5.1. General Considerations
Numerous papers have been published in the resistance field; however, in the maize genetic conference series, only several papers were published in this subject. In 2005, none of the 32 talks and only 3 of the 248 posters were connected to toxigenic fungi [
103]. In 2006, 258 posters and 37 talks were given, of which 2 mentioned Fusarium and 5 aflatoxin [
104]. In 2008, the abstracts of 54 talks and 241 posters contained no paper considering Fusarium or aflatoxin [
105]. In 2022, out of 27 talks and 219 posters, 2 focused on Fusarium and none considered aflatoxin [
106]. Although not all conferences are cited here, these few clearly demonstrate that the significance of toxigenic fungi is fairly low in the genetic field. The real situation is possibly not so bad. Moretti et al. [
107] analyzed the relationships between climate change (mostly warming) and mycotoxin-related problems, forecasting the increase in aflatoxin in South, North, and Middle Europe. In terms of Fusarium mycotoxins, fumonisins and DON are expected to be the most prominent, but plant breeding for resistance and better adaptation were not mentioned among the most important practices to balance the harmful effects of climate change on food safety. The study of Dovenyi-Nagy et al. [
108] is an exception, as they discussed the roles of resistance breeding in terms of reducing aflatoxin contamination in the field. Tirado et al. [
109], in their review, mentioned mycotoxins in one line, and, as an initiative, only worldwide mycotoxin regulations were mentioned in terms of helping to control food safety standards; however, this cannot solve the problem. Other authors [
110,
111] have responded similarly.
Resistance can be grouped into the physiological and physical (morphological) resistance of the maize grains [
8]. The literature references the influence of physical factors, such as husk leaves, ear angle, kernel characteristics, dehydration rate, and others [
112]; general experience suggests that these traits alone can lower the risk but are not capable of solving the overall problem. More importantly, for example, the thickness of the seedcoat and its waxiness, which are genetically determined, can have a significant effect in protecting grains against FER infection [
113,
114]; however, what is the relative case for
F. verticillioides and
A. flavus? The husk leaves play two key roles. When they remain closed until ripening, the higher humidity under the husk leaves can support the spreading of fungi. Thus, when the husk leaves open and dry-down can be accelerated, this indirectly inhibits fungal growth by decreasing the water activity (a
w) and, thus, the chance of infection. However, a vertically standing ear with opened husk leaves can lead rain into the ear, which can significantly increase the water content of the grains and support increased toxin contamination. The husk leaves on bowing ears protect the grains like an umbrella, and the water uptake of the grains remains moderate or close to zero. The dehydration rate of the kernel (drydown) also has an influence on ear rot and mycotoxin accumulation; for a long time, this has been considered an important breeding task to lower the dehydration cost after harvest. The largest problem seems to be related to the lack of new findings regarding the significance of physical barriers. Without any facts, we cannot hope to make any significant progress. It is understood that physiological dry-down and pathological dry-down following stalk rot inhibit fungal spreading [
115]. The latter causes a pseudoresistance to ear rot that, in a rainy season, will not be effective and supports epidemics [
115,
116]. For this reason, the development of stalk rot should be inhibited, as ear rot—particularly that caused by
F. graminearum—can give genetically valid results when stalk rot is absent or, on the scale of 0–5, only a rating of 1 is acceptable. This can be achieved through lower stand density, irrigation, or a combination of both. For this reason, in our tests, stalk rot is always checked. For ear rots caused by the much less pathogenic
F. verticillioides and
A. flavus, no data could be found, possibly because their influence on the grain yield is low [
15]. The author would like to see more papers in this field, as it is not an accident that a vast majority of publications have utilized artificial inoculation methods. The argument that artificial inoculation inhibits the effect of the physical protection is not well founded, as toothpick or silk channel inoculation will not lead to spreading in a resistant plant, while 70–90% infection severity can be achieved in a highly susceptible plant. Therefore, the identification of the resistance is possible, allowing for the selection of the highly resistant hybrids [
10]. It can be supposed that, at a high resistance level, the influence of physical resistance or susceptibility is much lower. As physical traits also have a genetic background, as much so as their physiological background, the differences between them should not be stressed but, rather, their combined application is suggested (when supported by solid scientific evidence).
The term “physiological resistance” could be changed, but its use should be avoided; for example, the waxiness of the seedcoat is produced by different genetic and physiological processes influencing plant behavior. The latest genetic research has described the resistance to toxigenic fungi in cereals as quantitatively inherited resistance (QDR)—meaning polygenetically determined traits [
117], as, until now, no evidence has shown monogenic traits to secure high levels of resistance alone. It should be noted that most of the resistance genes again rust diseases identified in wheat determine only a moderate susceptibility or resistance level, and only the most effective can provide field immunity [
118]. Furthermore, race nonspecific resistances governed by QTLs have also been determined with regard to rust disease in wheat [
119]. Considering the two resistance types, the common factor is that both can determine partial resistance resembling QDR [
117].
Munkvold [
120,
121] has summarized research from the USA and stated that Fusarium ear rot has not been reported in the literature, as found for
Gibberella zeae and
A. flavus. Since that the situation changed, the number of FER publications surpasses GER papers and is comparable with that of
A. flavus. Hung et al. [
122] added that all significant breeding activities were concentrated into the private industry, and, while the genetic basis was better explained, screening methods were found to be less adequate. This was the first summary considering QTL identification, and the polygenic nature of resistance was well supported. However, at this time, resistance to disease and toxin contamination was not mentioned as a separate problem. Munkvold [
123] stressed the need for the appropriate selection of hybrids and found it highly important to understand the pathogen composition in a region, as the resistance relations and their validity may change outside the region. To date, the knowledge on FER has improved significantly, but the aflatoxin contamination has become more problematic, partly due to the warmer and dryer seasons. In addition, the lack of reliable data on resistance greatly inhibits both production and research.
Not accidentally, researchers have observed rather good correlations between the amount of the disease (FER) and the toxin contamination in several tests [
124]. Functional genomics may help to identify further resistance paths against
F. verticillioides [
30]. Santiago et al. [
29] summarized that the symptom severity in resistance tests correlates well with lower toxin contamination; however, they stated that direct screening for lower mycotoxin contamination is expensive and time-consuming. A key problem in this context is that the data typically come from experiments on inbreds [
17,
32,
33]. For this, we need combining ability for grain yield and disease, as well as resistance to toxin accumulation that normally does not exist. For this reason, only hybrid resistance tests can advise us correctly.
It is widely accepted that resistance level and toxin contamination are closely related; thus, it is sufficient to select for high resistance to achieve low toxin contamination [
29]. Eller et al. [
125] stated that direct selection for toxin contamination is very expensive and, therefore, is not suggested. The highest infection was found six days after silking [
48]. A key problem is that, without ear rot-severity data, the toxicity data cannot be validated. We previously detected a more than ten-fold difference in mycotoxin contamination for 1% of FER, GER, and AER infection severity [
51] (
Table 4), but it is not possible to forecast toxicity for a significant number of genotypes. As we do not know which hybrid will react in a certain way, toxin measurement is necessary. An important aspect of toxin production is that the percentage of visual infection may present very large differences. For a natural infection, the case is similar [
10] for a percentage of ear infection, the DON content ranged between 0 mg/kg and 7.63 mg/kg for 2017/2018 and 0 mg/kg and 46.5 mg/kg for 2019/2020. The FUM content ranged between 0 mg/kg and 6.31 mg/kg for 2017/2018, where 1.2 and 58.2 mg/kg were the minimum and maximum rates, respectively. Aflatoxin ranged between 1.10 mg/kg and 63.5 mg/kg for 2017/2018 and 0–6439 mg/kg for 2019/2020 [
10]. These differences revealed no significant correlations. The case is similar when conducting artificial inoculation (
Table 4). The rate of maximum and minimum values was the lowest for DON (8.63-fold), medium for fumonisins (17.61-fold), and highest for aflatoxin B1 (58-fold). This serves as an argument to not predict toxin contamination based on visual infection.
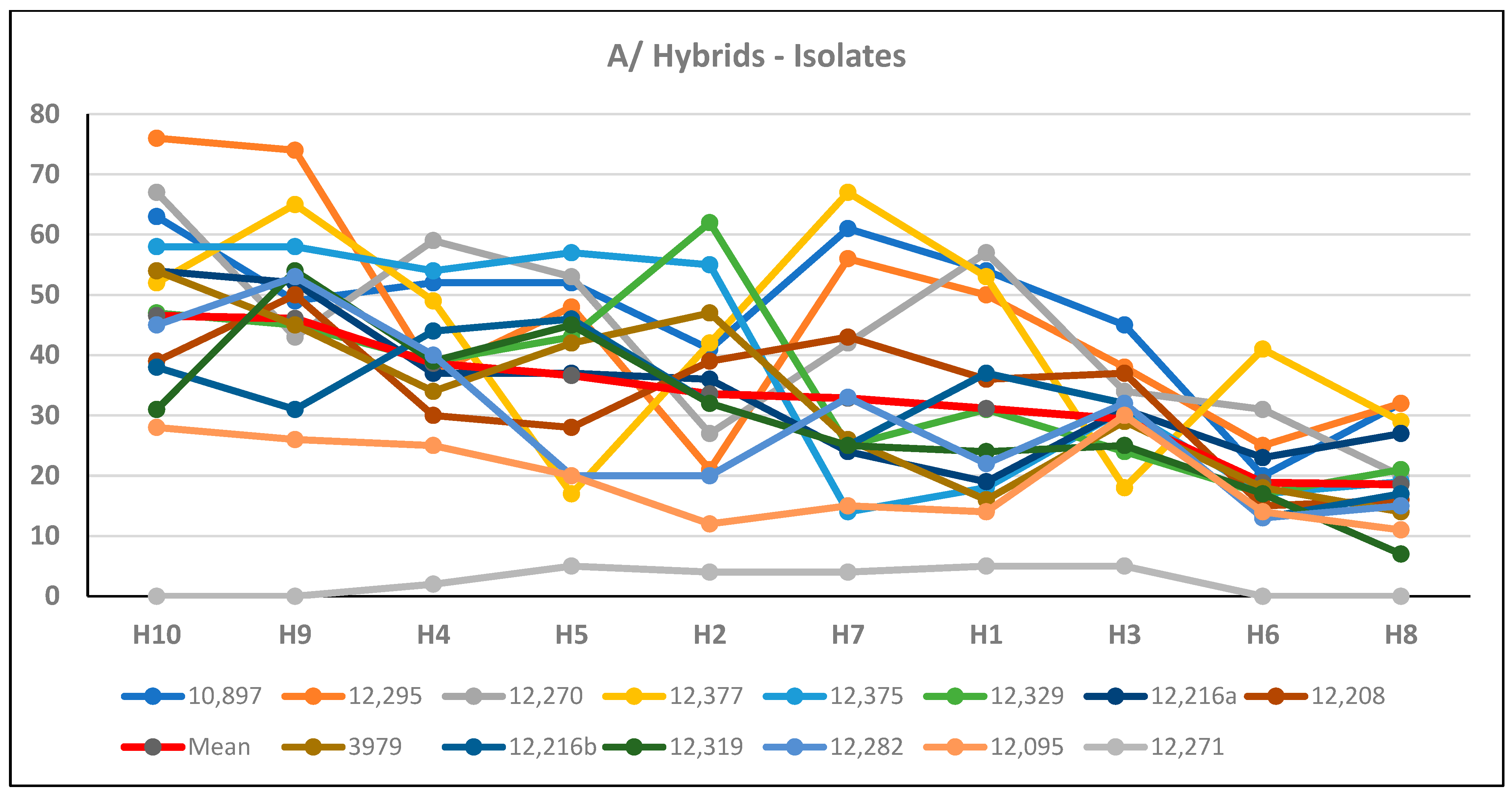
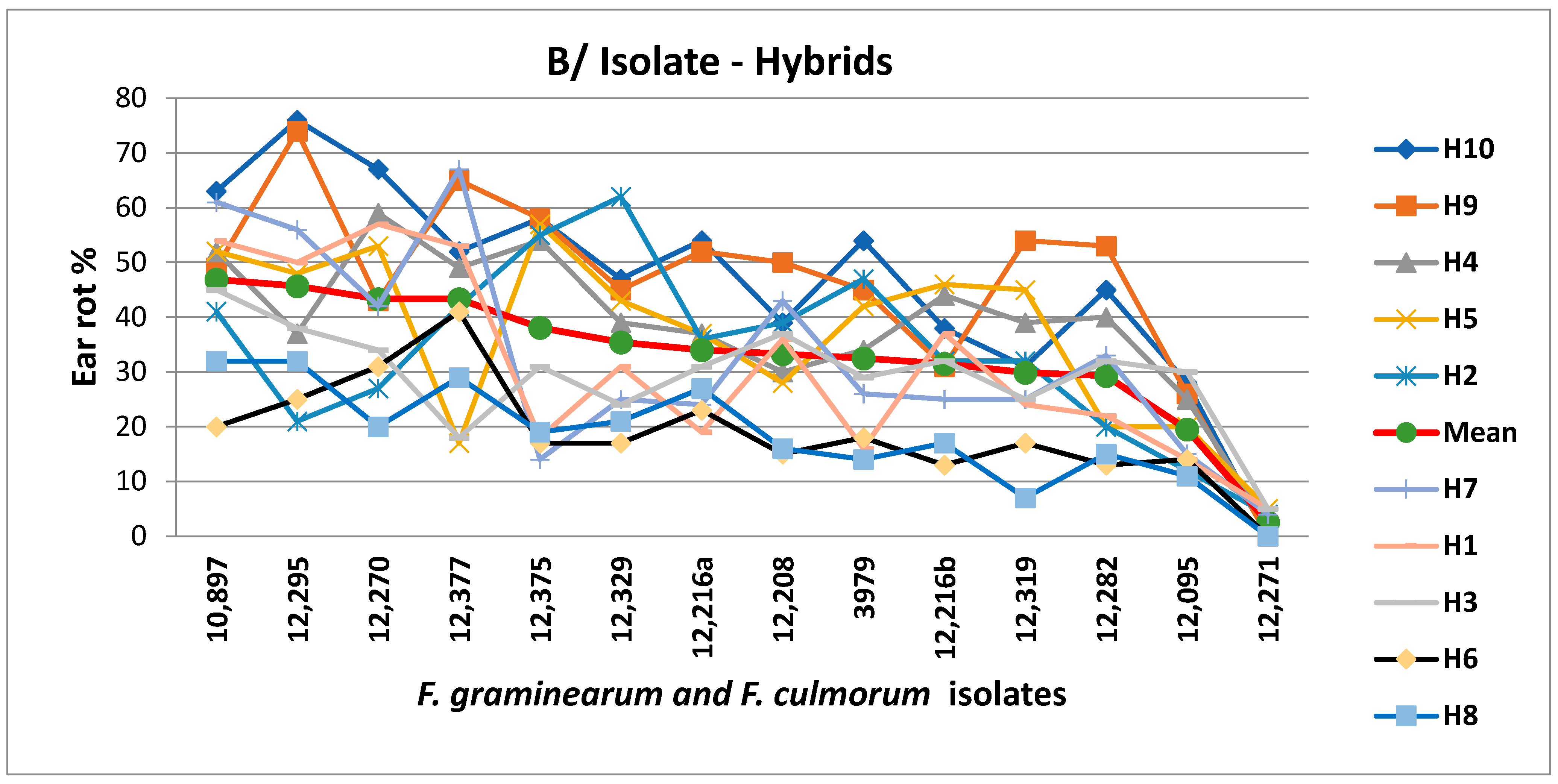
Looking at the variances (this serves as a stability index) with regard to different toxins, large differences between hybrids were found [
51]. Several gave a very uniform performance, and some others reacted very differently in the two years to the two isolates. So, hybrids with stable performance to different toxins and their producers could be identified. The summary table (
Table 5) shows all data of the hybrids that are important for the risk analysis [
51]. Among them, four hybrids were identified for which all of the data were in the low and low–medium groups. Only one hybrid had the worst (orange) category for all toxins, while the rest varied. So, without any logical order (e.g., from the DON response), no conclusions could be drawn for the other hybrids. Of course, the original data were also important in performing a risk analysis, as they reveal the causes of the correlation-breaking genotypes; however, even the specific DON-regulating agents remained unclear. Regardless, a risk analysis can still be conducted. On the other hand, such research can help to clarify the causes of the phenomenon.
In both research and variety registration, toxin analyses cannot be spared. Through the breeding process, we have the possibility to reduce the number of toxin tests. When a hybrid is susceptible, it can be discarded without a toxin test. Most importantly, those hybrids that have good resistance, but high toxin contamination are dangerous. For example, in 2017/2018, Korimbos and DKC 4590 had 0.04% and 0.36% FER severity at 0.81 and 2.31 mg/kg FUM, 0.05% and 0.15% AER severity at 298 and 2150 mg/kg AFB1, and 8.48% and 11.18% for GER with 26 and 9 mg/kg DON. At the same time, the naturally infected controls of several hybrids showed no visible fungal presence for AFB1 and presented 32 mg/kg contamination. Similar examples were also observed in the 2019/2020 hybrid tests, and, under natural infection, several hybrids produced 350–450 mg/kg aflatoxin or higher without visible infection. The extent to which such results are environmentally dependent remains to be determined. We concluded that the associated relationships are much more complicated. Even the resistance and toxin data correlations were variable for the different ear rots; they allow us to identify superior genotypes for good performance to all diseases and the accumulation of toxins [
10,
50,
52,
126].
5.2. Gibberella Ear Rot
GER is favored by higher precipitation during silking, followed by moderate temperatures, and high rainfall in maturing period, as occurred in 2014 in Hungary [
10,
120]. The causing agent,
F. graminearum, is highly pathogenic. Its isolates are mostly highly aggressive; therefore, it is easier to work with in a breeding program than the much less pathogenic
F. verticillioides or the even less pathogenic
A. flavus [
51,
52]. The susceptibility window is about 6 days during silking [
44,
45]. Miller et al. [
127] presented data on the process of silk infection. The cob was accessed in 5–7 days in susceptible genotypes but required 12–15 days in resistant genotypes. Entry into the ear was through the surface of the rachis, via the exterior growth of the grains or in the rachis through the pedicel. This would mean that inoculation is ideal at between 5 and 7 days after mid-silking, allowing for better a differentiation of the resistance level. The subjects of the tests were very early hybrids, as they are in commercial production, and their resistance and toxin risks have direct economic consequences [
47]. They pointed out that less than 10% of the tested hybrids presented a stable performance. Our recent results [
50,
51,
52] fully support this conclusion. The inheritance of the GER resistance had the highest rate for GER symptoms (17% increase across 27 hybrids). For DON, the mean heterosis was −35%; e.g., the majority of the hybrids had a higher DON content than the mean of the parents, and only seven hybrids were better than the parental mean was. So, the heterosis in symptoms will not agree with the DON contamination of the hybrids; the case seems to be more complicated than supposed [
126].
QTL research. Ali et al. [
40] identified 11 QTLs for
F. graminearum severity for ear rot resistance following silk inoculation, as well as 18 QTLs in kernel resistance, which accounted for 6.75–35% of the total variation identified. Only two QTLs were detected in more than two environments for silk resistance, and only one for kernel resistance. The disease evaluation followed Reid et al. [
44,
128]. The authors explained this result in terms of the high influence of the environment on the expression of resistance. In the author’s opinion, this might be true; however, the facts that all QTLs had a small effect and that the total variation (even in the best case) was only one-third of the total variation make it likely that there may have been problems in the testing methodology. The 1–2 QTLs that were active in all environments could make a large improvement in the resistance when used, while controlling only a small percentage of the total variation. In the test, no toxin contamination assessment was conducted, and, so, it is not known how far these resistance results may contribute to lower mycotoxin (e.g., DON and ZEN) contamination. Four QTLs were identified by Galiano-Carnerio et al. [
42], explaining 5.4–21.8% of the genotypic variance. Of these, only one was stable across all environments. Toxin data were not included.
Giorni et al. [
129] identified four QTLs in the LP4637/L4674 population (No. 298 RIL lines), explaining 11.2–11.8%, 3.4–5.1%, 6.2–7.6%, and 3.8–5% of the total variance (24.6–29.5%). This may be consistent with other sources, but the effectiveness of a breeding program that can maintain only 25–30% of the resistance is questionable, as this means that we do not have an explanation for 70–75% of the variance observed. The validation of QTLs is a general problem: out of five identified QTLs, none was stable, and the validation rates were disappointingly low, indicating GER resistance arising from many low-effect QTLs [
130]. A total of 140 RIL genotypes have were tested [
43] for silk and kernel resistance. These RILs showed highly significant differences. The QTLs were co-localized in chromosomes 1, 2, and 8 and differed in chromosomes 9 and 10. Chr. 9 seems to have an influence on silk resistance, while Chr. 10 seems to be specific to kernel resistance. As the data did not present any correlation with agronomic data, the grain yield response seems to be independent (only weak correlations) from GER resistance. As the disease severity did not differ significantly, the differences were not derived from the differences in aggressiveness between the two inoculation methods. Zhou et al. [
46] identified 11 QTLs, 5 of which were stable. The inheritance was additive + epistatic, and no one QTL explained more than 10% of the phenotypic variation; furthermore, no toxin measurements were conducted.
Yuan et al. [
131] were among the first to identify a putative resistance gene (guanylyl cyclase-like gene) against
F. graminearum in maize. Reid et al. [
44] tested the correlation between visual scores and DON contamination in hybrids. Ear rots higher than 25% GER contained high levels of DON. They concluded that visual symptoms should be suitable for breeding resistance to lower toxin accumulation and that no specific toxin measurements are necessary; this can be valid for this test, as we have cases for good agreement and no significant correlation, too. The problem is that high DON contamination can also be detected at 10% infection severity or lower. Tests with updated methodologies and toxin sampling showed that, depending on the experiment, 20–30% of the hybrids reacted differently, presenting higher (DON overproduction) or lower (relative DON resistance) DON contamination than that forecast by the regression function [
10,
48,
52]. This holds also for the natural infection, but the correlations were even lower and mostly not significant [
51]. We found hybrids presenting 25.8 mg/kg DON at 8.58% GER and 10.84 mg/kg DON at 12.33% GER in 2017/2018. Among the genotypes for 2019/2020 in the same paper, we observed a hybrid with 13.50% GER and 7.48 mg/kg DON, and another with 14.35% GER and 76.08 mg/kg DON. Similar deviations were found in a combining ability test of maize inbreds for GER and DON contamination, and the results indicated a more complicated situation than supposed by the literature result cited above [
126]. The GER and DON presented a correlation of only r = 0.54. For the hybrids, 19 reacted similarly with regard to GER and DON, while 8 gave very variable responses, indicating a much more complicated inheritance mechanism. Another study [
132] identified 4–6 QTLs for GER resistance and resistance to toxin accumulation in a mapping population of two inbreds, which explained 29–35% of the phenotypic variance. While the authors [
132] supposed a pleiotropic effect, the explanation may be different: when resistance and toxin contamination are closely correlated, it means that a lower infection severity automatically means lower DON contamination or higher severity with higher DON contamination, without any pleiotropic effect. When we tested hybrids with highly different origins, the picture differed, and a larger range of variation was observed [
51,
52]. Genome-wide association studies and genomic selection have also been conducted in order to utilize the higher resistance found often in landraces. Considering GER severity, eight QTLs (explaining 34% of the genetic variation) were identified in the Kemater Landrace Gelb which presented no significant correlations with agronomic traits such as days to silking and seed set [
60]; this is good news in terms of breeding for higher grain yield in all ripening classes.
Other approaches might also be interesting. Geranic acid has been found to act as an antifungal agent against
F. graminearum in maize. The authors produced transgenic maize plants via geraniol synthase. They observed a geraniol dihexose and four different hydroxyl-geranic acid-hexoses in the transgenic plants; however, these plants were not more resistant than the control material [
133]. A maize defensin peptide, PDC1, which is expressed in
Escherichia coli, has been assessed in maize against GER. The peptide restricted fungal growth and disease severity, but the influence on toxin content was not investigated [
134].
It seems that resistance to GER is more complicated than previously thought. The generally accepted excellent correlation between visual symptoms and DON contamination is not generally valid, with a smaller or higher part of the tested population not fitting the assumptions. For this reason, DON measurements cannot be excluded. No toxin measurements were made in many studies; therefore, the food safety risk of the hybrids cannot be reliably evaluated. Food safety problems are caused not by DON but, instead, its masked and acetylated versions, which are normally not controlled. Zearalenone data are nearly absent; however, it can be considered important to understand how resistance to DON accumulation corresponds to ZEN data. We should mention nivalenol, which has been found to be produced by isolates of
F. graminearum,
F. culmorum,
F. asiaticum, and other
Fusarium spp. As it is ten-fold more toxic than DON, its food safety risk may be highly significant. QTL analyses allow for one certain conclusion: the trait is polygenic, and the explained variability is seldom higher than 30%. This means that methodical problems may also contribute to this phenomenon, as evidenced in wheat [
135,
136].
5.3. Fusarium Ear Rot
For FER, the optimum temperature is 30 °C. In order to develop, it needs warm and dry weather during grain filling. Drought stress increases disease severity. The silking period also provides higher disease incidence and severity [
120,
137]. Significant resistance differences to FER have been documented in a long list of papers. In the test inbreds, hybrids were tested per se, but genetic studies between susceptible and resistant inbreds have been conducted using many different methods. Inbreds are generally more susceptible than hybrids and show highly significant resistance differences. In 2009, 0.56 and 240 mg/kg of fumonisin were measured by Balconi et al. [
138], but the correlations between symptom severity and fumonisin content were only r = 0.37 and r = 0.29. On this basis, the prediction of fumonisin production based on symptom severity does not seem to be very promising. Significant differences have also been found for hybrids [
10,
47,
50,
52]. Fumonisin production co-segregated with the aggressiveness of
F. verticillioides isolates, as was observed for DON in GER infection [
139]. As the aggressiveness of
F. verticillioides isolates is generally low, preselection of the isolates for artificial inoculation is necessary [
50,
52]. From a methodological point of view, it is interesting to note that lower and higher FB1 + 2 concentrations were found in high- and medium-rainfall zones in Zambia, respectively, without correlations with years and precipitation. In South Brazil, this is the ruling species (Tessmann, pers. comm., 2023). The planting time was neutral. Consequently, high- and low-resistance hybrids were not found under natural infection; however, under artificial inoculation, significant differences between hybrids were identified [
34].
To assess the resistance to FER, Araujo et al. [
15] compared three inoculation methods: aspersion (spraying), injection, and natural infection. In two locations, the control and inoculated variants did not differ significantly while, in the two other locations, the artificial inoculation was stronger, but the results differed. Differentiation of resistance was better in the regions with a higher temperature. It seems that the female flowering and drying of the grains were the most sensitive phases in terms of infection and fumonisin synthesis [
20,
29]. For the latter, insect wounds were mostly found to be responsible. Therefore, in resistance tests, insect-damaged ears should be discarded. It seems that the dent stage is critical, and the highest fumonisin accumulation is when just reaching full ripening; here, no insect is needed. The role of husk coverage from earlier papers was supported here [
140]. A non-excessive pericarp thickness also helped to reduce the fumonisin level. The structure of the pericarp and its wax layer can act as resistance factors protecting maize against
F. verticillioides and fumonisin contamination, as removing the wax layer increased the infection and FUM contamination severity [
113,
114,
141,
142]; furthermore, the diferulate concentration appeared to be proportional to the fumonisin contamination (R
2 = 0.82). Ivic et al. [
28] could not prove this role in hybrid tests. Both may be true, and we observed this often when testing different hybrid groups [
51,
52]. Munkvold [
120] reported on significant resistance differences with respect to FER, but the genotype responses differed in the network of countries and CIMMYT. Afolabi et al. [
14] reported a significant correlation between the incidence of ear rot and fumonisin contamination (r = 0.39 and 0.35 in two locations;
p = 0.0001), but they stressed that only five inbreds were found that produced consistently low fumonisin contamination (lower than 5 mg/kg) across years and locations. This agrees well with our data [
10,
50,
52]. These inbreds were successfully used in a breeding program.
This is important,
as these authors were among the first who stressed the significance of a stable low toxin response, and their practical significance could also be verified.
The combining ability is an important trait, and additive and non-additive gene effects should be identified [
143]. Hung and Holland [
122], in a diallel analysis, observed a 27% decrease in infection severity and 30% reduction in fumonisin content, compared to the parental inbreds. The general combining ability (GCA) and specific combining ability (SCA) were closely correlated (r = 0.78 or higher). Morales et al. [
144] evaluated external and internal infection, as well as FUM contamination. The four families provided a large variability in response, and genotypes with lower kernel density and larger cobs had higher FUM. It seems that QTLs could be classified as putatively resistance-specific and putatively for ear and resistance traits. This result supports the different QTL functions observed in wheat [
135,
136]. This method is used mostly for grain yield breeding and is seldom used in resistance breeding. However, more recently, molecular methods are more commonly used, but the significance of the combining ability has remained important.
QTL analyses. Resistant and susceptible lines were used to create a mapping population, and three QTLs were detected on chromosomes 4, 5, and 10. Of these, Chr. 4 explained 18% of the 90% higher infection severity variation and, so, was selected for further work by Chen et al. [
145]. No fumonisin tests were conducted. Wu et al. [
35] analyzed FER resistance by linkage mapping and GWAM. A total of 10 QTLs were identified that explained 1.0–7.1% of the phenotypic variance. Epistatic mapping was better, which explained 21–30% of the phenotypic variance. As such, multiple genes were identified with minor effect. No toxin tests were conducted. Giomi et al. [
20], through their QTL analysis, could explain 56–58.2% of the observed variation; this is among the highest that we found. Cao et al. [
21], from 364 differentially expressed genes (DEGs), identified four QTL genomic regions against FER which were active with respect to disease and toxin contamination resistance. A positive development is that gene functions could also be connected to these results, such as those related to cell wall biosynthesis and flavonoid biosynthesis. This is important, as the possibility to identify genes can play a decisive role in resistance development and expression. Another possible gene group in this line is connected to the salicylic acid and steroid signaling system in silks [
146]. Butron et al. [
18] detected 13 putative QTLs with respect to FER, located on all chromosomes except Chr. 5, all of which were identified as small-effect QTLs. Of them, two seemed to have an influence on FER and fumonisin contamination when inoculation tests were conducted without replications (in each block of 42 genotypes, supported by eight lines from the MAGIC population as controls); their reliability should not be overestimated. The rest varied; for example, QTLs with different functions were found. Data were transformed using the log
10(
p) method. One should be cautious when drawing far-reaching conclusions from non-replicated trials. Perez-Brito et al. [
147] could explain only 44% of the variability according to the QTLs identified; in retrospect, this is not a bad result, and most of the wheat QTL analyses have not provided significantly better results.
SNP and GWAS analyses. Genome-wide association mapping in 818 tropical inbred lines identified 45 SNPs and 15 haplotypes connected to FER resistance with 1–4% individual effect and of additive character. Altogether, five QTLs were identified in Population 1, which explained 49% of the total variance. For Population 2, six QTLs explained 25% of the phenotypic variance [
22]. They concluded that the value of the identified QTLs was limited, indicating a polygenic inheritance which differs from population to population. They stated that a combination to select with general adaptation may help in increasing FER resistance. It should be noted that toxin contamination was not analyzed; therefore, the food safety aspect of the results is unclear. They applied the kernel resistance test (nail punch/sponge inoculation) described by Drepper and Refro [
148]. Another study [
36] revealed seven SNP variants across 556 hybrids associated with 1–3% of the phenotypic variation; however, FUM was not controlled. Santiago et al. [
149] reported about 23 mapping populations on FER. Visual symptoms were analyzed in 19 populations only, and only 4 related visual symptoms to fumonisin contamination. This is very important, as breeders can better consider the fumonisin-producing capacity and differentiation of the breeding material. At this time (2020), no genes were known to underlie the QTLs. However, their data strongly supported the view that more effective breeding against FER is possible. Genome-wide association mapping (GWAS) is a new technology which can be conducted on hybrids and inbreds. As such, a large amount of material can be screened in a relatively short time. Coan et al. [
150] identified 14 SNPs, among which four genes were identified as defense-related proteins. They explained 15–25% of the phenotypic variance, but none was stable across the three environments tested. Zila et al. [
151] reported the GWAM results of 267 inbred lines in two environments. Eight SNP loci were identified with small additive effects, explaining 3–12% of the genotypic variation. Two SNPs were colocalized with genes having participation on programmed cell death. Another paper using GWA mapping identified FER QTLs and collocated the FER and FUM QTLs in five lines, and one determined only an anti-FUM effect. The high variation in the FUM data indicates that the additional genetic regulation of the FUM contamination is likely controlled by many other QTLs [
24]. They stated that the chance of this is low, however, as the detected QTLs covered only a small percentage of the variation, and many remained undetected. For this reason, the detected fumonisin differences are considered less reliable than visual resistance data. For most QTLs, they detected a high level of instability across environments, with heritability ranging between 0.36 and 0.56 [
24]. In maize cultivars, phenolic compounds presented high concentrations (822 mg/kg tyrosol equivalents) and had high resistance to fungal penetration and spreading by
F. verticillioides [
16]. Other resistance studies have identified resistance factors to FER such as the protein phosphatase 2A subunits [
152].
Another approach is elicitor research influencing plant defense systems. Small et al. [
153] tested the elicitor effect under known fungicide treatments; however, none of the elicitors and fungicides controlled FER and fumonisin contamination. The authors hope that better spraying technology, dose, and/or timing may lead to better results. On the other hand, this test proved that elicitors do not have general value, and we should identify specific elicitors (when they exist at all) in this context. Another paper went further and conducted a genome-wide association study on 183 inbred lines. Of 14 SNPs connected to FER resistance, 2 were connected to the starburst symptoms that seldom occur on FER-infected grains. Several QTLs were co-localized with known QTLs, and four were linked to SNPs encoding known defense-related proteins [
150]. This is important, as it indicates that the QTLs slowly receive genetic functions. Another paper also considered SNPs and identified 28 QTLs; however, only the
qRcfv2 gene could be validated.
A new QTL was determined by quantitative PCR, but no expression related to resistance was verified [
154]; furthermore, no toxin measurements were made. An interesting development is the identification of the
FvMK1 gene for
F. verticillioides in maize, which is an ortholog of the genes
FMK1 in
F. oxysporum and
GPMK1 in
F. graminearum [
155]. Importantly, the gene regulates multiple signaling pathways, pathogenicity, and FB1 biosynthesis. The question of how it behaves in resistant and susceptible maize genotypes remains open. Another aspect in this line of research [
156] is that natural infection might have breeding aspects that have seldom been applied. A total of 98 inbreds from Lancaster, IDT, SSS, and SSS/IDT groups were compared, and kernel and cob fractions were also compared for toxin contamination. The visible symptoms and fumonisin content were significantly correlated. However, in some inbreds, the cob had higher fumonisin content than the grains. The explanation for this is not the pathotype of the pathogen but possibly the larger water content of the cob, such that the fungus can grow faster and inoculate the germ part of grains sitting on the rachis [
52]. The difference between grain and cob moisture content varied between 11.5 and 28.3%. As fungal spread stops below 23% grain moisture [
79], fungal growth is possible in the cob for 5–10 days (or more), such that the toxin contamination can increase freely. This is also resistance dependent. Maize varieties have been tested in two locations in Italy: Bergamo and Cremona. The FER response after artificial inoculation was rather divergent; however, two varieties (VA117 and VA1213) were identified that had stable resistance in both locations [
31]. Another important aspect of relevant research was the identification of genes that play a role in FER resistance, such as the jasmonate-mediated monocot-specific 9-lipoxygenase
ZmLOX12 gene, which seems to play a key role in resistance to FER [
157]. The gene also acted against FB
1 accumulation. Disruption of the gene, however, allowed for the development of severe FER symptoms and high FB
1 contamination. In one study, breeding for
F. verticillioides resistance was successful, and five experimental hybrids were identified with low (<4 ppm) fumonisin level [
23].
Naturally, genetics-based studies have attempted to identify resistance genes, including gene analogs or putative genes having an influence on resistance expression. Zhang et al. [
155] identified the
FvMK1 mitogen-activated protein kinase as regulating conidium formation, pathogenesis, and FUM production, which seems to have some significance for plant infection.
The progress in basic research is clear, and the presence of genetically determined resistance to FER is proved and accepted. However, the toxin responses seem to be much more divergent than supposed. For this reason, as in F. graminearum, the key task is not determining the correlations and their closeness but the selection and identification of less susceptible and toxin-contaminated inbreds and hybrids. Both in the cited literature and according to our experience, 20–30% of the genotypes from inbred and hybrid testing may belong to this group. The problem of the explained variability is also a key factor. What can we expect from a QTL determining only 4% of the variation? The effect is probably not significant. In the author’s opinion, methodical problems may also be among the causes for these issues. By decreasing the background noise, better data can be obtained. Many candidate genes should be tested further, and the results may be surprising. To have a chance to make stable calculations, a rather broad database is necessary.
5.4. Aspergillus Ear Rot
At present, aflatoxin seems to be the largest problem of the three toxigenic species discussed. This is the least pathogenic, and, so, it is not easy to perform well-characterized experiments. About 50% of 80 isolates were found to be toxin-producing on rice, and only seven isolates were capable of producing toxins when inoculated into maize ears (Tóth 2023, unpublished). Therefore, the selection of a proper isolate requires care. Regarding this species, toxin data show high divergence and variability, and, in this case, it happens rather regularly that significant aflatoxin is produced without visual infection [
51]. It is not an accident that the different methodologies for AER have the largest share, as aflatoxin is the most toxic of the mycotoxins that we normally work with. As resistance is not easy to obtain and aflatoxin often gives variable results, it often occurs that strategies are suggested to solve it without including resistance. Thus, several strategies may be combined to inhibit aflatoxin contamination or destruct it to produce a healthy crop free of aflatoxin, even if increased host resistance seems to be the best solution [
4]. Many articles and reviews analyzing the management of the aflatoxin contamination do not speak about resistance as a possibility to combat the problem [
158], at most mentioning the necessity of the more resistant varieties are necessary, without any suggestion of how to achieve this. However, the publication of resistance studies and results is increasing. Womack et al. (2020) [
159] mapped 241 F
2:3 families for aflatoxin contamination. Xu et al. [
160] mentioned that resistance differences were presented but did not include information about registered hybrids against
A. flavus and aflatoxin contamination. Kumar et al. [
161] did not mention the possibility of utilizing plant resistance. Seed trade firms who suggest aflatoxin control plans for growers typically do not mention the use of more resistant hybrids [
162].
On the other hand, there is nearly 50-year-old research presenting significant research results indicating that resistance and breeding for resistance provide a good perspective for the reduction of aflatoxin contamination. This is the reason why we take this research direction seriously and analyze the problems of how to manage it with a much higher efficacy.
It is seldom that an acceptable correlation exists between Aspergillus symptoms and aflatoxin contamination [
10,
51]. In several cases, visually symptomless genotypes could yield several 100 mg/kg of AFB1, and, in several genotypes, very high overproduction of AFB1 was the case compared to the symptoms. In other cases, AFB1 contamination was much lower than predicted according to the ear infection severity. All of this was observed as a mean reaction of two isolates separately applied in the tests. It also occurred that the correlation between the results of the two isolates was also seldom significant. As a result, in 2021, we used three independent isolates to identify
A. flavus resistance and aflatoxin contamination with higher reliability. The main result was that we identified several hybrids with low variance across isolates and years for both infection severity (resistance) and low aflatoxin contamination. For others, extremely high variances were also computed. This agrees well with the conclusion of Wilcox et al. [
163]. As the aflatoxin production for 1% of isolates was the largest for AER (57-fold), many of them were correlation-breaking; it is no wonder that a generally valid correlation seldom would be the case [
51]. This underlines the necessity to identify hybrids with both low disease severity and low aflatoxin contamination, which was observed in about 20–30% of the tested hybrids; the rest may be suitable for the discovery of pathways for increasing or decreasing resistance-independent aflatoxin synthesis [
51]. Wahl et al. [
164] tested 295 experimental hybrids, in which the mean aflatoxin was 323–370 mg/kg. They identified 13 high-yielding hybrids with a lower aflatoxin level than the resistant control.
The polygenic nature of resistance and the low pathogenicity of the fungus do not help much to resolve the problem [
51]. On the other hand, many papers have reported significant resistance differences between maize genotypes, supporting the reality of breeding for resistance [
8,
159]. Aspergillus ear rot and aflatoxin with fumonisin contamination together can follow damage by insects [
3,
63]; therefore, insecticides or
Bt genes controlling FER and AER can inhibit toxin accumulation. The observation of preharvest aflatoxin contamination by
A. flavus without insect damage indicated the infection ability of the fungus without any insect mediation and raised the need to gain resistance to the fungus and its toxins [
85]. This is the subject of this treaty. In spite of the discovery of the significance of preharvest aflatoxin contamination in maize [
83,
84],
A. flavus was thought to be a storage fungus, following the classic terms [
79]. Therefore, in Europe, the preharvest nature of aflatoxin contamination was only proved recently; for example, in Hungary, this was in 2022 [
51]. Based on this, the preharvest contamination was not controlled and was classified automatically as post-harvest contamination.
Earlier and recent findings cited in this review support the view [
8,
85] that real genetic differences exist, and resistance is believed to be primarily additive. It was stressed 36 years ago that both genetic and non-genetic management should be applied, as no practices had a major and consistent effect. It was added that resistance and aflatoxin regulation might arise from different mechanisms. Furthermore, the resistance of dent inbreds was about eight-fold better than that of sweetcorn inbreds. There are examples of studies on
A. flavus resistance and resistance to aflatoxin contamination [
108]. Abbas et al. [
165] tested maize hybrids for natural aflatoxin and fumonisin contents. AFB1 varied between 27 and 641 μg/kg, while, for FB1, the variation was between 12.1 and 70 mg/kg. As the data came from one year, we can only suppose that genetic differences were behind these variations. As tests were conducted in freshly harvested maize, the mycotoxin contamination was of a preharvest character. It is remarkable that the aflatoxin contamination was, in epidemic years, comparable with Hungarian data at a very low natural visual infection level. Adejuma and Adejoro [
90] posed genetic resistance as a possible solution, only citing previous data. Munkvold [
121] reported on the early achievements in aflatoxin resistance research in the USA. Inbreds and hybrids with good resistance were identified and bred and were also introduced into the market. Active breeding programs have also been reported. Notably, it had been stressed 20 years ago that plants under stress are more susceptible to toxigenic fungi. Darrah and Zuber [
166] have reported strong and significant differences in the aflatoxin response between flint and dent inbreds; the flints were generally better, but the results differed between locations. Henry et al. [
38] chose to take a practical approach, identifying resistant inbreds (KO679Y and CUBA117:S15-101-001-B-B-B-B) through resistance testing, along with similar resistant and susceptible control lines with known response to
A. flavus. The hybrids were promising in terms of both symptom and aflatoxin reduction. The disease pressure was high, so differences in resistance were well-differentiated. Henry et al. [
27] inoculated 20 inbreds with
F. verticillioides and
A. flavus. For FER, the correlation between ear rot severity and fumonisin content was r = 0.74, while that for
A. flavus was r = 0.61 (both significant). Of the 20 hybrids, 3 were resistant to both fungi and their toxins, 3 were highly susceptible to both, and the 14 others varied (the aflatoxin data seemed to be very high, such as 10,592 mg/kg (I checked the original data, this is written, but might be a mistake instead of μg/kg), even though it was ln(y + 1)-transformed data. However, they concentrated on the use of atoxic
A. flavus strains to reduce aflatoxin contamination. The subjects of the resistance tests were mostly inbreds, but hybrids were also tested for practical reasons [
164]. A recent review [
167] identified stress-related genes and antifungal proteins in endosperm, rachis, embryo, and silk tissues before
A. flavus infection and following inoculation. These proteins increased at a much higher speed in the susceptible lines. Plants having a transformed a-amylase inhibitor protein originating from
Lablab purpurea could destroy 56% of aflatoxin contamination in the kernel tissue.
QTL, SNP, and GWAM. Aspergillus resistance in maize is still an unsolved problem. Baisakh et al. [
168] concluded that, while AER-resistance loci could be identified, their phenotypic expression was very variable. In a meta-analysis of 276 from 356 identified QTLs, 58 MQTLs were identified in all 10 chromosomes. A meta-analysis of the differently expressed genes resulted in 591 genes with a putative response to
A. flavus only. Of these, 14 SNPs were identified, and 12 MQTL-linked (meta-QTL) SSR markers identified three markers that could discriminate 14 resistant and 8 susceptible cultivars. However, no aflatoxin mapping was made, such that forecasting of the aflatoxin content could not be concluded. Another paper [
169] reported that most commercial hybrids are susceptible to
A. flavus and aflatoxin contamination. Through a multiple QTL analysis, several dozen QTLs with small effect were identified. An additional Pathway Association Study (PAST) highlighted several genes, including mechanisms that could determine a more or highly specific and well-inheritable trait allowing for more effective genomic selection. Aflatoxin data were not analyzed. It seems that the small-effect QTLs provide low probability to reach an effective genetic tool, even though a convincing resistance difference is demonstrated in Willcox et al. [
163] verifying QTLs against
A. flavus from the resistant maize donor Mp313E under different genetic backgrounds, which were stable under different environmental conditions. Of the 20 QTLs, 5 were identical to those mapped in the Mp313 × B73 mapping population. For this reason, we can determine valuable and stable sources for breeding; on the other hand, the 15 QTLs without stability should not be considered target QTLs. Another paper [
170,
171] conducted
A. flavus inoculation, but only aflatoxin contamination was measured in order to supposedly identify the disease and toxin resistance. From the identified QTLs, only a minority proved to be effective for more than one year, and three of those detected in 1996/1997 were tested further for breeding use. As symptoms were not tested, the relationship between fungal resistance and resistance against toxin accumulation could not be verified. The other important conclusion was that the stability of the QTLs (or stability of the resistance) is a key problem in breeding. Another important result is that, in 1990, the Aspergillus-resistant germplasm Mp313E was released, followed by Mp420, Mp715, Mp717, Mp718, and MP719 [
172,
173]. The only problem here is that all aflatoxin data were transformed using the function ln(y + 1), such that the real aflatoxin data remained hidden, and it is not possible to see the original toxin concentrations. When grain is sold, the key question is whether the total aflatoxin contamination is lower or higher than 20 μg/kg. The ln(y + 1) function is uninteresting. As
A. flavus data come mostly from temperate regions and not from the most-affected tropical region, tropical maize may become an important tool for finding better sources of resistance [
174]. Seven SNPs have been found to be significantly correlated with AER resistance. A pathway analysis revealed 56 associated pathways, including phytoalexins, signaling hormones, and starch biosynthesis growth, as well as metabolic pathways. It seems that GWAS and pathway analysis can identify resistance candidate genes. The chitinase genes of maize might play a similar role [
175], and one such gene is close to the peak of a recently published QTL [
176].
Musungu et al. [
177] detected an aflatoxin regulation cluster based on reactive oxygen species, which influences vesicular aflatoxin transport. As these events occur in the first 1–3 days after inoculation, this might explain early Aspergillus infection and aflatoxin transport and production [
177]. Another innovative technology is gene silencing [
178]; in this case, the
A. flavus alkaline protease was tested, and eight variants were produced and backcrossed to susceptible parents. In the laboratory, an 84% reduction in aflatoxin was observed. The small RNAs presented a 1000-fold higher concentration in transformed plants; therefore, it is probable that the lower
A. flavus severity and aflatoxin contamination is a result of gene silencing, achieved through the use of host-induced gene silencing (HIGS, host-induced gene silencing) technology. Another result in this line is the silencing of the kernel-specific RNA gene cassette targeting
aflC gene [
179]. No aflatoxin was observed in the transformed plant, while, in the control, aflatoxin at the level of thousands of parts per billion was recorded. Antifungal proteins have been identified [
180] at higher levels in resistant plants compared with susceptible ones. A stress-related peroxiredoxin antioxidant (PER1) was cloned, and its expression in the resistant genotype was significantly higher than is a susceptible B73 line.
The question remains the same as Warburton and Williams [
39] have asked previously: “Will this work lead to practical resistance results against
A. flavus and aflatoxin contamination?” The presented data over several decades [
83,
84,
85,
166,
181,
182,
183], along with later published papers (cited by Mesterhazy) [
8] and more recent publications, have presented convincing data for differences in resistance; therefore, our answer is yes, this is possible. Nobody said that this resistance is immunity. It is polygenic and partial. The most complex polygenic trait is yielding ability. In spite of this, nobody states that a breeding for higher a yield would be impossible. Resistance testing methods have been evaluated, presenting convincing, significant, and large resistance differences that are suitable for utilization in breeding. Later developments concentrated on the molecular genetic work, but the underlying methodology remained the same. As the basic data were problematic, all conclusions had this problem. Statistical methods have even been developed to improve the quality of the data, with moderate success. At the same time, it is also true that a large number of QTLs are false positives or false negatives, environmentally dependent, and explain only a minor part of the variability observed. SNP and GWAM data are also problematic in many cases, and, similarly to QTLs, they typically explain only a smaller part of the phenotypic variability and likely have lower reliability. However, this problem has not been widely discussed in the existing literature. Most articles have concentrated on the visual symptoms, only recently coming to the realization that ear rot resistance and aflatoxin contamination are well-correlated in a number of genotypes, while, in others, the case is different. We consider all data to be highly reliable. It is uncertain whether current phenotyping methods are really sufficient to allow for solid genetic conclusions to be drawn from experimental data. Even so, most of the data refer to mainly additive inheritance, while, to a lesser degree, epistatic effects have also been reported. The big picture is possible in this light, but the conclusions for individual cases might be problematic. The existing data do not address the issue of immunity against
A. flavus. However, the breeding of low-risk and stable hybrids is possible with the present knowledge. According to breeders, validated and effective genes or germplasms are required. To make such measurements, we have to prove their stability. For this, we need much more data than we have obtained to date. With increasing experience, the risk can be further decreased. At present, several resistance gene candidates show promise.
5.5. Common or Diverse Resistance Mechanisms against Ear Rot Fungi
The idea of the common resistance was implicitly included in the idea of resistance to natural ear rots. In wheat, such a common resistance has been proved [
86,
87,
88]. For this reason, it is interesting to consider whether such a case is possible in maize. However, it is more important to determine how can we build up a screening system in which all three major pathogens are well-represented. This idea is not new. In our previous review article [
8], several papers were cited which sought relationships between
F. graminearum and
F. culmorum,
F. graminearum and
F. verticillioides, and
F. verticillioides and
A. flavus. In every paper, low-to-moderate correlations were found, but no general consensus was reached. Except us nobody have dealt with the resistance to all three toxigenic species and their toxin relations.
The only positive exception was the correlation series between resistance to
F. graminearum and
F. culmorum, which was consequently close and highly significant [
10,
48,
52,
73,
184], indicating the similar resistance background with respect to these fungi. This is valid also for resistance to DON accumulation.
Bolduan et al. [
17], (as shown in their Figure 3 [
17]), observed a phenotypic correlation between
F. graminearum and
F. verticillioides symptoms (r = 0.63,
p = 0.05). Looking at the plotted data, it is clear that, for up to 70% GER severity, no significant FUM contamination was present; meanwhile, inbreds having near 100% GER varied between 0 and 40% FER. For mycotoxins, the original data were not given (only the ln mg/kg data), and the correlation between DON and fumonisin contamination (ln mg/kg) reached r = 0.59 (
p = 0.05). Of the hybrids, three were of types having very different responses for the toxins; for example, an inbred at DON 6.8 presented no fumonisin contamination. This indicates that a significant part of the inbreds behave similarly with respect to the two toxins, while the rest did not. Therefore, the behavior of DON cannot be forecast with the accuracy required for a breeding program. Other researchers [
185] have identified a 2OGD superfamily gene group, which was detected as having similar functions against the two ear rot pathogens. This explains the similarities in resistance to the two fungal species in several genotypes, as also detected previously [
50]; however, this result was not consistent for the toxin response or in another hybrid group with an entirely different genetic background. As the paper did not present aflatoxin data, the food safety aspects remain unclear. Cary et al. [
4] suggested combining the resistance of African and USA lines with respect to
A. flavus,
F. verticillioides, and others. The beta-glucuronidase (
GUS) reporter gene has an influence on
A. flavus resistance in maize. The GER and FER resistance additive and dominant effects were predominant in most environments in response to these diseases and their toxins. However, the stability of their performance was low. Different genetic–environmental interactions were observed, indicating some degree of common resistance. Chiuraise et al. [
23] have found, in crosses of fumonisin- and aflatoxin-resistant lines, that several lines had good resistance against both toxins. A dissimilarity between Hungarian and Kenyan data was the higher infection severity in response to both pathogens: fumonisin maxima were higher in Kenya, while aflatoxin maxima were much higher in Hungary [
10,
52]. In a larger number of hybrids, several were identified to have common resistance, but the majority of the hybrids responded differently [
10,
52]. A meta-analysis of QTLs and candidate genes was conducted in 224 QTL from 15 papers in a dense genome-wide SNP analysis and identified a total of 40 MQTLs (metaQTLs) [
186], of which 29 were associated with 2–5 FER- and/or GER-related traits, 28 were common for FER and GER, and 19 were common for silk and kernel resistance. Promising candidate genes were also identified. As mycotoxin responses were not assessed, the value of the work in this respect remains unclear. The main message of this work is that QTLs with distinct functions exist, making the problem even more complicated but potentially explaining the varying responses to different
Fusarium species and inoculation methods.
This is similar to the results that we obtained in wheat [
135,
136] and later in maize [
51]. Guche et al. [
25] identified the
ZmLOXs genes participating in oxylipin biosynthesis as taking part in the resistance to
F. verticillioides and
A. flavus.
ZmLOX4 seems to be the most effective, which was identified in the inbreds Tzi18, Mo17, and W22. Oxylipin synthesis is connected to linoleic peroxidation by 9-LOXs and their accumulation. Triglyceride peroxidation was only observed in genotypes with resistance to
F. verticillioides. The mechanisms were remarkably similar against the two fungal species, and they also were found to influence the mycotoxin content. Stagnati et al. [
49] tested a hybrid collection against
F. verticillioides and
A. flavus. Regarding the symptoms, seven presented resistances to both pathogens, while, for toxins, only three were superior. These findings support the Hungarian results. Bolduan [
17] obtained a moderate (r
p = 0.59,
p = 0.05) (r
p = correlation, phenotypic) to moderately high (r
g = 0.77) correlation (r
g = correlation, genetic) between the two toxin reactions of the inbreds. Another paper [
187] reported on crosses using the inbreds Co430 (
F. graminearum) and Mp420 (
A. flavus) with a susceptible inbred and backcrossed. Their results suggested that the best lines selected from these crosses also had resistance to
F. verticillioides. The authors hoped that this result would indicate general significance. Abbas et al. [
37] did not find any correlation between aflatoxin and fumonisin contamination; however, with other plant materials, significant positive or medium correlations were obtained [
165]. This seemingly contradictory situation is the same in Hungary, where the results changed from test to test.
On the other hand, the selection of S5 inbreds resulted in several cases besides increased resistance to
F. verticillioides, such as resistance to
F. graminearum [
188]. Guo et al. [
26] screened 87 inbred lines mostly from China and Mexico and tested them for preharvest aflatoxin and fumonisin contamination. The aflatoxin level ranged from 50 to 1524 mg/kg, while the fumonisin level varied between 0.6 and 124 mg/kg (natural infection). Five lines showed very low aflatoxin content (artificial), four presented very low fumonisin content (ELISA test was used), and only one (TUN61) showed good resistance to both. The data support the conclusions we obtained from maize hybrids [
10].
We reported a common resistance to
F. graminearum and
F. culmorum [
189], which later tests further confirmed [
50,
52].
The resistances to
F. graminearum,
F. verticillioides, and
A. flavus have been assessed over the last 10 years. We decided to test hybrids instead of inbred lines, as we aimed to help growers by determining which hybrid has high, low, or medium risk with respect to disease and toxins. The results clearly indicated that a common resistance does not exist to the three main pathogens in maize; therefore, resistance should be evaluated separately. In several tests, significant correlations were found between resistance responses while, in others, no correlation existed. The correlations between toxin contamination were sometimes exceptionally high, and, in other tests, no significant results were obtained. Until 2020, only two isolates had been tested together for a toxigenic species; since 2021, three isolates have been tested [
10,
50,
52]. Through these tests, the stability of the responses could be determined. Despite the fact that the hybrids showed very variable resistance to disease and toxin accumulation, hybrids with low and stable resistance, even to all toxigenic species and their toxins, could be identified.
The differences in resistance tend to be highly significant, with the difference between minimum and maximum values often being higher than ten-fold. This makes it possible to identify more resistant hybrids, which should be preferred during variety registration, by screening commercial varieties. On the other hand, the exclusion of the highly susceptible hybrids will result in a sharp improvement of food and feed safety, and, through increasing breeding activities, even better control may be possible. This contributes to food and feed security, and significantly more grain can be used for food and feed and not lost.






{kind=link}
{kind=link}