
Maternal Serum tRNA-Derived Fragments (tRFs) as Potential Candidates for Diagnosis of Fetal Congenital Heart Disease
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Specimen Collection and Processing
2.2. RNA Extraction and Preservation
2.3. Library Preparation and Sequencing
2.4. Bioinformatic Analysis of tRFs/tiRNAs
2.5. Quantitative Real-Time PCR (qRT-PCR) for tRFs/tiRNA
2.6. Receiver Operating Characteristic Analysis
2.7. Functional Analysis of tRFs/tiRNAs
2.8. Statistical Analysis
3. Results
3.1. Catalogue of tRFs/tiRNAs Expressed in Pregnant Women with Fetal Normal Heart and Fetal CHD
3.2. Differentially Expressed tRFs/tiRNAs between Pregnant Women with Fetal Normal Heart and Fetal CHD
3.3. Predictive Value of Specific tRFs/tiRNAs as Diagnostic Biomarkers for CHD
3.4. Potential Targets and Functional Analysis of Target Genes for Predicting Differential Expression of tRFs/tiRNAs
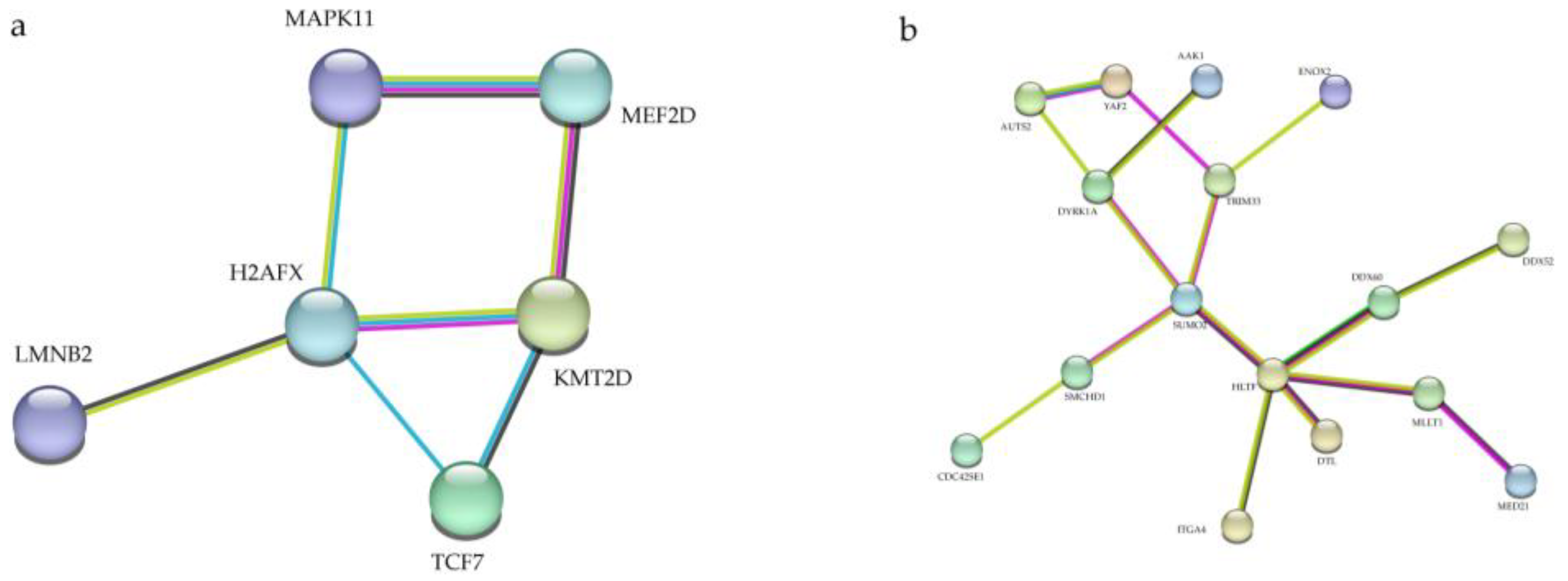
3.5. Protein–Protein Interaction (PPI) Network Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitchell, S.C.; Korones, S.B.; Berendes, H.W. Congenital Heart Disease in 56,109 Births Incidence and Natural History. Circulation 1971, 43, 323–332. [Google Scholar] [CrossRef]
- Dastgiri, S.; Stone, D.H.; Le-Ha, C.; Gilmour, W.H. Prevalence and secular trend of congenital anomalies in Glasgow, UK. Arch. Dis. Child. 2002, 86, 257–263. [Google Scholar] [CrossRef]
- Tennant, P.W.; Pearce, M.S.; Bythell, M.; Rankin, J. 20-year survival of children born with congenital anomalies: A population-based study. Lancet 2010, 375, 649–656. [Google Scholar] [CrossRef]
- van der Linde, D.; Konings, E.E.M.; Slager, M.A.; Witsenburg, M.; Helbing, W.A.; Takkenberg, J.J.M.; Roos-Hesselink, J.W. Birth Prevalence of Congenital Heart Disease Worldwide: A Systematic Review and Meta-Analysis. J. Am. Coll. Cardiol. 2011, 58, 2241–2247. [Google Scholar] [CrossRef]
- Hoffman, J.I.; Kaplan, S.; Liberthson, R.R. Prevalence of congenital heart disease. Am. Hearth J. 2004, 147, 425–439. [Google Scholar] [CrossRef]
- Reller, M.D.; Strickland, M.J.; Riehle-Colarusso, T.; Mahle, W.T.; Correa, A. Prevalence of Congenital Heart Defects in Metropolitan Atlanta, 1998–2005. J. Pediatr. 2008, 153, 807–813. [Google Scholar] [CrossRef]
- Hoffman, J.I.; Kaplan, S. The incidence of congenital heart disease. J. Am. Coll. Cardiol. 2002, 39, 1890–1900. [Google Scholar] [CrossRef]
- Leirgul, E.; Fomina, T.; Brodwall, K.; Greve, G.; Holmstrøm, H.; Vollset, S.E.; Tell, G.S.; Øyen, N. Birth prevalence of congenital heart defects in Norway 1994-2009—A nationwide study. Am. Heart J. 2014, 168, 956–964. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, S.; Zühlke, L.; Black, G.; Choy, M.-K.; Li, N.; Keavney, B.D. Global birth prevalence of congenital heart defects 1970–2017: Updated systematic review and meta-analysis of 260 studies. Leuk. Res. 2019, 48, 455–463. [Google Scholar] [CrossRef]
- Bakker, M.K.; Bergman, J.E.H.; Krikov, S.; Amar, E.; Cocchi, G.; Cragan, J.; Walle, H.E.K.D.; Gatt, M.; Groisman, B.; Liu, S.; et al. Prenatal diagnosis and prevalence of critical congenital heart defects: An international retrospective cohort study. BMJ Open 2019, 9, e028139. [Google Scholar] [CrossRef] [Green Version]
- Donofrio, M.T.; Moon-Grady, A.J.; Hornberger, L.K.; Copel, J.A.; Sklansky, M.S.; Abuhamad, A.; Cuneo, B.F.; Huhta, J.C.; Jonas, R.A.; Krishnan, A.; et al. Diagnosis and Treatment of Fetal Cardiac Disease: A scientific statement from the American Heart Association. Circulation 2014, 129, 2183–2242. [Google Scholar] [CrossRef] [PubMed]
- Sholler, G. Echocardiography in congenital heart disease: Diagnosis, misdiagnosis, and ownership. J. Paediatr. Child Health 2001, 37, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Turton, N.; Swan, R.; Mahenthiralingam, T.; Pitts, D.; Dykes, I.M. The Functions of Long Non-Coding RNA during Embryonic Cardiovascular Development and Its Potential for Diagnosis and Treatment of Congenital Heart Disease. J. Cardiovasc. Dev. Dis. 2019, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Nagy, O.; Baráth, S.; Ujfalusi, A. The role of microRNAs in congenital heart disease. Ejifcc 2019, 30, 165–178. [Google Scholar] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines. Indicates that Thousands of Human Genes are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef]
- Zhu, S.; Cao, L.; Zhu, J.; Kong, L.; Jin, J.; Qian, L.; Zhu, C.; Hu, X.; Li, M.; Guo, X.; et al. Identification of maternal serum microRNAs as novel non-invasive biomarkers for prenatal detection of fetal congenital heart defects. Clin. Chim. Acta Int. J. Clin. Chem. 2013, 424, 66–72. [Google Scholar] [CrossRef]
- Shen, Y.; Yu, X.; Zhu, L.; Li, T.; Yan, Z.; Guo, J. Transfer RNA-derived fragments and tRNA halves: Biogenesis, biological functions and their roles in diseases. J. Mol. Med. 2018, 96, 1167–1176. [Google Scholar] [CrossRef]
- Xie, Y.; Yao, L.; Yu, X.; Ruan, Y.; Li, Z.; Guo, J. Action mechanisms and research methods of tRNA-derived small RNAs. Signal Transduct. Target. Ther. 2020, 5, 109. [Google Scholar] [CrossRef]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef]
- Kumar, P.; Kuscu, C.; Dutta, A. Biogenesis and Function of Transfer RNA-Related Fragments (tRFs). Trends Biochem. Sci. 2016, 41, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Yeung, M.L.; Bennasser, Y.; Watashi, K.; Le, S.-Y.; Houzet, L.; Jeang, K.-T. Pyrosequencing of small non-coding RNAs in HIV-1 infected cells: Evidence for the processing of a viral-cellular double-stranded RNA hybrid. Nucleic Acids Res. 2009, 37, 6575–6586. [Google Scholar] [CrossRef] [PubMed]
- Dhahbi, J.M.; Spindler, S.R.; Atamna, H.; Boffelli, D.; Martin, D.I. Deep Sequencing of Serum Small RNAs Identifies Patterns of 5′ tRNA Half and YRNA Fragment Expression Associated with Breast Cancer. Biomark. Cancer 2014, 6, 37–47. [Google Scholar] [CrossRef]
- Keam, S.P.; Hutvagner, G. tRNA-Derived Fragments (tRFs): Emerging New Roles for an Ancient RNA in the Regulation of Gene Expression. Life 2015, 5, 1638–1651. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, H.; Liu, X.; Nguyen, H.C.; Zhang, S.; Fish, L.; Tavazoie, S.F. Endogenous tRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement. Cell 2015, 161, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Liu, Y.; Yan, Y.; Shen, Y.; Li, Z.; Li, X.; Zhang, Y.; Xue, Z.; Peng, C.; Chen, X.; et al. Differential Expression Profiles and Function Predictions for tRFs & tiRNAs in Skin Injury Induced by Ultraviolet Irradiation. Front. Cell Dev. Biol. 2021, 9, 707572. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, A.E.; Eggens, V.R.C.; Caglayan, A.O.; Reuter, M.S.; Scott, E.; Coufal, N.G.; Silhavy, J.L.; Xue, Y.; Kayserili, H.; Yasuno, K.; et al. CLP1 Founder Mutation Links tRNA Splicing and Maturation to Cerebellar Development and Neurodegeneration. Cell 2014, 157, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Huang, Y.; Wu, X.; Zheng, M.; Xia, Y.; Fu, Z.; Ge, H.; Wang, S.; Xie, H. Hypoxia-induced tRNA-derived fragments, novel regulatory factor for doxorubicin resistance in triple-negative breast cancer. J. Cell. Physiol. 2019, 234, 8740–8751. [Google Scholar] [CrossRef]
- Xu, C.; Fu, Y. Expression Profiles of tRNA-Derived Fragments and Their Potential Roles in Multiple Myeloma. OncoTargets Ther. 2021, 14, 2805–2814. [Google Scholar] [CrossRef]
- Chen, Y.-X.; Ding, J.; Zhou, W.-E.; Zhang, X.; Sun, X.-T.; Wang, X.-Y.; Zhang, C.; Li, N.; Shao, G.-F.; Hu, S.-J.; et al. Identification and Functional Prediction of Long Non-Coding RNAs in Dilated Cardiomyopathy by Bioinformatics Analysis. Front. Genet. 2021, 12, 648111. [Google Scholar] [CrossRef]
- Huang, Y.; Ge, H.; Zheng, M.; Cui, Y.; Fu, Z.; Wu, X.; Xia, Y.; Chen, L.; Wang, Z.; Wang, S.; et al. Serum tRNA-derived fragments (tRFs) as potential candidates for diagnosis of nontriple negative breast cancer. J. Cell. Physiol. 2020, 235, 2809–2824. [Google Scholar] [CrossRef]
- Sun, C.; Yang, F.; Zhang, Y.; Chu, J.; Wang, J.; Wang, Y.; Zhang, Y.; Li, J.; Li, Y.; Fan, R.; et al. tRNA-Derived Fragments as Novel Predictive Biomarkers for Trastuzumab-Resistant Breast Cancer. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 49, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, C.; Huang, S.; Ma, Y.; Fang, J.; Chen, Y. A Downmodulated MicroRNA Profiling in Patients with Gastric Cancer. Gastroenterol. Res. Pract. 2017, 2017, 1526981. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Hong, X.; Zhou, W.; Zhang, Y. Expression profiles of tRNA-derived fragments and their potential roles in ovarian endometriosis. Epigenomics 2020, 12, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liang, T.; Zhang, F.; Liu, J.; Fu, Y. tRNA-derived fragments as novel potential biomarkers for relapsed/refractory multiple myeloma. BMC Bioinform. 2021, 22, 238. [Google Scholar] [CrossRef]
- Martin, M.J.E.j. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Wang, J.; Dong, P.-K.; Xu, X.-F.; Huang, T.; Mao, S.; Wang, Q.-G.; Hao, J.; Liu, X.-H.; Sun, X.-D.; Kang, K.; et al. Identification of tRNA-derived Fragments and Their Potential Roles in Atherosclerosis. Curr. Med. Sci. 2021, 41, 712–721. [Google Scholar] [CrossRef]
- Greiner, M.; Pfeiffer, D.; Smith, R. Principles and practical application of the receiver-operating characteristic analysis for diagnostic tests. Prev. Vet. Med. 2000, 45, 23–41. [Google Scholar] [CrossRef]
- Pencina, M.J.; D’Agostino, R.B., Sr.; D’Agostino, R.B., Jr.; Vasan, R.S. Evaluating the added predictive ability of a new marker: From area under the ROC curve to reclassification and beyond. Stat. Med. 2008, 27, 157–172. [Google Scholar] [CrossRef]
- Yuan, J.; Adamski, R.; Chen, J. Focus on histone variant H2AX: To be or not to be. FEBS Lett. 2010, 584, 3717–3724. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, B.; Kessinger, C.; Kobayashi, J.; Chen, B.P.; Chen, D.J.; Chatterjee, A.; Burma, S. DNA-PK phosphorylates histone H2AX during apoptotic DNA fragmentation in mammalian cells. DNA Repair 2006, 5, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Solier, S.; Sordet, O.; Kohn, K.W.; Pommier, Y. Death Receptor-Induced Activation of the Chk2- and Histone H2AX-Associated DNA Damage Response Pathways. Mol. Cell. Biol. 2009, 29, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhu, F.; Cho, Y.-Y.; Tang, F.; Zykova, T.; Ma, W.-Y.; Bode, A.M.; Dong, Z. Cell Apoptosis: Requirement of H2AX in DNA Ladder Formation, but Not for the Activation of Caspase-3. Mol. Cell 2006, 23, 121–132. [Google Scholar] [CrossRef]
- Lu, C.; Shi, Y.; Wang, Z.; Song, Z.; Zhu, M.; Cai, Q.; Chen, T. Serum starvation induces H2AX phosphorylation to regulate apoptosis via p38 MAPK pathway. FEBS Lett. 2008, 582, 2703–2708. [Google Scholar] [CrossRef]
- Sandhu, S.; Wu, X.; Nabi, Z.; Rastegar, M.; Kung, S.; Mai, S.; Ding, H. Loss of HLTF function promotes intestinal carcinogenesis. Mol. Cancer 2012, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Blue, G.M.; Kirk, E.P.; Giannoulatou, E.; Sholler, G.F.; Dunwoodie, S.L.; Harvey, R.P.; Winlaw, D.S. Advances in the Genetics of Congenital Heart Disease: A Clinician’s Guide. J. Am. Coll. Cardiol. 2017, 69, 859–870. [Google Scholar] [CrossRef] [PubMed]
- McCracken, C.; Spector, L.G.; Menk, J.S.; Knight, J.H.; Vinocur, J.M.; Thomas, A.S.; Oster, M.E.; Louis, J.D.S.; Moller, J.H.; Kochilas, L. Mortality Following Pediatric Congenital Heart Surgery: An Analysis of the Causes of Death Derived From the National Death Index. J. Am. Hearth Assoc. 2018, 7, e010624. [Google Scholar] [CrossRef] [PubMed]
- Krishna, M.R.; Kumar, R.K. Diagnosis and Management of Critical Congenital Heart Diseases in the Newborn. Indian J. Pediatr. 2020, 87, 365–371. [Google Scholar] [CrossRef]
- Kim, H.K.; Fuchs, G.; Wang, S.; Wei, W.; Zhang, Y.; Park, H.; Roy-Chaudhuri, B.; Li, P.; Xu, J.; Chu, K.; et al. A transfer-RNA-derived small RNA regulates ribosome biogenesis. Nature 2017, 552, 57–62. [Google Scholar] [CrossRef]
- Andersen, K.R.; Jensen, T.H.; Brodersen, D.E. Take the “A” tail—quality control of ribosomal and transfer RNA. Biochim. Biophys. Acta 2008, 1779, 532–537. [Google Scholar] [CrossRef]
- Green, D.; Fraser, W.D.; Dalmay, T. Transfer RNA-derived small RNAs in the cancer transcriptome. Pflug. Arch. Eur. J. Physiol. 2016, 468, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Lage, K.; Greenway, S.C.; Rosenfeld, J.A.; Wakimoto, H.; Gorham, J.M.; Segrè, A.V.; Roberts, A.E.; Smoot, L.B.; Pu, W.T.; Pereira, A.C.; et al. Genetic and environmental risk factors in congenital heart disease functionally converge in protein networks driving heart development. Proc. Natl. Acad. Sci. USA 2012, 109, 14035–14040. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Hu, X.; Li, X.; Wen, B.; Wang, L.; Wang, C. Parental non-hereditary teratogenic exposure factors on the occurrence of congenital heart disease in the offspring in the northeastern Sichuan, China. Sci. Rep. 2020, 10, 3905. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Number | Age at Diagnosis | Reproductive History | Gestational Age | Exposure to Harmful Substances | Clinical Diagnosis |
|---|---|---|---|---|---|
| Samples were used for high-throughput sequencing | |||||
| D5 | 26 | G2P1 | 23W + 3D | NO | Tetralogy of Fallot, Pulmonary atresia |
| D25 | 30 | G1P0 | 23W + 2D | NO | Total anomalous pulmonary venous drainage |
| D26 | 31 | G2P1 | 23W + 0D | NO | Endocardial cushion defect |
| D24 | 35 | G1P0 | 23W + 0D | NO | Tetralogy of Fallot |
| D27 | 36 | G1P0 | 24W + 2D | NO | Hypoplastic right heart, Tricuspid Atresia, Pulmonary valve atresia |
| D40 | 33 | G1P0 | 24W + 2D | NO | Tricuspid Atresia, Ventricular septal defect |
| Samples were used for follow-up qRT-PCR | |||||
| A01 | 29 | G1P0 | 25W + 5D | NO | Transposition of great arteries, Ventricular septal defect, Aortic valve stenosis with insufficiency of closure |
| A02 | 29 | G1P0 | 28W + 0D | NO | Aortic valve stenosis, Right-sided aortic arch |
| A03 | 30 | G1P0 | 22W + 6D | NO | Double outlet of the right ventricle |
| A04 | 34 | G3P1 | 24W + 4D | NO | Tetralogy of Fallot |
| A05 | 29 | G1P0 | 31W + 3D | NO | Ventricular septal defect |
| A06 | 27 | G1P0 | 33W + 3D | NO | Pulmonary valve stenosis with incomplete closure |
| A07 | 30 | G2P1 | 25W + 1D | NO | Total anomalous pulmonary venous drainage |
| A08 | 34 | G2P1 | 26W + 3D | NO | Pulmonary artery crossover |
| A09 | 38 | G5P1 | 23W + 6D | NO | Pulmonary valve stenosis |
| A10 | 32 | G2P1 | 27W + 3D | NO | Umbilical vein—coronary sinus—right atrium link |
| A11 | 36 | G2P1 | 28W + 0D | NO | The right heart is larger than the left heart |
| A12 | 23 | G1P0 | 25W + 6D | NO | Ventricular septal defect, Right-sided aortic arch |
| A13 | 39 | G3P1 | 33W + 0D | NO | The right heart is larger than the left heart, Aortic arch narrowing |
| A14 | 40 | G5P4 | 24W + 3D | NO | Hypoplastic left heart syndrome |
| A15 | 29 | G1P0 | 24W + 1D | NO | Pulmonary valve stenosis |
| A16 | 34 | G2P0 | 28W + 6D | NO | The right heart is larger than the left heart |
| A17 | 29 | G2P0 | 24W + 2D | NO | Moderate tricuspid regurgitation |
| A18 | 30 | G2P1 | 23W + 5D | NO | Transposition of the great arteries |
| A19 | 29 | G1P0 | 24W + 6D | NO | Ventricular septal defect, Coarctation of the Aorta, Transposition of the great arteries |
| A20 | 33 | G4P1 | 25W + 5D | NO | Abnormal development of the aortic arch |
| A21 | 31 | G1P0 | 34W + 5D | NO | Hypoplastic right heart |
| A22 | 33 | G2P1 | 25W + 3D | NO | The right ventricle is smaller than the left ventricle |
| A23 | 29 | G1P0 | 24W + 5D | NO | Aortic arch stenosis |
| A24 | 28 | G1P0 | 27W + 4D | NO | Aortic arch stenosis |
| A25 | 29 | G2P1 | 28W + 5D | NO | Small left atrium |
| A26 | 33 | G1P0 | 25W + 3D | NO | Complete atrial septal defect |
| A27 | 33 | G2P1 | 24W + 2D | NO | Ventricular septal defect, Aortic arch stenosis, Possible aortic arch dissection |
| A28 | 31 | G3P2 | 27W + 3D | NO | Mid-large tricuspid regurgitation |
| A29 | 27 | G2P0 | 22W + 3D | NO | Total anomalous pulmonary venous drainage |
| A30 | 27 | G1P0 | 25W + 3D | NO | Ventricular septal defect |
| A31 | 24 | G1P0 | 25W + 4D | NO | Transposition of the great arteries, Ventricular septal defect |
| A32 | 29 | G1P0 | 24W + 6D | NO | Right-sided aortic arch |
| A33 | 31 | G1P0 | 24W + 2D | NO | Ventricular septal defect, Aorta riding across |
| A34 | 29 | G1P0 | 28W + 2D | NO | Ventricular septal defect |
| A35 | 31 | G3P1 | 24W + 5D | NO | Ventricular septal defect, Aortic valve stenosis |
| Sample Number | Age at Diagnosis | Reproductive History | Gestational Age | Exposure to Harmful Substances | Whether the Fetal Heart Is Healthy |
|---|---|---|---|---|---|
| Samples were used for high-throughput sequencing | |||||
| C28 | 34 | G2P1 | 23W + 6D | NO | YES |
| C29 | 28 | G1P0 | 24W + 1D | NO | YES |
| C30 | 31 | G2P0 | 23W + 6D | NO | YES |
| C31 | 28 | G1P0 | 23W + 6D | NO | YES |
| C32 | 27 | G1P0 | 24W + 0D | NO | YES |
| C35 | 28 | G1P0 | 23W + 6D | NO | YES |
| Samples were used for follow-up qRT-PCR | |||||
| B1 | 34 | - | - | NO | YES |
| B2 | 35 | - | - | NO | YES |
| B3 | 34 | - | - | NO | YES |
| B4 | 36 | - | - | NO | YES |
| B5 | 33 | - | - | NO | YES |
| B6 | 28 | - | - | NO | YES |
| B7 | 26 | - | - | NO | YES |
| B8 | 36 | - | - | NO | YES |
| B9 | 34 | - | - | NO | YES |
| B10 | 36 | - | - | NO | YES |
| B11 | 32 | - | - | NO | YES |
| B12 | 33 | - | - | NO | YES |
| B13 | 48 | - | - | NO | YES |
| B14 | 30 | - | - | NO | YES |
| B15 | 31 | - | - | NO | YES |
| B16 | 31 | - | - | NO | YES |
| B17 | 31 | - | - | NO | YES |
| B18 | 28 | - | - | NO | YES |
| B19 | 36 | - | - | NO | YES |
| B20 | 36 | - | - | NO | YES |
| B21 | 35 | - | - | NO | YES |
| B22 | 31 | - | - | NO | YES |
| B23 | 18 | - | - | NO | YES |
| B24 | 34 | - | - | NO | YES |
| B25 | 30 | - | - | NO | YES |
| B26 | 36 | - | - | NO | YES |
| B27 | 36 | - | - | NO | YES |
| B28 | 32 | - | - | NO | YES |
| B29 | 27 | - | - | NO | YES |
| B30 | 28 | - | - | NO | YES |
| B31 | 31 | - | - | NO | YES |
| B32 | 31 | - | - | NO | YES |
| B33 | 32 | - | - | NO | YES |
| B34 | 33 | - | - | NO | YES |
| B35 | 32 | - | - | NO | YES |
| B36 | 41 | - | - | NO | YES |
| B37 | 30 | - | - | NO | YES |
| B38 | 30 | - | - | NO | YES |
| B39 | 30 | - | - | NO | YES |
| B40 | 31 | - | - | NO | YES |
| B41 | 34 | - | - | NO | YES |
| B42 | 32 | - | - | NO | YES |
| B43 | 27 | - | - | NO | YES |
| B44 | 30 | - | - | NO | YES |
| B45 | 33 | - | - | NO | YES |
| B46 | 26 | - | - | NO | YES |
| B47 | 27 | - | - | NO | YES |
| B48 | 27 | - | - | NO | YES |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, E.; Wu, L.; Chen, B.; Xu, S.; Fu, Z.; Wu, Y.; Wu, Y.; Gu, H. Maternal Serum tRNA-Derived Fragments (tRFs) as Potential Candidates for Diagnosis of Fetal Congenital Heart Disease. J. Cardiovasc. Dev. Dis. 2023, 10, 78. https://doi.org/10.3390/jcdd10020078
Lu E, Wu L, Chen B, Xu S, Fu Z, Wu Y, Wu Y, Gu H. Maternal Serum tRNA-Derived Fragments (tRFs) as Potential Candidates for Diagnosis of Fetal Congenital Heart Disease. Journal of Cardiovascular Development and Disease. 2023; 10(2):78. https://doi.org/10.3390/jcdd10020078
Chicago/Turabian StyleLu, Enkang, Lijun Wu, Bin Chen, Shipeng Xu, Ziyi Fu, Yun Wu, Yanhu Wu, and Haitao Gu. 2023. "Maternal Serum tRNA-Derived Fragments (tRFs) as Potential Candidates for Diagnosis of Fetal Congenital Heart Disease" Journal of Cardiovascular Development and Disease 10, no. 2: 78. https://doi.org/10.3390/jcdd10020078