
Floristic Similarities between the Lichen Flora of Both Sides of the Drake Passage: A Biogeographical Approach


Abstract
:1. Introduction
2. Materials and Methods
2.1. Common Species between Navarino Island and the West Antarctic Region
2.2. Similarity Indexes
2.3. Distributional Pattern Analysis
3. Results
3.1. Common Species between Navarino Island and the West Antarctic Region and Similarity Indexes
3.2. Distributional Pattern Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Convey, P.; Bowman, V.; Chown, S.L.; Francis, J.; Fraser, C.; Smellie, J.L.; Storey, B.; Terauds, A. Ice-Bound Antarctica: Biotic Consequences of the Shift from a Temperate to a Polar Climate. In Mountains, Climate and Biodiversity, 1st ed.; Hoorn, C., Perrigo, A., Antonelli, A., Eds.; John Wiley & Sons: Oxford, UK, 2018; pp. 355–373. ISBN 978-1-119-15987-2. [Google Scholar]
- Muñoz, J.; Felicisimo, A.M.; Cabezas, F.; Burgaz, A.R.; Martinez, I. Wind as a long-distance dispersal vehicle in the Southern Hemisphere. Science 2004, 304, 1144–1147. Available online: https://www.science.org/doi/10.1126/science.1095210 (accessed on 15 September 2023). [CrossRef] [PubMed]
- Terauds, A.; Chown, S.L.; Morgan, F.; Peat, H.J.; Watts, D.J.; Keys, H.; Convey, P.; Bergstrom, D.M. Conservation biogeography of the Antarctic. Divers. Distrib. 2012, 18, 726–741. [Google Scholar] [CrossRef]
- Chown, S.L.; Clarke, A.; Fraser, C.I.; Cary, S.C.; Moon, K.L.; McGeoch, M.A. The changing form of Antarctic biodiversity. Nature 2015, 522, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Rozzi, R.; Massardo, F.; Anderson, C.; Berghoefer, A.; Mansilla, A.; Mansilla, M.; Plana, J.; Berghöfer, U.; Araya, P.; Barros, E.; et al. La Reserva de Biosfera Cabo de Hornos, 1st ed.; Ediciones Universidad de Magallanes: Punta Arenas, Chile, 2006. [Google Scholar]
- Laguna, C.; Pintado, A.; Green, A.; Blanquer, J.M.; Sancho, L.G. Distributional and ecophysiological study on the Antarctic lichens species pair Usnea antarctica/Usnea aurantiaco-atra. Pol. Biol. 2016, 39, 1183–1195. [Google Scholar] [CrossRef]
- Etayo, J.; Sancho, L.G.; Gómez-Bolea, A.; Søchting, U.; Aguirres, F.; Rozzi, R. Catalogue of lichens (and some related fungi) of Navarino Island, Cape Horn Biosphere Reserve, Chile. Ann. Inst. Patagon. 2021, 49, 1–110. [Google Scholar] [CrossRef]
- Sancho, L.G.; Kappen, L.; Schroeter, B. The lichen genus Umbilicaria on Livingston Island (South Shetland Islands, Antarctica). Antarct. Sci. 1992, 4, 189–196. [Google Scholar] [CrossRef]
- Sancho, L.G.; Schulz, F.; Schroeter, B.; Kappen, L. Bryophyte and lichen flora of South Bay (Livingston Island, South Shetland Islands, Antarctica). Nova Hedwig. 1999, 68, 301–337. [Google Scholar] [CrossRef]
- Søchting, U.; Øvstedal, D.O.; Sancho, L.G. The lichens of Hurd Peninsula, Livingston Island, South Shetlands, Antarctica. In Contributions to Lichenology. Festschrift in Honour of Hannes Hertel, 1st ed.; Döbbeler, P., Rambold, G., Eds.; Bibliotheca Lichenologica; J. Cramer in der Gebrüder Borntraeger: Berlin/Germany, Germany, 2004; Volume 88, pp. 607–658. ISBN 978-3-443-58067-4. [Google Scholar]
- Burgaz, A.R.; Raggio, J. The Cladoniaceae of Navarino Island (Prov. Antártica Chilena, Chile). Mycotaxon 2007, 99, 103–116. [Google Scholar]
- Øvstedal, D.O.; Lewis-Smith, R.I. Lichens of Antarctica and South Georgia. A Guide to their Identification and Ecology, 1st ed.; Cambridge University Press: Cambridge, UK, 2001; ISBN 978-0-521-66241-3. [Google Scholar]
- Olech, M. Lichens of King George Island, Antarctica, 1st ed.; Institute of Botany of Jagiellonian University: Kraków, Poland, 2004; ISBN 978-83-915161-7-1. [Google Scholar]
- Meyer, D.; Zeileis, A.; Hornik, K. vcd: Visualizing Categorical Data; R Package Version 1.4.8. 2020. Available online: https://CRAN.R-project.org/package=vcd (accessed on 16 October 2023).
- Mangiafico, S. Rcompanion: Functions to Support Extension Education Program Evaluation. R Package Version 4.1.0. 2022. Available online: https://CRAN.R-project.org/package=rcompanion (accessed on 16 October 2023).
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arachchige, C.; Arppe, A.; Baddeley, A.; Barton, K.; Bolker, B.; et al. DescTools: Tools for descriptive statistics. R package Version 0.99.30. 2019. Available online: https://CRAN.R-project.org/package=DescTools (accessed on 16 October 2023).
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Dunnington, D.; Posit, P.B.C. Ggplot2: Elegant Graphics for Data Analysis. R package Version 3.4.4. 2016. Available online: https://CRAN.R-project.org/package=ggplot2 (accessed on 16 October 2023).
- Türk, R.; Hogg, I.; Cox, E.R.; Sancho, L.G.; Williamson, S.N.; Bryan, V.; Green, T.G.A. The lichens of Cambridge Bay and vicinity. Nunavut, Victoria Island, Canada. University of Salzburg: Salzbug, Austria, 2024; to be submitted. [Google Scholar]
- Garrido-Benavent, I.; De los Ríos, A.; Pérez-Ortega, S. From Alaska to Antarctica: Species boundaries and genetic diversity of Prasiola (Trebouxiophyceae), a foliose chlorophyte associated with the bipolar lichen-forming fungus Mastodia tessellata. Mol. Phylogenet. Evol. 2016, 107, 117–131. [Google Scholar] [CrossRef]
- Garrido-Benavent, I.; Pérez-Ortega, S.; De los Ríos, A.; Fernández-Mendoza, F. Amphitropical variation of the algal partners of Pseudephebe (Parmeliaceae, lichenized fungi). Symbiosis 2020, 82, 35–48. [Google Scholar] [CrossRef]
- Zúñiga, C.; Leiva, D.; Ramírez-Fernández, L.; Carú, M.; Yahr, R.; Orlando, J. Phylogenetic Diversity of Peltigera cyanolichens and Their Photobionts in Southern Chile and Antarctica. Microbes Environ. 2015, 30, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Ochyra, R.; Smith, R.I.L.; Bednarek-Ochyra, H. The Illustrated Moss Flora of Antarctica, 1st ed.; Cambridge University Press: Cambridge, UK, 2008; ISBN 978-0-521-81402-7. [Google Scholar]
- Segovia, R.; Armesto, J.J. The Gondwanan legacy in South American biogeography. J. Biogeogr. 2015, 42, 209–217. [Google Scholar] [CrossRef]
- Barreda, V.D.; Palazzesi, L.; Pujana, R.R.; Panti, C.; Tapia, M.J.; Fernández, D.A.; Noetinger, S. The Gondwanan heritage of the Eocene–Miocene Patagonian floras. J. S. Am. Earth Sci. 2021, 107, 103022. [Google Scholar] [CrossRef]
- Vanderpoorten, A.; Gradstein, S.R.; Carine, M.A.; Devos, N. The ghosts of Gondwana and Laurasia in modern liverwort distributions. Biol. Rev. 2010, 85, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Convey, P.; Biersma, E.M.; Casanova-Katny, A.; Maturana, C.S. Refuges of Antarctic diversity. In Past Antarctica: Paleoclimatology and Climate Change, 1st ed.; Oliva, M., Ruiz-Fernández, J., Eds.; Academic Press: New York, NY, USA, 2020; pp. 181–200. ISBN 978-0-12-817925-3. [Google Scholar]
- Stevens, M.I.; Mackintosh, A.N. Location, location, location: Survival of Antarctic biota requires the best real estate. Biol. Lett. 2023, 19, 20220590. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.A.; Ireland, L.; Convey, P.; Fleming, A.H. Assessing the effectiveness of specially protected areas for conservation of Antarctica’s botanical diversity. Conserv. Biol. 2016, 30, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Huiskes, A.H.L.; Gremmen, N.J.M.; Bergstrom, D.M.; Frenot, Y.; Hughes, K.A.; Imura, S.; Kiefer, K.; Lebouvier, M.; Lee, J.E.; Tsujimoto, M.; et al. Aliens in Antarctica: Assessing transfer of plant propagules by human visitors to reduce invasion risk. Biol. Conserv. 2014, 171, 278–284. [Google Scholar] [CrossRef]
- Coronato, A.; Borromei, A.M.; Ponce, J.F.; Candel, S.; Musotto, L.; Fernández, M.; Laprida, C.; Mehl, A.; Montes, A.; San-Martín, C.; et al. Holocene environmental changes in the fuegian forest and steppe, Argentina. J. S. Am. Earth Sci. 2022, 119, 103952. [Google Scholar] [CrossRef]
- Unkel, I.; Fernandez, M.; Björcka, S.; Ljung, K.; Wohlfarth, B. Records of environmental changes during the Holocene from Isla de los Estados (54.4°S), southeastern Tierra del Fuego. Glob. Planet. Change 2010, 74, 99–113. [Google Scholar] [CrossRef]
- Reynhout, S.; Sagredo, E.; Kaplan, M.; Aravena, J.C.; Martini, M.; Moreno, P.; Rojas, M.; Schwartz, R.; Schaefer, J. Holocene glacier fluctuations in Patagonia are modulated by summer insolation intensity and paced by Southern Annular Mode-like variability. Quat. Sci. Rev. 2019, 220, 178–187. [Google Scholar] [CrossRef]
- Galloway, D.J. Flora of New Zealand: Lichens, Including Lichen-Forming and Lichenicolous Fungi. Revised Second Edition, 2nd ed.; Manaaki Whenua Press: Lincoln, New Zealand, 2007; ISBN 987-0-478-09376-6. [Google Scholar]
- Sancho, L.; De Los Ríos, A.; Pintado, A.; Colesie, C.; Raggio, J.; Ascaso, C.; Green, A. Himantormia lugubris, an Antarctic endemic on the edge of the lichen symbiosis. Symbiosis 2020, 82, 49–58. [Google Scholar] [CrossRef]
- Colesie, C.; Walshaw, C.V.; Sancho, L.G.; Davey, M.P.; Gray, A. Antarctica’s vegetation in a changing climate. Wiley Interdiscip. Rev. Clim. Change 2023, 14, e810. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Distribution | Species | Habitat | Substrate Type | ||||
|---|---|---|---|---|---|---|---|
| Coastal | Forest | Alpine | Epiphytic | Saxicolous | Terricolous | ||
| bipolar | Acarospora badiofusca (Nyl.) Th. Fr. | + | + | ||||
| cosmopolitan | Agonimia tristicula (Nyl.) Zahlbr. | + | + | ||||
| bipolar | Amandinea punctata (Hoffm.) Coppins & Scheid. | + | + | + | |||
| bipolar | Arthrorhaphis citrinella (Ach.) Poelt | + | + | ||||
| endemic | Austroplaca ambitiosa (Darb.) Søchting, Frödén & Arup | + | + | ||||
| endemic | Austroplaca cirrochrooides (Nyl.) Søchting, Frödén & Arup s.lat. | + | + | ||||
| endemic | Austroplaca millegrana (Müll. Arg.) Søchting, Frödén & Arup s.lat. | + | + | ||||
| cosmopolitan | Baeomyces rufus (Huds.) Rebent. | + | + | ||||
| austral | Bibbya bullata (Meyen & Flot.) Kistenich, Timdal, Bendiksby & S. Ekman | + | + | + | + | ||
| bipolar | Caloplaca phaeocarpella (Nyl.) Zahlbr | + | + | ||||
| cosmopolitan | Candelariella vitellina (Ehrh.) Müll. Arg. | + | + | ||||
| bipolar | Carbonea vorticosa (Flörke) Hertel | + | + | + | |||
| bipolar | Catapyrenium cinereum (Pers.) Körb. | + | + | ||||
| bipolar | Cetraria aculeata (Schreb.) Fr. | + | + | ||||
| bipolar | Cetraria islandica (L.) Ach. | + | + | ||||
| bipolar | Chrysothrix chlorina (Ach.) J. R. Laundon | + | + | ||||
| austral | Cladia aggregata (Sw.) Nyl. | + | + | + | |||
| bipolar | Cladonia asahinae J.W. Thompson | + | + | + | |||
| bipolar | Cladonia bellidiflora (Ach.) Schaerer | + | + | + | |||
| bipolar | Cladonia borealis Stenroos | + | + | + | |||
| bipolar | Cladonia carneola (Fr.) Fr. | + | + | ||||
| endemic | Cladonia cervicornis spp. mawsonii (C.W. Dodge) S. Stenroos & Ahti | + | + | ||||
| cosmopolitan | Cladonia chlorophaea (Flörke ex Sommerf.) Spreng. | + | + | + | + | ||
| cosmopolitan | Cladonia cornuta (L.) Hoffm. | + | + | + | |||
| cosmopolitan | Cladonia fimbriata (L.) Fr. | + | + | ||||
| bipolar | Cladonia gracilis (L.) Willd. | + | + | + | |||
| endemic | Cladonia lepidophora Ahti & Kashiw. | + | + | ||||
| bipolar | Cladonia mitis Sandst. | + | + | + | |||
| endemic | Cladonia novochlorophaea (Sipman) Brodo & Ahti | + | + | ||||
| bipolar | Cladonia phyllophora Hoffm. | + | + | ||||
| bipolar | Cladonia pleurota (Flörke) Schaerer | + | + | ||||
| cosmopolitan | Cladonia pocillum (Ach.) Grognot | + | + | ||||
| cosmopolitan | Cladonia pyxidata (L.) Hoffm. | + | + | ||||
| bipolar | Cladonia rangiferina (L.) Weber ex Wigg. | + | + | + | |||
| austral | Cladonia sarmentosa (Hook f. & Taylor) C.W. Dodge | + | + | ||||
| bipolar | Cladonia scabriuscula (Delise) Nyl. | + | + | ||||
| cosmopolitan | Cladonia squamosa (Scop.) Hoffm. | + | + | ||||
| austral | Cladonia subsubulata Nyl. | + | + | ||||
| bipolar | Cladonia subulata (L.) F.H. Wigg. | + | + | ||||
| bipolar | Cladonia sulphurina (Michaux) Fr. | + | + | + | |||
| austral | Cladonia ustulata (Hook. F. & Taylor) Leight. | + | + | + | |||
| austral | Coccotrema cucurbitula (Mont.) Müll. Arg. | + | + | + | + | ||
| austral | Coelopogon epiphorellum (Nyl.) Brusse & Kärnefelt | + | + | + | + | ||
| bipolar | Cystocoleus ebeneus (Dillwyn) Twaites | + | + | + | + | ||
| bipolar | Frutidella caesioatra (Schaer.) Kalb | + | + | ||||
| austral | Gondwania sublobulata (Nyl.) S.Y. Kondr., Kärnefelt, Elix, A. Thell, J. Kim, M.-H. Jeong, N.-N. Yu, A.S. Kondr. & Hur | + | + | ||||
| bipolar | Gowardia nigricans (Ach.) Halonen | + | + | + | |||
| endemic | Haematomma erythromma (Nyl.) Zahlbr. | + | + | + | |||
| cosmopolitan | Hydropunctaria maura (Wahlenb.) C. Keller, Gueidan & Thüs | + | + | ||||
| bipolar | Hypogymnia lugubris var. lugubris (Pers.) Krog | + | + | + | |||
| bipolar | Japewia tornoensis (Nyl.) Tønsberg | + | + | + | |||
| bipolar | Lecanora epibryon (Ach.) Ach. | + | + | ||||
| cosmopolitan | Lecanora flotoviana Spreng. | + | + | ||||
| bipolar | Lecanora intricata (Ach.) Ach. | + | + | ||||
| endemic | Lecanora physciella (Darb.) Hertel | + | + | + | + | ||
| bipolar | Lecanora polytropa (Hoffm.) Rabenh. | + | + | ||||
| bipolar | Lecidea atrobrunnea (Ram. ex Lam. et DC.) Schaerer | + | + | ||||
| bipolar | Lecidella elaeochroma (Ach.) M. Choisy | + | + | ||||
| bipolar | Lecidella patavina (A. Massal.) Knoph & Leuckert | + | + | ||||
| bipolar | Lecidella stigmatea (Ach.) Hertel & Leuckert | + | + | ||||
| bipolar | Lecidella wulfenii (Hepp) Körb. | + | + | ||||
| bipolar | Lecidoma demissum (Rustr.) Gotth. Schneid. & Hertel | + | + | ||||
| bipolar | Lepraria caesioalba (B. de Lesd.) J.R. Laundon | + | + | + | + | + | |
| endemic | Leptogium menziesii (Ach.) Mont. | + | + | + | |||
| endemic | Leptogium puberulum Hue | + | + | ||||
| bipolar | Massalongia patagonica Kitaura & Lorenz | + | + | + | |||
| bipolar | Mastodia tessellata (Hook. f. & Harv.) Hook. f. & Harv. | + | + | ||||
| bipolar | Megalaria grossa (Pers. ex Nyl.) Hafellner | + | + | + | |||
| bipolar | Megaspora verrucosa (Ach.) Hafellner & V. Wirth | + | + | + | |||
| cosmopolitan | Melanohalea elegantula (Zahlbr.) O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. & Lumbsch | + | + | ||||
| endemic | Melanohalea ushuaiensis (Zahlbr.) O. Blanco et al. | + | + | + | + | ||
| endemic | Menegazzia magellanica R. Sant. | + | + | + | |||
| bipolar | Micarea incrassata Hedl. | + | + | ||||
| bipolar | Mycobilimbia hypnorum (Lib.) Kalb & Hafellner | + | + | ||||
| austral | Notoparmelia cunninghamii (Cromb.) A. Crespo, Ferencová & Divakar | + | + | ||||
| endemic | Ochrolechia antarctica (Müll. Arg.) Darb. | + | + | + | |||
| bipolar | Ochrolechia frigida (Sw.) Lynge | + | + | + | |||
| bipolar | Pannaria hookeri (Borrer ex Sm.) Nyl. | + | + | + | |||
| cosmopolitan | Parmelia saxatilis (L.) Ach. | + | + | + | |||
| cosmopolitan | Peltigera didactyla (With.) J.R. Laundon | + | + | ||||
| cosmopolitan | Peltigera rufescens (Weiss) Humb. | + | + | ||||
| endemic | Peltularia fuegiana Henssen & P.M. Jørg. | + | + | + | + | + | |
| endemic | Pertusaria spegazzinii Müll. Arg. | + | + | ||||
| bipolar | Phaeophyscia endococcina (Körb.) Moberg | + | + | ||||
| cosmopolitan | Physcia caesia (Hoffm.) Fürnr. | + | + | ||||
| cosmopolitan | Physcia dubia (Hoffm.) Lettau | + | + | ||||
| bipolar | Physconia muscigena (Ach.) Poelt | + | + | ||||
| cosmopolitan | Placidium squamulosum (Ach.) O. Breuss | + | + | ||||
| endemic | Poeltidea perusta (Nyl.) Hertel & Hafellner | + | + | + | + | ||
| bipolar | Polycauliona candelaria (L.) Frödén, Arup & Søchting | + | + | + | + | + | |
| bipolar | Pseudephebe minuscula (Nyl.) Brodo & D. Hawksw. | + | + | ||||
| endemic | Psoroma antarcticum Hong & Elvebakk | + | + | ||||
| endemic | Psoroma cinnamomeum Malme | + | + | + | + | ||
| austral | Psoroma fruticulosum James & Henssen | + | + | ||||
| cosmopolitan | Psoroma hypnorum (Vahl) Gray | + | + | ||||
| endemic | Ramalina terebrata Hook. & Taylor | + | + | ||||
| bipolar | Rhizocarpon geminatum Körb. | + | + | + | |||
| bipolar | Rhizocarpon geographicum (L.) DC. | + | + | + | + | ||
| bipolar | Rhizocarpon polycarpum (Hepp) Th. Fr. | + | + | ||||
| bipolar | Rinodina olivaceobrunnea C.W. Dodge & G.E. Baker | + | + | + | |||
| austral | Rinodina peloleuca (Nyl.) Müll. Arg. | + | + | ||||
| cosmopolitan | Sarcogyne privigna (Ach.) A. Massal. | + | + | ||||
| austral | Siphulastrum mamillatum (Hook. F. & Taylor) D.J. Galloway | + | + | ||||
| cosmopolitan | Sphaerophorus globosus (Huds.) Vainio | + | + | ||||
| bipolar | Sporastastia testudinea (Ach.) A. Massal. | + | + | ||||
| bipolar | Stereocaulon alpinum Laurer | + | + | + | |||
| austral | Stereocaulon glabrum (Müll. Arg.) Vain. | + | + | ||||
| cosmopolitan | Tephromela atra (Huds.) Hafellner | + | + | ||||
| bipolar | Toniniopsis bagliettoana (A. Massal. & De Not.) Kistenich & Timdal | + | + | ||||
| bipolar | Trapeliopsis granulosa (Hoffm.) Lumbsch | + | + | ||||
| bipolar | Tremolecia atrata (Ach.) Hertel | + | + | ||||
| bipolar | Umbilicaria decussata (Vill.) Zahlbr. | + | + | ||||
| bipolar | Umbilicaria nylanderiana (Zahlbr.) H. Magn. | + | + | ||||
| bipolar | Umbilicaria umbilicarioides (Stein.) Krog & Swinscow | + | + | ||||
| austral | Usnea aurantiaco-atra (Jacq.) Bory | + | + | ||||
| bipolar | Usnea cf. sphacelata R. Br. | + | + | + | + | + | |
| endemic | Usnea subantarctica F.J. Walker | + | + | ||||
| austral | Usnea trachycarpa (Stirton) Müll. Arg. | + | + | ||||
| endemic | Verrucaria dispartita Vain. | + | + | ||||
| austral | Verrucaria durietzii I.M. Lamb | + | + | ||||
| bipolar | Verrucaria halizoa Leight. | + | + | ||||
| bipolar | Verrucaria margacea (Wahlenb. in Ach.) Wahlenb. | + | + | ||||
| austral | Verrucaria tesselatula Nyl. | + | + | ||||
| cosmopolitan | Xanthoria elegans (Link) Th. Fr. | + | + | ||||
| Morphology | Absolute Abundance | Relative Abundance |
|---|---|---|
| crustose | 56 | 45.2% |
| foliose | 28 | 22.6% |
| fruticose | 16 | 12.9% |
| cladonoid | 24 | 19.3% |
| Habitat | Substrate Type | Distribution | |||
|---|---|---|---|---|---|
| Endemic | Austral | Bipolar | Cosmopolitan | ||
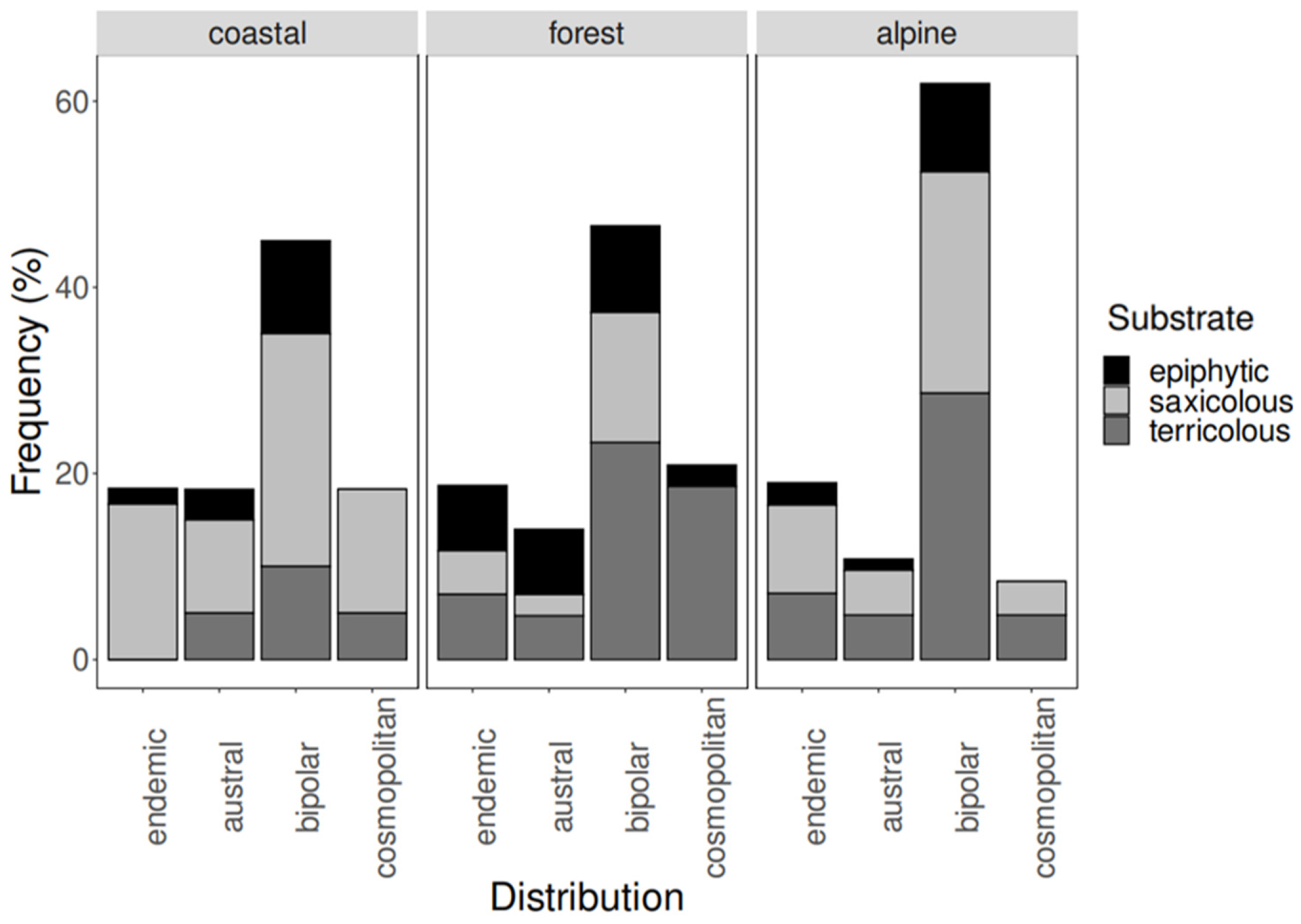
| coastal | epiphytic | 1.7% | 3.3% | 10.0% | 0.0% |
| saxicolous | 16.7% | 10.0% | 25.0% | 13.3% | |
| terricolous | 0.0% | 5.0% | 10.0% | 5.0% | |
| forest | epiphytic | 7.0% | 7.0% | 9.3% | 2.3% |
| saxicolous | 4.7% | 2.3% | 14.0% | 0.0% | |
| terricolous | 7.0% | 4.7% | 23.3% | 18.6% | |
| alpine | epiphytic | 2.4% | 1.2% | 9.5% | 0.0% |
| saxicolous | 9.5% | 4.8% | 23.8% | 3.6% | |
| terricolous | 7.1% | 4.8% | 28.6% | 4.8% | |
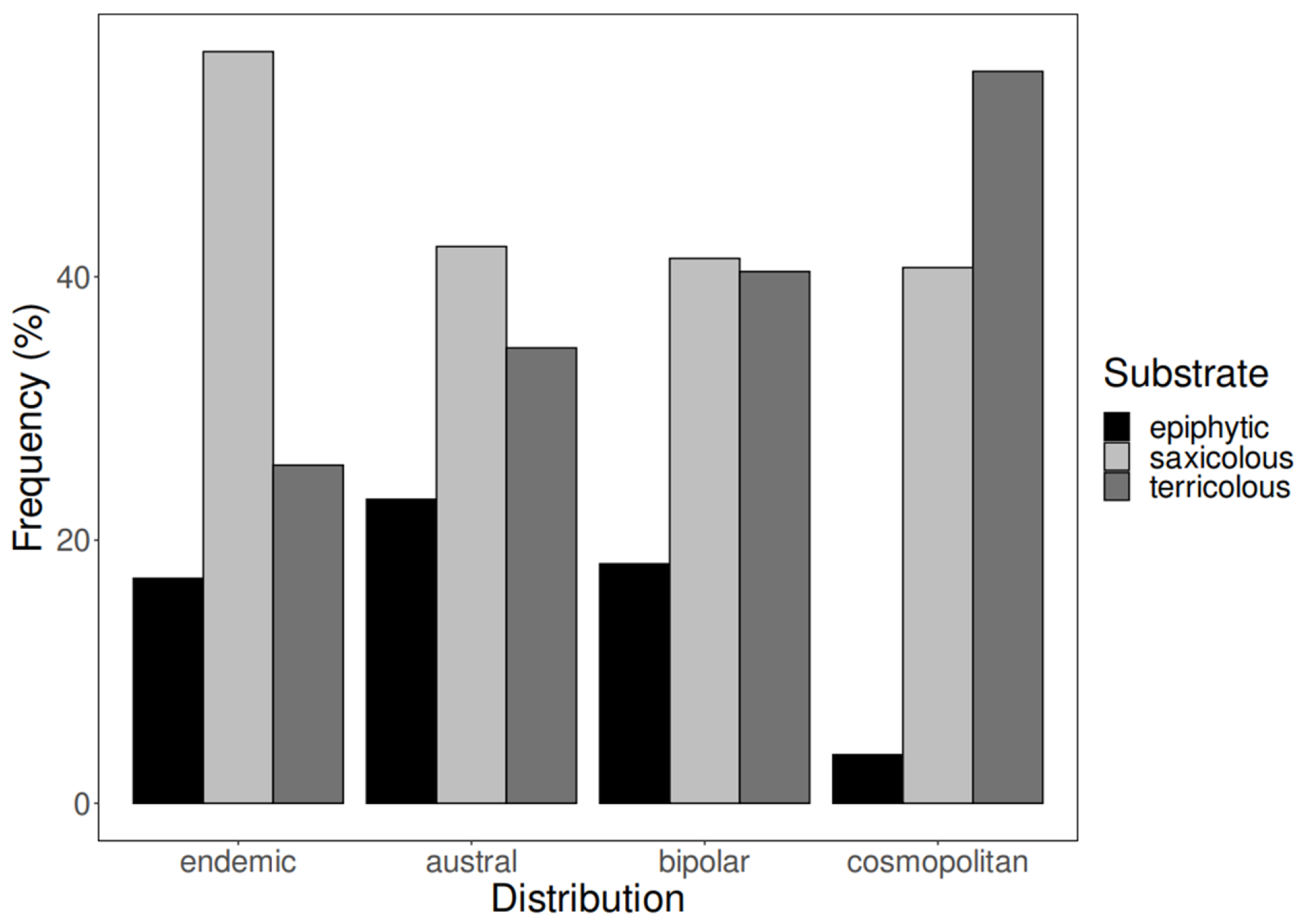
| Substrate Type | Habitat | ||
|---|---|---|---|
| Coastal | Forest | Alpine | |
| epiphytic | 15.0% | 25.6% | 13.1% |
| saxicolous | 65.0% | 20.9% | 41.7% |
| terricolous | 20.0% | 53.5% | 45.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sancho, L.G.; Aramburu, A.; Etayo, J.; Beltrán-Sanz, N. Floristic Similarities between the Lichen Flora of Both Sides of the Drake Passage: A Biogeographical Approach. J. Fungi 2024, 10, 9. https://doi.org/10.3390/jof10010009
Sancho LG, Aramburu A, Etayo J, Beltrán-Sanz N. Floristic Similarities between the Lichen Flora of Both Sides of the Drake Passage: A Biogeographical Approach. Journal of Fungi. 2024; 10(1):9. https://doi.org/10.3390/jof10010009
Chicago/Turabian StyleSancho, Leopoldo G., Ana Aramburu, Javier Etayo, and Núria Beltrán-Sanz. 2024. "Floristic Similarities between the Lichen Flora of Both Sides of the Drake Passage: A Biogeographical Approach" Journal of Fungi 10, no. 1: 9. https://doi.org/10.3390/jof10010009