furanoid lignan | C18H18O4 | 298.1235 | 298.12050905 |
| 33 | B | + | 1.799 | Luteolin 3′,5-dimethyl ether | Flavonoid | C23H24O11 | 314.0776 | 314.07903816 |
| 34 | B | − | 4.136 | 5-Caffeoylquinic acid | Phenol | C16H18O9 | 354.095 | 354.09508215 |
| 35 | B | − | 4.206 | L-Phenylalanine | Aminoacid | C9H11NO2 | 165.0789 | 165.078978594 |
| 36 | B | + | 4.743 | Tryptamine | Alkylindole | C10H12N2 | 160.0993 | 160.100048391 |
| 37 | B | − | 4.939 | Lycoperoside F | Steroidal glycoalkaloid | C58H95NO29 | 1269.5977 | 1269.59897599 |
| 38 | B | − | 5.026 | Phloretin 3′,5′-Di-C-glucoside | Flavonoid glycoside | C27H34O15 | 598.1898 | 598.18977037 |
| 39 | B | − | 5.108 | Taxifolin 3,7-dirhamnoside | Flavonoid glycoside | C27H32O15 | 596.1727 | 596.17412031 |
| 40 | B | − | 5.131 | Naringenin-7-O-glucoside | Flavonoid glycoside | C21H22O10 | 434.1213 | 434.12129689 |
| 41 | B | + | 5.182 | Anhydropisatin | Isoflavonoid | C17H12O5 | 296.0672 | 296.06847348 |
| 42 | B | + | 5.198 | Indioside D | Saponin | C51H84O23 | 1064.5417 | 1064.54033892 |
| 43 | B | +/− | 5.226 | Biochanin A | Flavonoid | C16H12O5 | 284.0681 | 284.06847348 |
| 44 | B | − | 5.254 | 12-Hydroxyjasmonic acid glucoside | Fatty acyl glycoside | C18H28O9 | 388.1723 | 388.17333247 |
| 45 | B | + | 5.278 | Phloroglucinol | Benzenetriol | C6H6O3 | 126.0318 | 126.031694049 |
| 46 | B | − | 5.410 | Formononetin 7-O-beta-D-glucoside-6″-O-malonate | Flavonoid | C25H24O12 | 516.1261 | 516.12677620 |
| 47 | B | − | 5.416 | N-Acetyltryptophan | Aminoacid | C13H14N2O3 | 246.099 | 246.10044231 |
| 48 | B | + | 5.737 | Isoesculeogenin A | Saponin | C27H45NO4 | 447.3326 | 447.33485892 |
| 49 | B | − | 6.030 | (2R,3R)-fustin | Flavonoid | C15H12O6 | 288.0632 | 288.06338810 |
| 50 | B | + | 6.221 | Butin | Flavonoid | C15H12O5 | 272.0677 | 272.06847348 |
| 51 | B | − | 7.207 | Nordihydrocapsiate | Phenol | C17H26O4 | 294.1835 | 294.18310931 |
| 52 | B | +/− | 9.733 | Colneleic acid | Fatty acid | C18H30O3 | 294.2197 | 294.21949481 |
| 53 | B/L | − | 4.796 | Quinic acid | Cyclic carboxylic acid | C7H12O6 | 192.0631 | 192.06338810 |
| 54 | B/L | +/− | 4.851 | Quercetin 3-(2G-apiosylrutinoside) | Flavonoid glycoside | C32H38O20 | 742.1959 | 742.19564360 |
| 55 | B/L | + | 5.009 | Panasenoside | Flavonoid glycoside | C27H30O16 | 610.1529 | 610.15338487 |
| 56 | B/L | + | 5.010 | Robinetin | Flavonoid | C15H10O7 | 302.0414 | 302.04265265 |
| 57 | B/L | − | 5.029 | Isoorientin 2″-O-glucopyranoside | Flavonoid glycoside | C27H30O16 | 610.1535 | 610.15338487 |
| 58 | B/L | − | 5.093 | Caffeic acid | Phenol | C15H18O9 | 180.0419 | 180.04225873 |
| 59 | B/L | +/− | 5.229 | Dehydrotomatine | Steroidal glycoalkaloid | C50H81NO21 | 1031.5343 | 1031.53010859 |
| 60 | B/L | +/− | 5.267 | α-Tomatine | Steroidal glycoalkaloid | C50H83NO21 | 1033.545 | 1033.54575866 |
| 61 | B/L | + | 5.273 | Robeneoside B | Steroidal glycoalkaloid | C45H73NO17 | 899.488 | 899.48784986 |
| 62 | B/L | + | 5.303 | δ-Tomatine | Steroidal glycoalkaloid | C33H55NO7 | 577.3983 | 577.39785309 |
| 63 | B/L | +/− | 5.318 | β1-Tomatine | Steroidal glycoalkaloid | C45H75NO17 | 901.4983 | 901.50349992 |
| 64 | B/L | + | 6.975 | Tomatidine | Steroidal alkaloid | C27H45NO | 415.3428 | 415.345029678 |
| 65 | B/L | + | 7.213 | Solasodine | Steroidal alkaloid | C27H43NO2 | 413.329 | 413.329379614 |
| Source: Tomato Leaves | Bare Micro vs. Bare | Bare Bio vs. Bare | Bare Micro+Bio vs. Bare | Eco Micro vs. Bare Micro | Nov Micro vs. Bare Micro | Eco Micro+Bio vs. Bare Micro+Bio | Nov Micro+Bio vs. Bare Micro+Bio |
|---|---|---|---|---|---|---|---|
| UP | 129 | 159 | 138 | 46 | 53 | 142 | 176 |
| DOWN | 159 | 129 | 150 | 169 | 162 | 104 | 70 |
| Source: Tomato Berries | Bare Micro vs. Bare | Bare Bio vs. Bare | Bare Micro+Bio vs. Bare | Eco Micro vs. Bare Micro | Nov Micro vs. Bare Micro | Eco Micro+Bio vs. Bare Micro+Bio | Nov Micro+Bio vs. Bare Micro+Bio |
|---|---|---|---|---|---|---|---|
| UP | 106 | 92 | 73 | 225 | 252 | 25 | 38 |
| DOWN | 208 | 222 | 241 | 129 | 102 | 52 | 39 |
| Compound | vs. Bare Ctrl | vs. Bare Micro | |||
|---|---|---|---|---|---|
| Bio | Micro | Bio+Micro | Nov Micro | Eco Micro | |
| p-Coumaroylquinic acid | ↓ | ↓↓↓ | ↓ | ND | ND |
| Phenethylamine | ↓↓↓ | ↓↓↓ | - | ND | ND |
| Lupinisoflavone E | ↑ | ↑↑↑ | - | ↓↓↓ | ↓↓↓ |
| 4-Caffeoylquinic acid | - | ↓↓↓ | ↑ | ↑↑↑ | ↑↑ |
| Manghaslin | ↑ | - | ↑↑↑ | ND | ND |
| Cucurbitacin K 2-O-beta-D-glucopyranoside | ↓ | ↓↓↓ | ↓ | ↑↑↑ | ↑↑↑ |
| Hyperin | ND | ND | ND | ↓↓↓ | ↓ |
| Hydroxytomatine | - | - | ↑↑↑ | ↓↓↓ | ↓ |
| Phaseolic acid | ↑ | - | ↑↑ | ND | ND |
| Fisetin | - | ↑ | - | ↑↑ | ↓ |
| Myricitrin | ND | ND | ND | ↑↑ | ↑↑↑ |
| Uttroside B | ↑↑ | ↓ | ↑↑ | ND | ND |
| Lycoperoside C | ND | ND | ND | ↓↓ | ↓ |
| γ-Tomatine | - | - | ↓ | ↑↑ | ↑↑↑ |
| N-malonyltryptophan | ↑ | ↓ | - | ND | ND |
| 2-Methoxymedicarpin | ↑ | ↑↑ | ↑↑↑ | ND | ND |
| Colnelenic acid | ND | ND | ND | ↓↓↓ | ↓ |
| Apo-13-zeaxanthinone | ND | ND | ND | ↓↓↓ | ↓ |
| (+)-Medicarpin | ↑↑↑ | ↑↑↑ | ↑↑↑ | ND | ND |
| N-trans-feruloyltyramine | ND | ND | ND | ↓↓↓ | ↓ |
| Lotaustralin | ND | ND | ND | ↓ | ↓ |
| Linamarin | ↓↓↓ | ↓↓ | ↓ | ND | ND |
| Dihydrozeatin | ↓ | - | ↓↓ | ↓↓ | ↓↓ |
| (+)-Ligballinol | - | ↑ | ↓↓ | ND | ND |
| Luteolin 3′,5′-dimethyl ether | ↑ | ↑↑↑ | - | ↓↓↓ | ↓↓↓ |
| 5-Caffeoylquinic acid | ↓ | ↓ | ↓ | - | - |
| L-Phenylalanine | ↓ | ↓ | ↓ | - | - |
| Lycoperoside F | ↓ | ↓ | ↓ | ND | ND |
| Phloretin 3′, 5′- Di-C-glucoside | ND | ND | ND | - | - |
| Taxifolin 3,7-dirhamnoside | ↓ | ↓ | ↓ | - | - |
| Naringenin-7-O-glucoside | ↓↓ | ↓↓ | ↓ | - | ↑ |
| Anhydropisatin | ↓ | ↑ | ↑ | ↓↓ | ↓ |
| Biochanin A | ↓↓ | ↓↓ | ↑ | - | ↑↑ |
| 12-Hydroxyjasmonic acid glucoside | ↓ | ↓ | ↓ | - | - |
| Formononetin 7-O-beta-D-glucoside-6″-O-malonate | ↓↓ | ↓ | ↓ | - | - |
| N-Acetyltryptophan | ND | ND | ND | ↑ | ↑ |
| Isoesculeogenin A | ↑ | - | - | ND | ND |
| (2R,3R)-fustin | ↑↑ | ↑↑ | ↑↑ | - | - |
| Butin | ND | ND | ND | ↑ | ↑↑ |
| Nordihydrocapsiate | ND | ND | ND | ↓↓↓ | ↓↓ |
| Colneleic acid | ND | ND | ND | - | - |
| Quinic acid | ↓↑ | ↓↓↓↓ | ↓↑ | ↓↓↓↓↓ | ↓↓↓↓ |
| Quercetin 3-(2G-apiosylrutinoside) | ND ↑ | ND - | ND ↑ | - ND | - ND |
| Panasenoside | ND - | ND ↑ | ND - | -↓↓↓ | -↓ |
| Robinetin | -↑ | -↑ | -↑ | ND↓ | ND↓ |
| Isoorientin 2″-O-glucopyranoside | - - | - - | - - | - ND | ↑ ND |
| Caffeic acid | ↓ - | ↓ - | ↓ - | ND ↑↑↑ | ND ↑↑↑ |
| Dehydrotomatine | ↑↑- | ↑↑↑↓ | ↑↑↑- | ND↓↓↓ | ND↓ |
| α-Tomatine | -- | -↓ | -↑ | ND↓↓ | ND↓ |
| Robeneoside B | ND ND | ND ND | ND ND | ↑↓↓↓ | -↓ |
| δ-Tomatine | -- | - - | - - | ND↓↓↓ | ND↓ |
| β 1-Tomatine | ND↓ | ND↓↓↓ | ND↓ | -↓↓↓ | ↑↑↓ |
| Tomatidine | ↑ - | ↑↑ - | ↑↑ - | ↑↑↓↓↓ | ↑↓ |
| Solasodine | ↑- | - - | -- | ↑↑↓↓↓ | ↑↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staropoli, A.; Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Pironti, A.; Lombardi, N.; Nanni, B.; Mori, M.; Vinale, F.; Woo, S.L.; et al. Biodegradable Mulch Films and Bioformulations Based on Trichoderma sp. and Seaweed Extract Differentially Affect the Metabolome of Industrial Tomato Plants. J. Fungi 2024, 10, 97. https://doi.org/10.3390/jof10020097
Staropoli A, Di Mola I, Ottaiano L, Cozzolino E, Pironti A, Lombardi N, Nanni B, Mori M, Vinale F, Woo SL, et al. Biodegradable Mulch Films and Bioformulations Based on Trichoderma sp. and Seaweed Extract Differentially Affect the Metabolome of Industrial Tomato Plants. Journal of Fungi. 2024; 10(2):97. https://doi.org/10.3390/jof10020097
Chicago/Turabian StyleStaropoli, Alessia, Ida Di Mola, Lucia Ottaiano, Eugenio Cozzolino, Angela Pironti, Nadia Lombardi, Bruno Nanni, Mauro Mori, Francesco Vinale, Sheridan Lois Woo, and et al. 2024. "Biodegradable Mulch Films and Bioformulations Based on Trichoderma sp. and Seaweed Extract Differentially Affect the Metabolome of Industrial Tomato Plants" Journal of Fungi 10, no. 2: 97. https://doi.org/10.3390/jof10020097