

Circulating and Cardiac Tissue miRNAs in Children with Dilated Cardiomyopathy
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Samples
2.2. miR Array and RNA-Seq
2.3. Array Analysis
2.4. Pathway Analysis
2.5. Statistical Methods
3. Results
3.1. Study Participant Characteristics
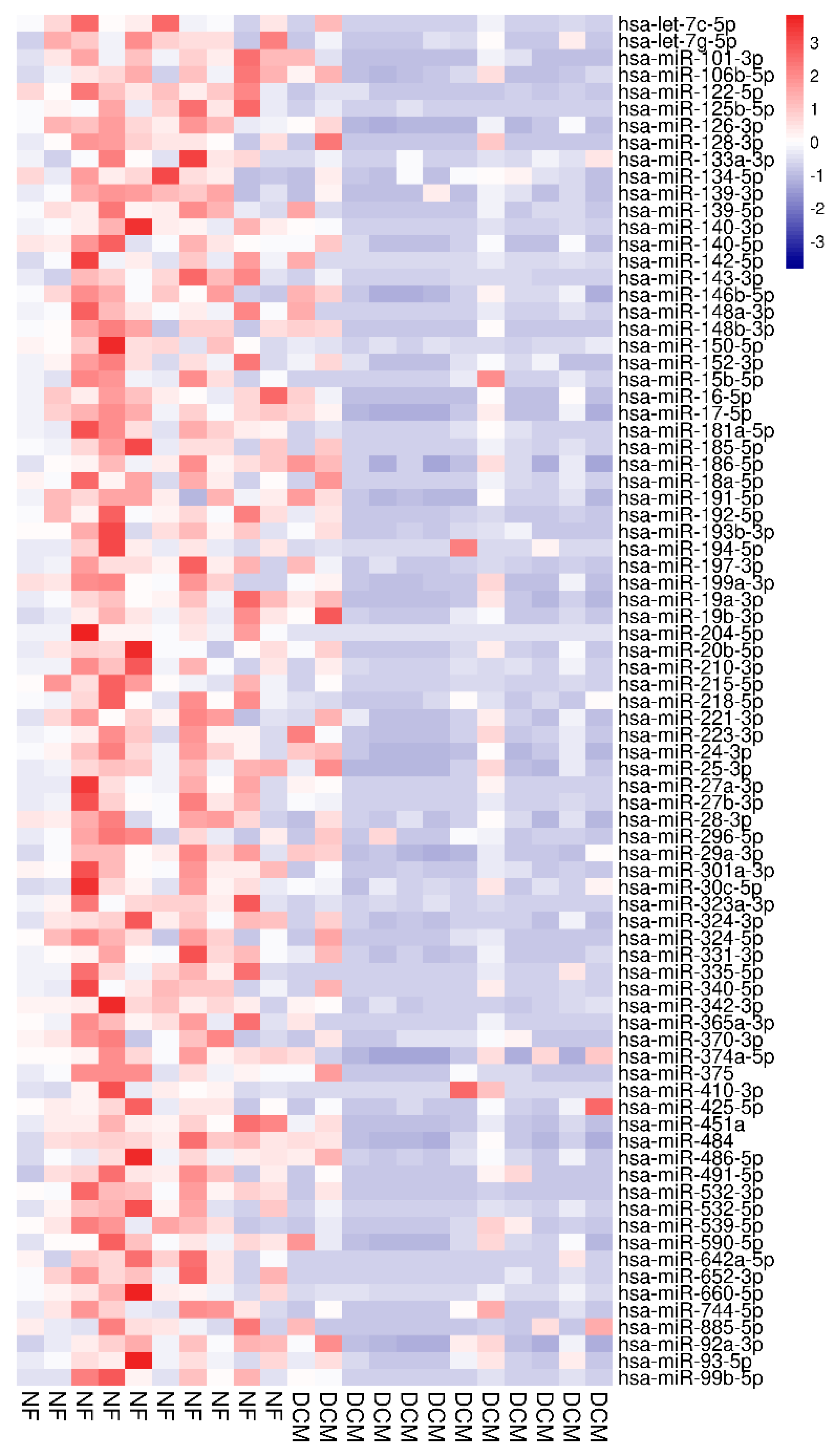
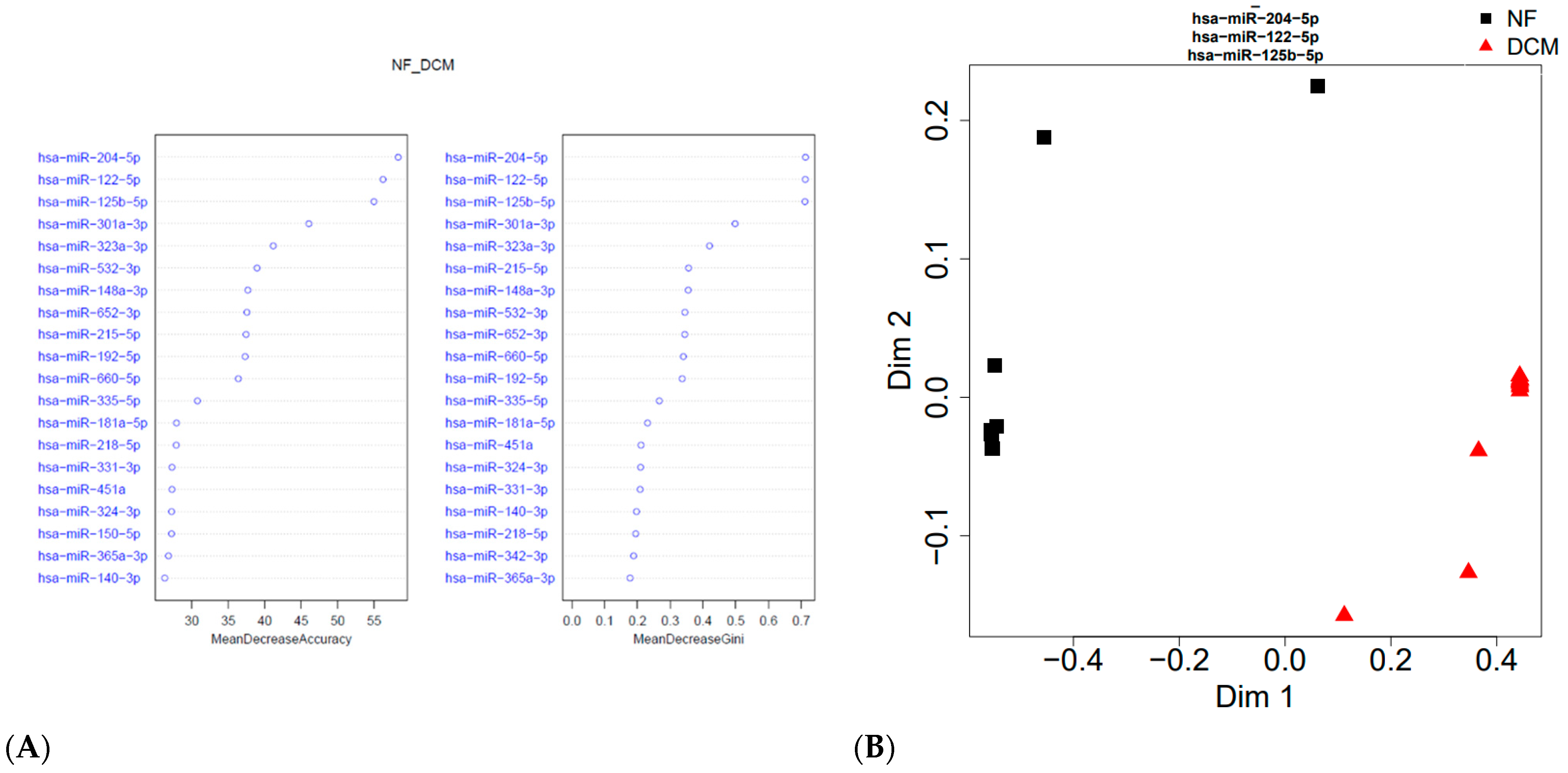
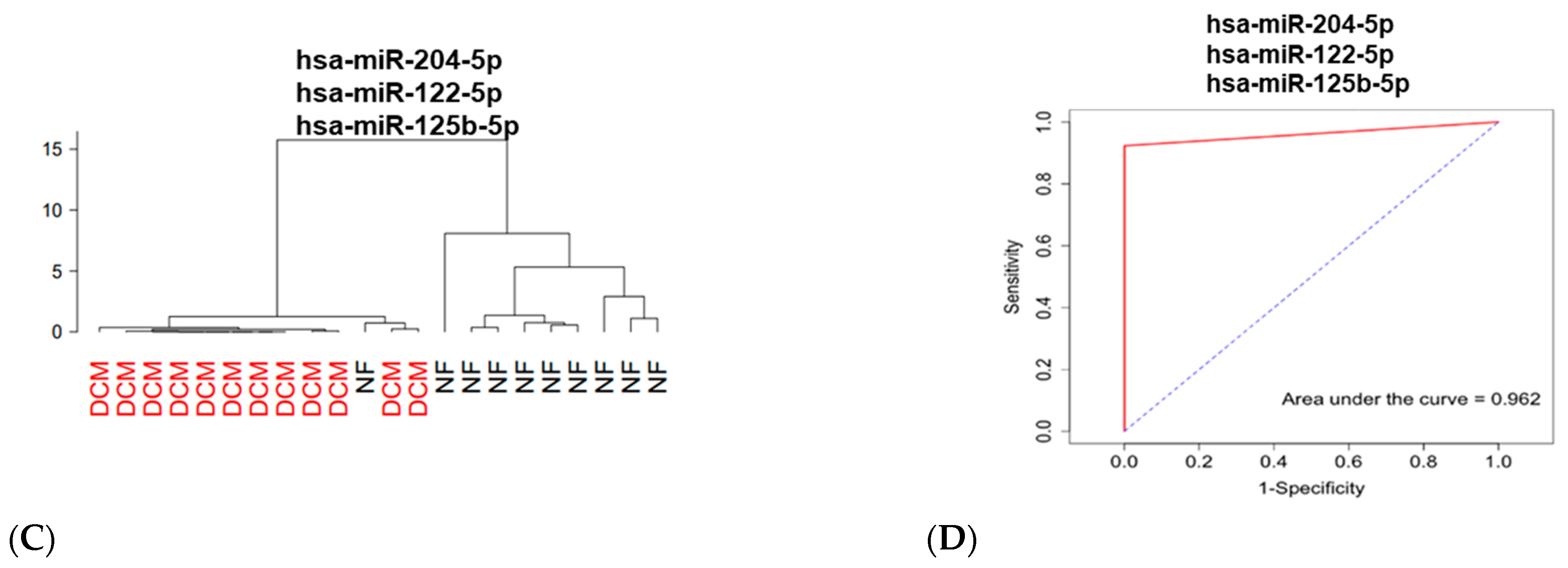
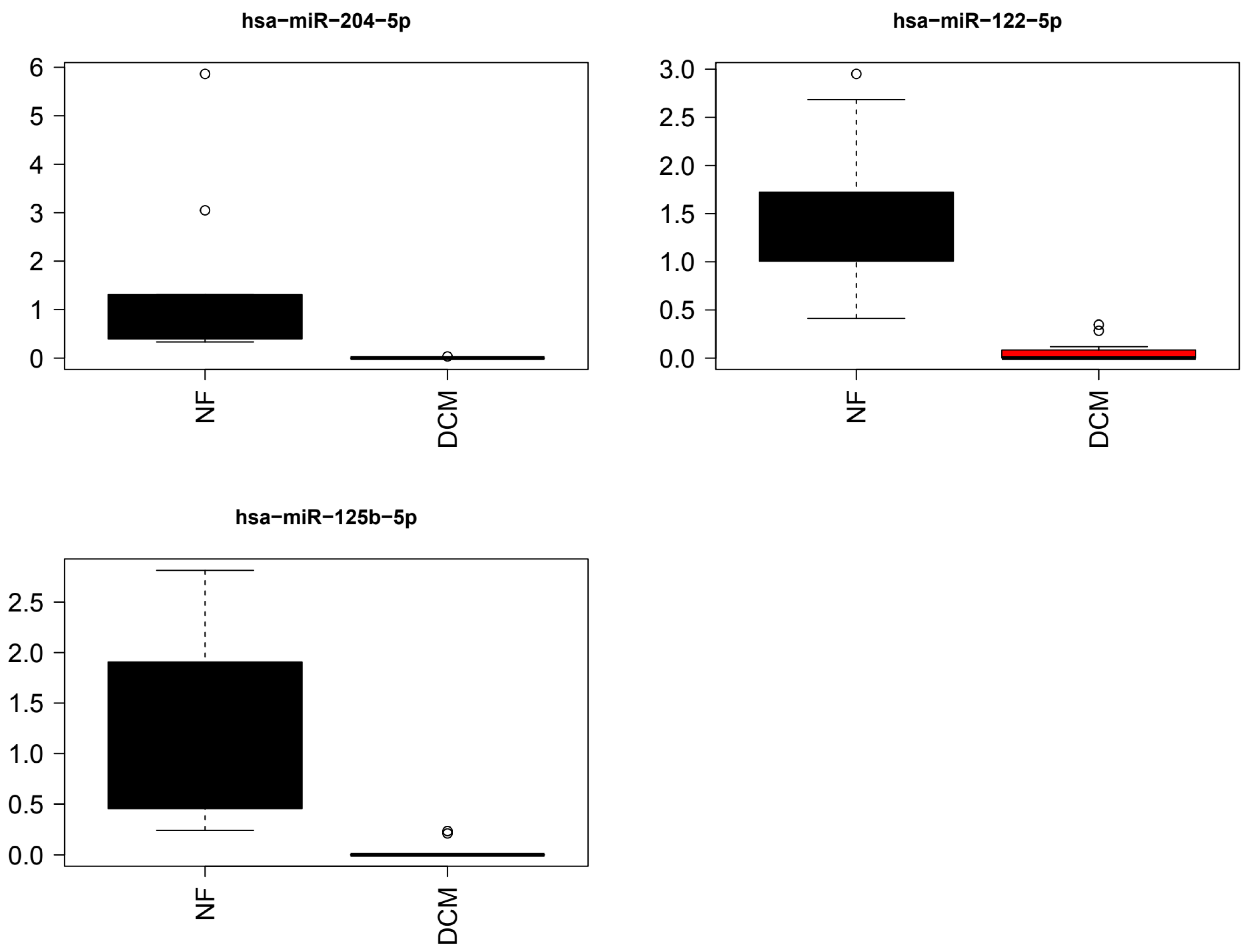
3.2. Identification of Differentially Expressed Circulating miRs
3.3. Association between Circulating and Cardiac Tissue miRs
3.4. Pathway Analysis
4. Discussion
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kirk, R.; Edwards, L.B.; Kucheryavaya, A.Y.; Aurora, P.; Christie, J.D.; Dobbels, F.; Rahmel, A.O.; Stehlik, J.; Hertz, M.I. The Registry of the International Society for Heart and Lung Transplantation: Thirteenth official pediatric heart transplantation report—2010. J. Heart Lung Transpl. 2010, 29, 1119–1128. [Google Scholar] [CrossRef]
- Jayaprasad, N. Heart Failure in Children. Heart Views 2016, 17, 92–99. [Google Scholar] [CrossRef]
- Shaddy, R.E.; Boucek, M.M.; Hsu, D.T.; Boucek, R.J.; Canter, C.E.; Mahony, L.; Ross, R.D.; Pahl, E.; Blume, E.D.; Dodd, D.A.; et al. Carvedilol for children and adolescents with heart failure: A randomized controlled trial. JAMA 2007, 298, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Canter, C.E.; Shi, L.; Colan, S.D.; Dodd, D.A.; Everitt, M.D.; Hsu, D.T.; Jefferies, J.L.; Kantor, P.F.; Pahl, E.; et al. Survival Without Cardiac Transplantation Among Children With Dilated Cardiomyopathy. J. Am. Coll. Cardiol. 2017, 70, 2663–2673. [Google Scholar] [CrossRef] [PubMed]
- Towbin, J.A.; Lowe, A.M.; Colan, S.D.; Sleeper, L.A.; Orav, E.J.; Clunie, S.; Messere, J.; Cox, G.F.; Lurie, P.R.; Hsu, D.; et al. Incidence, causes, and outcomes of dilated cardiomyopathy in children. JAMA 2006, 296, 1867–1876. [Google Scholar] [CrossRef]
- Nakano, S.J.; Miyamoto, S.D.; Movsesian, M.; Nelson, P.; Stauffer, B.L.; Sucharov, C.C. Age-related differences in phosphodiesterase activity and effects of chronic phosphodiesterase inhibition in idiopathic dilated cardiomyopathy. Circ. Heart Fail. 2015, 8, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Woulfe, K.C.; Siomos, A.K.; Nguyen, H.; SooHoo, M.; Galambos, C.; Stauffer, B.L.; Sucharov, C.; Miyamoto, S. Fibrosis and Fibrotic Gene Expression in Pediatric and Adult Patients With Idiopathic Dilated Cardiomyopathy. J. Card. Fail. 2017, 23, 314–324. [Google Scholar] [CrossRef]
- Tatman, P.D.; Woulfe, K.C.; Karimpour-Fard, A.; Jeffrey, D.A.; Jaggers, J.; Cleveland, J.C.; Nunley, K.; Taylor, M.R.; Miyamoto, S.D.; Stauffer, B.L.; et al. Pediatric dilated cardiomyopathy hearts display a unique gene expression profile. JCI Insight 2017, 2, e94249. [Google Scholar] [CrossRef]
- Miyamoto, S.D.; Stauffer, B.L.; Nakano, S.; Sobus, R.; Nunley, K.; Nelson, P.; Sucharov, C.C. Beta-adrenergic adaptation in paediatric idiopathic dilated cardiomyopathy. Eur. Heart J. 2014, 35, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, B.L.; Russell, G.; Nunley, K.; Miyamoto, S.D.; Sucharov, C.C. miRNA expression in pediatric failing human heart. J Mol. Cell Cardiol. 2013, 57, 43–46. [Google Scholar] [CrossRef]
- Hailu, F.T.; Karimpour-Fard, A.; Toni, L.S.; Bristow, M.R.; Miyamoto, S.D.; Stauffer, B.L.; Sucharov, C.C. Integrated analysis of miRNA-mRNA interaction in pediatric dilated cardiomyopathy. Pediatr Res. 2021, 92, 98–108. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Zhou, S.S.; Jin, J.P.; Wang, J.Q.; Zhang, Z.G.; Freedman, J.H.; Zheng, Y.; Cai, L. miRNAS in cardiovascular diseases: Potential biomarkers, therapeutic targets and challenges. Acta Pharmacol. Sin. 2018, 39, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Vegter, E.L.; van der Meer, P.; de Windt, L.J.; Pinto, Y.M.; Voors, A.A. MicroRNAs in heart failure: From biomarker to target for therapy. Eur. J. Heart Fail. 2016, 18, 457–468. [Google Scholar] [CrossRef]
- Miyamoto, S.D.; Karimpour-Fard, A.; Peterson, V.; Auerbach, S.R.; Stenmark, K.R.; Stauffer, B.L.; Sucharov, C.C. Circulating microRNA as a biomarker for recovery in pediatric dilated cardiomyopathy. J. Heart Lung Transpl. 2015, 34, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Mariner, P.D.; Korst, A.; Karimpour-Fard, A.; Stauffer, B.L.; Miyamoto, S.D.; Sucharov, C.C. Improved Detection of Circulating miRNAs in Serum and Plasma Following Rapid Heat/Freeze Cycling. Microrna 2018, 7, 138–147. [Google Scholar] [CrossRef]
- Jone, P.N.; Korst, A.; Karimpour-Fard, A.; Thomas, T.; Dominguez, S.R.; Heizer, H.; Anderson, M.S.; Glode, M.P.; Sucharov, C.C.; Miyamoto, S.D. Circulating microRNAs differentiate Kawasaki Disease from infectious febrile illnesses in childhood. J. Mol. Cell. Cardiol. 2020, 146, 12–18. [Google Scholar] [CrossRef]
- Jeffrey, D.A.; Pires Da Silva, J.; Garcia, A.M.; Jiang, X.; Karimpour-Fard, A.; Toni, L.S.; Lanzicher, T.; Pena, B.; Miyano, C.A.; Nunley, K.; et al. Serum circulating proteins from pediatric patients with dilated cardiomyopathy cause pathologic remodeling and cardiomyocyte stiffness. JCI Insight 2021, 6, e148637. [Google Scholar] [CrossRef]
- Cakmak, H.A.; Coskunpinar, E.; Ikitimur, B.; Barman, H.A.; Karadag, B.; Tiryakioglu, N.O.; Kahraman, K.; Vural, V.A. The prognostic value of circulating microRNAs in heart failure: Preliminary results from a genome-wide expression study. J. Cardiovasc. Med. 2015, 16, 431–437. [Google Scholar] [CrossRef]
- de Gonzalo-Calvo, D.; Davalos, A.; Fernandez-Sanjurjo, M.; Amado-Rodriguez, L.; Diaz-Coto, S.; Tomas-Zapico, C.; Montero, A.; Garcia-Gonzalez, A.; Llorente-Cortes, V.; Heras, M.E.; et al. Circulating microRNAs as emerging cardiac biomarkers responsive to acute exercise. Int. J. Cardiol. 2018, 264, 130–136. [Google Scholar] [CrossRef]
- Bar, C.; Thum, T.; de Gonzalo-Calvo, D. Circulating miRNAs as mediators in cell-to-cell communication. Epigenomics 2019, 11, 111–113. [Google Scholar] [CrossRef]
- Chu, M.; Wu, R.; Qin, S.; Hua, W.; Shan, Z.; Rong, X.; Zeng, J.; Hong, L.; Sun, Y.; Liu, Y.; et al. Bone Marrow-Derived MicroRNA-223 Works as an Endocrine Genetic Signal in Vascular Endothelial Cells and Participates in Vascular Injury From Kawasaki Disease. J. Am. Heart Assoc. 2017, 6, e004878. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Qin, S.; Li, W.; Wu, W.; Yang, J.; Chu, M.; Li, X.; Huo, Y.; Schaer, G.L.; Wang, S.; et al. An Endocrine Genetic Signal Between Blood Cells and Vascular Smooth Muscle Cells: Role of MicroRNA-223 in Smooth Muscle Function and Atherogenesis. J. Am. Coll. Cardiol. 2015, 65, 2526–2537. [Google Scholar] [CrossRef]
- Jansen, F.; Stumpf, T.; Proebsting, S.; Franklin, B.S.; Wenzel, D.; Pfeifer, P.; Flender, A.; Schmitz, T.; Yang, X.; Fleischmann, B.K.; et al. Intercellular transfer of miR-126-3p by endothelial microparticles reduces vascular smooth muscle cell proliferation and limits neointima formation by inhibiting LRP6. J. Mol. Cell. Cardiol. 2017, 104, 43–52. [Google Scholar] [CrossRef]
- Jiang, X.; Sucharov, J.; Stauffer, B.L.; Miyamoto, S.D.; Sucharov, C.C. Exosomes from pediatric dilated cardiomyopathy patients modulate a pathological response in cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H818–H826. [Google Scholar] [CrossRef]
- Gaddam, R.R.; Kim, Y.R.; Jacobs, J.S.; Yoon, J.Y.; Li, Q.; Cai, A.; Shankaiahgari, H.; London, B.; Irani, K.; Vikram, A. The microRNA-204-5p inhibits APJ signalling and confers resistance to cardiac hypertrophy and dysfunction. Clin. Transl. Med. 2022, 12, e693. [Google Scholar] [CrossRef]
- Wang, R.; Ding, Y.D.; Gao, W.; Pei, Y.Q.; Yang, J.X.; Zhao, Y.X.; Liu, X.L.; Shen, H.; Zhang, S.; Yu, L.; et al. Serum microRNA-204 levels are associated with long-term cardiovascular disease risk based on the Framingham risk score in patients with type 2 diabetes: Results from an observational study. J. Geriatr. Cardiol. 2020, 17, 330–337. [Google Scholar] [PubMed]
- Voellenkle, C.; van Rooij, J.; Cappuzzello, C.; Greco, S.; Arcelli, D.; Di Vito, L.; Melillo, G.; Rigolini, R.; Costa, E.; Crea, F.; et al. MicroRNA signatures in peripheral blood mononuclear cells of chronic heart failure patients. Physiol. Genom. 2010, 42, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Vizi, D.; Khammy, O.; Mariani, J.A.; Kaye, D.M. The transcardiac gradient of cardio-microRNAs in the failing heart. Eur. J. Heart Fail. 2016, 18, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Chen, M.; Li, L.; He, M.; Hu, D.; Zhang, X.; Li, J.; Tanguay, R.M.; Feng, J.; Cheng, L.; et al. Circulating MicroRNAs and the occurrence of acute myocardial infarction in Chinese populations. Circ. Cardiovasc. Genet. 2014, 7, 189–198. [Google Scholar] [CrossRef]
- Wang, X.; Ha, T.; Zou, J.; Ren, D.; Liu, L.; Zhang, X.; Kalbfleisch, J.; Gao, X.; Williams, D.; Li, C. MicroRNA-125b protects against myocardial ischaemia/reperfusion injury via targeting p53-mediated apoptotic signalling and TRAF6. Cardiovasc. Res. 2014, 102, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.H.; Zhang, K. MiR-122 regulates cell apoptosis and ROS by targeting DJ-1 in renal ischemic reperfusion injury rat models. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8830–8838. [Google Scholar]
- Wang, Y.; Liang, H.; Jin, F.; Yan, X.; Xu, G.; Hu, H.; Liang, G.; Zhan, S.; Hu, X.; Zhao, Q.; et al. Injured liver-released miRNA-122 elicits acute pulmonary inflammation via activating alveolar macrophage TLR7 signaling pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 6162–6171. [Google Scholar] [CrossRef]
- Song, J.J.; Yang, M.; Liu, Y.; Song, J.W.; Wang, J.; Chi, H.J.; Liu, X.Y.; Zuo, K.; Yang, X.C.; Zhong, J.C. MicroRNA-122 aggravates angiotensin II-mediated apoptosis and autophagy imbalance in rat aortic adventitial fibroblasts via the modulation of SIRT6-elabela-ACE2 signaling. Eur. J. Pharmacol. 2020, 883, 173374. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, K.C.; Sparagna, G.C.; Sucharov, C.C.; Miyamoto, S.D.; Grudis, J.E.; Sobus, R.D.; Hijmans, J.; Stauffer, B.L. Dysregulation of cardiolipin biosynthesis in pediatric heart failure. J. Mol. Cell. Cardiol. 2014, 74, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Tian, R. Mitochondrial dysfunction in pathophysiology of heart failure. J. Clin. Investig. 2018, 128, 3716–3726. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Fold Change | qValue_wilcox |
|---|---|---|
| hsa-let-7c-5p | −0.89117 | 0.006447 |
| hsa-let-7g-5p | −0.74433 | 0.011349 |
| hsa-miR-101-3p | −0.84796 | 0.006447 |
| hsa-miR-106b-5p | −0.59646 | 0.015417 |
| hsa-miR-122-5p | −1.46702 | 0.002372 |
| hsa-miR-125b-5p | −1.1267 | 0.002331 |
| hsa-miR-126-3p | −0.66084 | 0.004093 |
| hsa-miR-128-3p | −0.59713 | 0.020073 |
| hsa-miR-133a-3p | −1.13195 | 0.048692 |
| hsa-miR-134-5p | −0.63877 | 0.04314 |
| hsa-miR-139-3p | −0.77407 | 0.011413 |
| hsa-miR-139-5p | −0.70595 | 0.006682 |
| hsa-miR-140-3p | −1.09951 | 0.002472 |
| hsa-miR-140-5p | −0.77305 | 0.008129 |
| hsa-miR-142-5p | −0.80908 | 0.00872 |
| hsa-miR-143-3p | −1.16193 | 0.00387 |
| hsa-miR-146b-5p | −0.38407 | 0.048692 |
| hsa-miR-148a-3p | −1.0221 | 0.002982 |
| hsa-miR-148b-3p | −0.7088 | 0.028505 |
| hsa-miR-150-5p | −0.72873 | 0.002472 |
| hsa-miR-152-3p | −0.91037 | 0.021286 |
| hsa-miR-15b-5p | −0.74914 | 0.009937 |
| hsa-miR-16-5p | −0.67087 | 0.004093 |
| hsa-miR-17-5p | −0.63261 | 0.005181 |
| hsa-miR-181a-5p | −0.96636 | 0.002372 |
| hsa-miR-185-5p | −0.9488 | 0.009937 |
| hsa-miR-186-5p | −0.40663 | 0.040325 |
| hsa-miR-18a-5p | −0.66868 | 0.012857 |
| hsa-miR-191-5p | −0.4202 | 0.046648 |
| hsa-miR-192-5p | −0.84033 | 0.002472 |
| hsa-miR-193b-3p | −0.87364 | 0.006447 |
| hsa-miR-194-5p | −0.57466 | 0.012857 |
| hsa-miR-197-3p | −0.79624 | 0.006447 |
| hsa-miR-199a-3p | −0.75922 | 0.008129 |
| hsa-miR-19a-3p | −0.6605 | 0.016026 |
| hsa-miR-19b-3p | −0.47258 | 0.018154 |
| hsa-miR-204-5p | −1.42388 | 0.002331 |
| hsa-miR-20b-5p | −0.70575 | 0.034222 |
| hsa-miR-210-3p | −1.05106 | 0.006447 |
| hsa-miR-215-5p | −1.11726 | 0.002372 |
| hsa-miR-218-5p | −1.05591 | 0.005706 |
| hsa-miR-221-3p | −0.73779 | 0.029695 |
| hsa-miR-223-3p | −0.56938 | 0.026463 |
| hsa-miR-24-3p | −0.64909 | 0.012857 |
| hsa-miR-25-3p | −0.62173 | 0.011006 |
| hsa-miR-27a-3p | −1.06271 | 0.012857 |
| hsa-miR-27b-3p | −1.28136 | 0.003618 |
| hsa-miR-28-3p | −0.66936 | 0.009539 |
| hsa-miR-296-5p | −0.73451 | 0.040325 |
| hsa-miR-29a-3p | −0.53525 | 0.011349 |
| hsa-miR-301a-3p | −0.9541 | 0.002372 |
| hsa-miR-30c-5p | −0.77104 | 0.029695 |
| hsa-miR-323a-3p | −1.05664 | 0.002372 |
| hsa-miR-324-3p | −0.86761 | 0.002982 |
| hsa-miR-324-5p | −0.69613 | 0.014925 |
| hsa-miR-331-3p | −0.67157 | 0.00387 |
| hsa-miR-335-5p | −1.18965 | 0.002372 |
| hsa-miR-340-5p | −0.58158 | 0.015343 |
| hsa-miR-342-3p | −0.5613 | 0.00369 |
| hsa-miR-365a-3p | −1.25129 | 0.002787 |
| hsa-miR-370-3p | −0.62826 | 0.044334 |
| hsa-miR-374a-5p | −0.54854 | 0.038544 |
| hsa-miR-375 | −0.8206 | 0.008122 |
| hsa-miR-410-3p | −0.48513 | 0.038751 |
| hsa-miR-425-5p | −0.59274 | 0.016943 |
| hsa-miR-451a | −0.80526 | 0.002372 |
| hsa-miR-484 | −0.56538 | 0.002372 |
| hsa-miR-486-5p | −0.64054 | 0.038544 |
| hsa-miR-491-5p | −0.98398 | 0.006447 |
| hsa-miR-532-3p | −0.83425 | 0.002372 |
| hsa-miR-532-5p | −1.29285 | 0.006447 |
| hsa-miR-539-5p | −0.73027 | 0.012857 |
| hsa-miR-590-5p | −0.6414 | 0.018154 |
| hsa-miR-642a-5p | −0.62641 | 0.005706 |
| hsa-miR-652-3p | −0.91256 | 0.002372 |
| hsa-miR-660-5p | −1.06831 | 0.00387 |
| hsa-miR-744-5p | −0.71785 | 0.009539 |
| hsa-miR-885-5p | −0.90867 | 0.029695 |
| hsa-miR-92a-3p | −0.49284 | 0.029695 |
| hsa-miR-93-5p | −0.71524 | 0.028889 |
| hsa-miR-99b-5p | −1.44717 | 0.008129 |
| miRNA | Increased in Sera | Decreased in Sera | Increased in Cardiac Tissue | Decreased in Cardiac Tissue |
|---|---|---|---|---|
| hsa-miR-204-5p | x | x | ||
| hsa-miR-125b-5p | x | x | ||
| hsa-miR-301a-3p | x | x | ||
| hsa-miR-133a-3p | x | x | ||
| hsa-miR-150-5p | x | x | ||
| hsa-miR-486-5p | x | x | ||
| hsa-miR-17-5p | x | x | ||
| hsa-miR-92a-3p | x | x | ||
| hsa-miR-20a-5p | x | |||
| hsa-miR-212-3p | x | |||
| hsa-miR-132-3p | x | |||
| hsa-miR-193a-5p | x | |||
| hsa-miR-345-5p | x | |||
| hsa-let-7d-5p | x | |||
| hsa-miR-574-3p | x | |||
| hsa-miR-376c-3p | x | |||
| hsa-miR-195-5p | x | |||
| hsa-miR-130b-3p | x | |||
| hsa-miR-122-5p | x | |||
| hsa-miR-99b-5p | x | |||
| hsa-miR-532-5p | x | |||
| hsa-miR-27b-3p | x | |||
| hsa-miR-365a-3p | x | |||
| hsa-miR-335-5p | x | |||
| hsa-miR-143-3p | x | |||
| hsa-miR-215-5p | x | |||
| hsa-miR-140-3p | x | |||
| hsa-miR-660-5p | x | |||
| hsa-miR-27a-3p | x | |||
| hsa-miR-323a-3p | x | |||
| hsa-miR-218-5p | x | |||
| hsa-miR-210-3p | x | |||
| hsa-miR-148a-3p | x | |||
| hsa-miR-491-5p | x | |||
| hsa-miR-181a-5p | x | |||
| hsa-miR-185-5p | x | |||
| hsa-miR-652-3p | x | |||
| hsa-miR-152-3p | x | |||
| hsa-miR-885-5p | x | |||
| hsa-let-7c-5p | x | |||
| hsa-miR-193b-3p | x | |||
| hsa-miR-324-3p | x | |||
| hsa-miR-101-3p | x | |||
| hsa-miR-192-5p | x | |||
| hsa-miR-532-3p | x | |||
| hsa-miR-375 | x | |||
| hsa-miR-142-5p | x | |||
| hsa-miR-451a | x | |||
| hsa-miR-197-3p | x | |||
| hsa-miR-139-3p | x | |||
| hsa-miR-140-5p | x | |||
| hsa-miR-30c-5p | x | |||
| hsa-miR-199a-3p | x | |||
| hsa-miR-15b-5p | x | |||
| hsa-let-7g-5p | x | |||
| hsa-miR-221-3p | x | |||
| hsa-miR-296-5p | x | |||
| hsa-miR-539-5p | x | |||
| hsa-miR-744-5p | x | |||
| hsa-miR-93-5p | x | |||
| hsa-miR-148b-3p | x | |||
| hsa-miR-139-5p | x | |||
| hsa-miR-20b-5p | x | |||
| hsa-miR-324-5p | x | |||
| hsa-miR-331-3p | x | |||
| hsa-miR-16-5p | x | |||
| hsa-miR-28-3p | x | |||
| hsa-miR-18a-5p | x | |||
| hsa-miR-126-3p | x | |||
| hsa-miR-19a-3p | x | |||
| hsa-miR-24-3p | x | |||
| hsa-miR-590-5p | x | |||
| hsa-miR-134-5p | x | |||
| hsa-miR-370-3p | x | |||
| hsa-miR-642a-5p | x | |||
| hsa-miR-25-3p | x | |||
| hsa-miR-128-3p | x | |||
| hsa-miR-106b-5p | x | |||
| hsa-miR-425-5p | x | |||
| hsa-miR-340-5p | x | |||
| hsa-miR-194-5p | x | |||
| hsa-miR-223-3p | x | |||
| hsa-miR-484 | x | |||
| hsa-miR-342-3p | x | |||
| hsa-miR-374a-5p | x | |||
| hsa-miR-29a-3p | x | |||
| hsa-miR-410-3p | x | |||
| hsa-miR-19b-3p | x | |||
| hsa-miR-191-5p | x | |||
| hsa-miR-186-5p | x | |||
| hsa-miR-146b-5p | x |
| miRNA | Coefficient | p-Value | q Value |
|---|---|---|---|
| hsa-miR-125b-5p | 32.33467 | 0.006746 | 0.007709 |
| hsa-133a-3p | −29.7126 | 0.004939 | 0.006585 |
| hsa-miR-150-5p | −31.855 | 0.000407 | 0.001084 |
| hsa-miR-17-5p | −36.6685 | 0.000131 | 0.000526 |
| hsa-miR-204-5p | 10.94161 | 0.002227 | 0.003563 |
| hsa-miR-301a-3p | 35.57683 | 0.000112 | 0.000526 |
| hsa-miR-486-5p | −30.6944 | 0.000592 | 0.001184 |
| hsa-miR-92a-3p | −31.5682 | 0.012177 | 0.012177 |
| Pathway | p-Value |
|---|---|
| Adipocytokine signaling pathway | 1.40 × 10−12 |
| Mitophagy—animal | 2.86 × 10−12 |
| VEGF signaling pathway | 1.38 × 10−11 |
| Fatty acid biosynthesis | 6.64 × 10−11 |
| Toll-like receptor signaling pathway | 6.73 × 10−11 |
| IL-17 signaling pathway | 7.41 × 10−11 |
| Pyruvate metabolism | 4.16 × 10−10 |
| Glucagon signaling pathway | 1.17 × 10−9 |
| Carbohydrate digestion and absorption | 1.99 × 10−9 |
| Chemokine signaling pathway | 4.82 × 10−9 |
| Hedgehog signaling pathway | 5.05 × 10−9 |
| AMPK signaling pathway | 1.44 × 10−8 |
| Signaling pathways regulating pluripotency of stem cells | 1.49 × 10−8 |
| Wnt signaling pathway | 2.37 × 10−8 |
| Autophagy—other | 4.16 × 10−8 |
| Hippo signaling pathway | 5.00 × 10−8 |
| Fatty acid degradation | 9.44 × 10−8 |
| TGF-beta signaling pathway | 1.00 × 10−7 |
| Hypertrophic cardiomyopathy HCM | 1.01 × 10−7 |
| Glycolysis Gluconeogenesis | 1.21 × 10−7 |
| PPAR signaling pathway | 1.41 × 10−7 |
| Oxidative phosphorylation | 2.45 × 10−7 |
| TNF signaling pathway | 3.92 × 10−7 |
| Arrhythmogenic right ventricular cardiomyopathy ARVC | 8.56 × 10−7 |
| p53 signaling pathway | 1.02 × 10−6 |
| Ubiquitin mediated proteolysis | 1.47 × 10−6 |
| Cytokine–cytokine receptor interaction | 2.85 × 10−6 |
| Dilated cardiomyopathy DCM | 3.15 × 10−6 |
| FoxO signaling pathway | 4.08 × 10−6 |
| Hippo signaling pathway—multiple species | 4.56 × 10−6 |
| mTOR signaling pathway | 1.06 × 10−5 |
| Fat digestion and absorption | 1.33 × 10−5 |
| Citrate cycle TCA cycle | 2.57 × 10−5 |
| Autophagy—animal | 2.71 × 10−5 |
| Cardiac muscle contraction | 3.04 × 10−5 |
| Phosphatidylinositol signaling system | 3.49 × 10−5 |
| PI3K-Akt signaling pathway | 4.11 × 10−5 |
| MAPK signaling pathway | 4.43 × 10−5 |
| cAMP signaling pathway | 4.51 × 10−5 |
| Pathway | p-Value |
|---|---|
| Fructose and mannose metabolism | 0.0072082 |
| Propanoate metabolism | 0.0082026 |
| Inositol phosphate metabolism | 0.0100905 |
| Carbohydrate digestion and absorption | 0.0118522 |
| Valine, leucine, and isoleucine degradation | 0.0119277 |
| VEGF signaling pathway | 0.013063 |
| Citrate cycle TCA cycle | 0.015308 |
| Arachidonic acid metabolism | 0.0164423 |
| Pyruvate metabolism | 0.0165376 |
| Fatty acid biosynthesis | 0.0197426 |
| Riboflavin metabolism | 0.0218427 |
| Pyrimidine metabolism | 0.0247384 |
| Glucagon signaling pathway | 0.0401259 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hailu, F.T.; Karimpour-Fard, A.; Neltner, B.; Stauffer, B.L.; Lipshultz, S.; Miyamoto, S.D.; Sucharov, C.C. Circulating and Cardiac Tissue miRNAs in Children with Dilated Cardiomyopathy. J. Cardiovasc. Dev. Dis. 2023, 10, 391. https://doi.org/10.3390/jcdd10090391
Hailu FT, Karimpour-Fard A, Neltner B, Stauffer BL, Lipshultz S, Miyamoto SD, Sucharov CC. Circulating and Cardiac Tissue miRNAs in Children with Dilated Cardiomyopathy. Journal of Cardiovascular Development and Disease. 2023; 10(9):391. https://doi.org/10.3390/jcdd10090391
Chicago/Turabian StyleHailu, Frehiwet T., Anis Karimpour-Fard, Bonnie Neltner, Brian L. Stauffer, Steven Lipshultz, Shelley D. Miyamoto, and Carmen C. Sucharov. 2023. "Circulating and Cardiac Tissue miRNAs in Children with Dilated Cardiomyopathy" Journal of Cardiovascular Development and Disease 10, no. 9: 391. https://doi.org/10.3390/jcdd10090391