
A Review on Pathological and Diagnostic Aspects of Emerging Viruses—Senecavirus A, Torque teno sus virus and Linda Virus—In Swine
 , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Senecavirus A
3. Torque teno sus virus (TTSuV)
4. Lateral Shaking Inducing Neuro-Degenerative Agent (LindaV)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- González, N.; Marquès, M.; Nadal, M.; Domingo, J.L. Meat consumption: Which are the current global risks? A review of recent (2010–2020) evidences. Food Res. Int. 2020, 137, 109341. [Google Scholar] [CrossRef] [PubMed]
- VanderWaal, K.; Deen, J. Global trends in infectious diseases of swine. Proc. Natl. Acad. Sci. USA 2018, 115, 11495–11500. [Google Scholar] [CrossRef] [PubMed]
- Augère-Granier, M.L. The EU Pig Meat Sector. EPRS|European Parliamentary Research Service. 2020, pp. 1–20. Available online: https://www.europarl.europa.eu/RegData/etudes/BRIE/2020/652044/EPRS_BRI (accessed on 10 February 2022).
- Ito, S.; Bosch, J.; Martínez-Avilés, M.; Sánchez-Vizcaíno, J.M. The Evolution of African Swine Fever in China: A Global Threat? Front. Vet. Sci. 2022, 9, 828498. [Google Scholar] [CrossRef]
- Bonazzi, G.; Camanzi, P.; Ferri, G.; Manghi, E.; Iotti, M. Economic Sustainability of Pig Slaughtering Firms in the Production Chain of Denomination of Origin Hams in Italy. Sustainability 2021, 13, 7639. [Google Scholar] [CrossRef]
- Alarcón, L.V.; Allepuz, A.; Mateu, E. Biosecurity in pig farms: A review. Porc. Health Manag. 2021, 7, 181. [Google Scholar] [CrossRef]
- Espinosa, R.; Tago, D.; Treich, N. Infectious Diseases and Meat Production. Environ. Resour. Econ. 2020, 76, 1019–1044. [Google Scholar] [CrossRef]
- Drew, T.W. The emergence and evolution of swine viral diseases: To what extent have husbandry systems and global trade contributed to their distribution and diversity? Rev. Sci. Tech. Oie 2011, 30, 95–106. [Google Scholar] [CrossRef]
- Nathues, H.; Alarcon, P.; Rushton, J.; Jolie, R.; Fiebig, K.; Jimenez, M.; Geurts, V. Cost of porcine reproductive and respiratory syndrome virus at individual farm level–An economic disease model. Prev. Vet. Med. 2017, 142, 16–29. [Google Scholar] [CrossRef]
- Baekbo, P.; Kristensen, C.S.; Larsen, L.E. Porcine Circovirus Diseases: A review of PMWS. Transbound. Emerg. Dis. 2012, 59, 60–67. [Google Scholar] [CrossRef]
- Vidigal, P.M.; Mafra, C.L.; Silva, F.M.; Fietto, J.L.; Júnior, A.S.; Almeida, M.R. Tripping over emerging pathogens around the world: A phylogeographical approach for determining the epidemiology of Porcine circovirus-2 (PCV-2), considering global trading. Virus Res. 2012, 163, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Gaudreault, N.N.; Madden, D.W.; Wilson, W.C.; Trujillo, J.D.; Richt, J.A. African Swine Fever Virus: An Emerging DNA Arbovirus. Front. Vet. Sci. 2020, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- Stenfeldt, C.; Diaz-San Segundo, F.; de Los Santos, T.; Rodriguez, L.L.; Earzt, J. The Pathogenesis of Foot-and-Mouth Disease in Pigs. Front. Vet. Sci. 2016, 3, 41. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J. Emerging and Re-emerging Swine Viruses. Transbound. Emerg. Dis. 2012, 59, 85–102. [Google Scholar] [CrossRef] [PubMed]
- Perfumo, C.J.; Pereda, A.; Jongkaewwattana, A.; Chen, Z.; Perez, D.R.; Ma, J. Editorial: Emerging Swine Viruses. Front. Vet. Sci. 2020, 7, 132. [Google Scholar] [CrossRef]
- Trovato, M.; Sartorius, R.; D’Apice, L.; Manco, R.; De Berardinis, P. Viral Emerging Diseases: Challenges in Developing Vaccination Strategies. Front. Immunol. 2020, 11, 2130. [Google Scholar] [CrossRef]
- Hales, L.M.; Knowles, N.J.; Reddy, P.S.; Xu, L.; Hay, C.; Hallenbeck, P.L. Complete genome sequence analysis of Seneca Valley virus-001, a novel oncolytic picornavirus. J. Gen. Virol. 2008, 89, 1265–1275. [Google Scholar] [CrossRef]
- Leme, R.A.; Alfieri, A.F.; Alfieri, A.A. Update on Senecavirus Infection in Pigs. Viruses 2017, 9, 170. [Google Scholar] [CrossRef]
- Venkataraman, S.; Reddy, S.P.; Loo, J.; Idamakanti, N.; Hallenbeck, P.L.; Reddy, V.S. Structure of Seneca Valley Virus-001: An Oncolytic Picornavirus Representing a New Genus. Structure 2008, 16, 1555–1561. [Google Scholar] [CrossRef]
- Li, C.; Wang, H.; Shi, J.; Yang, D.; Zhou, G.; Chang, J.; Cameron, C.E.; Woodman, A.; Yu, L. Senecavirus-Specific Recombination Assays Reveal the Intimate Link between Polymerase Fidelity and RNA Recombination. J. Virol. 2019, 93, 576. [Google Scholar] [CrossRef]
- Knowles, N.J.; Hales, B.H.; Jones, J.G.; Landgraf, J.A.; House, K.L.; Skele, K.D.; Burroughs, K.D.; Hallenbeck, P.L. Epidemiology of Seneca Valley virus: Identification and characterization of isolates from pigs in the United States, p G2. In Proceedings of the EUROPIC 2006: XIVth Meeting of the European Study on Molecular Biology of Picornaviruses Saariselka, Inari, Finland, 26 November–1 December 2006. [Google Scholar]
- Pasma, T.; Davidson, S.; Shaw, S.L. Idiopathic vesicular disease in swine in Manitoba. Can. Vet. J. Rev. Vet. Can. 2008, 49, 84–85. [Google Scholar]
- Singh, K.; Corner, S.; Clark, S.G.; Scherba, G.; Fredrickson, R. Seneca Valley virus and vesicular lesions in a pig with idiopathic vesicular disease. J. Vet. Sci. Technol. 2012, 3, 1–3. [Google Scholar]
- Vannucci, F.A.; Linhares, D.C.L.; de Barcellos, D.E.S.N.; Lam, H.C.; Collins, J.; Marthaler, D. Identification and Complete Genome of Seneca Valley Virus in Vesicular Fluid and Sera of Pigs Affected with Idiopathic Vesicular Disease, Brazil. Transbound. Emerg. Dis. 2015, 62, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Saporiti, V.; Fritzen, J.T.T.; Feronato, C.; Leme, R.; Lobato, Z.; Alfieri, A.; Alfieri, A.A. A ten years (2007–2016) retrospective serological survey for Seneca Valley virus infection in major pig producing states of Brazil. Vet. Res. Commun. 2017, 41, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Piñeyro, P.; Chen, Q.; Zheng, Y.; Li, G.; Rademacher, C.; Derscheid, R.; Guo, B.; Yoon, K.-J.; Madson, D.; et al. Full-Length Genome Sequences of Senecavirus A from Recent Idiopathic Vesicular Disease Outbreaks in U.S. Swine. Genome Announc. 2015, 3, e01270-15. [Google Scholar] [CrossRef]
- Baker, K.L.; Mowrer, C.; Canon, A.; Linhares, D.C.L.; Rademacher, C.; Karriker, L.A.; Holtkamp, D.J. Systematic Epidemiological Investigations of Cases of Senecavirus A in US Swine Breeding Herds. Transbound. Emerg. Dis. 2017, 64, 11–18. [Google Scholar] [CrossRef]
- Canning, P.; Canon, A.; Bates, J.L.; Gerardy, K.; Linhares, D.C.L.; Piñeyro, P.E.; Schwartz, K.J.; Yoon, K.J.; Rademacher, C.J.; Holtkamp, D.; et al. Neonatal Mortality, Vesicular Lesions and Lameness Associated with Senecavirus A in a U.S. Sow Farm. Transbound. Emerg. Dis. 2016, 63, 373–378. [Google Scholar] [CrossRef]
- Wang, L.; Prarat, M.; Hayes, J.; Zhang, Y. Detection and Genomic Characterization of Senecavirus A, Ohio, USA, 2015. Emerg. Infect. Dis. 2016, 22, 1321–1323. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, X.; Chen, Y.; He, X.; Zhang, G.; Ma, J. Complete Genome Sequence of Seneca Valley Virus CH-01-2015 Identified in China. Genome Announc. 2016, 4, e01509-15. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, X.; Bai, Y.; Sun, B.; Xie, Q.; Ma, J. The First Identification and Complete Genome of Senecavirus A Affecting Pig with Idiopathic Vesicular Disease in China. Transbound. Emerg. Dis. 2017, 64, 1633–1640. [Google Scholar] [CrossRef]
- Wang, M.; Chen, L.; Pan, S.; Mou, C.; Shi, K.; Chen, Z. Molecular evolution and characterization of novel Seneca Valley virus (SVV) strains in South China. Infect. Genet. Evol. 2019, 69, 58649665. [Google Scholar] [CrossRef]
- Liu, J.; Zha, Y.; Li, H.; Sun, Y.; Wang, F.; Lu, R.; Ning, Z. Novel Recombinant Seneca Valley Virus Isolated from Slaughtered Pigs in Guangdong Province. Virol. Sin. 2019, 34, 722–724. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, X.-X.; Ruan, H.; Qiao, S.; Deng, R.; Zhang, G. Isolation of Three Novel Senecavirus A Strains and Recombination Analysis Among Senecaviruses in China. Front. Vet. Sci. 2020, 7, 2. [Google Scholar] [CrossRef]
- Sun, D.; Vannucci, F.; Knutson, T.P.; Corzo, C.; Marthaler, D. Emergence and whole-genome sequence of Senecavirus A in Colombia. Transbound. Emerg. Dis. 2017, 64, 1346–1349. [Google Scholar] [CrossRef]
- Saeng-Chuto, K.; Rodtian, P.; Temeeyasen, G.; Wegner, M.; Nilubol, D. The first detection of Senecavirus A in pigs in Thailand, 2016. Transbound. Emerg. Dis. 2017, 65, 285–288. [Google Scholar] [CrossRef]
- Arzt, J.; Bertram, M.R.; Vu, L.T.; Pauszek, S.J.; Hartwig, E.J.; Smoliga, G.R.; Palinski, R.; Stenfeldt, C.; Fish, I.H.; Hoang, B.H.; et al. First Detection and Genome Sequence of Senecavirus A in Vietnam. Microbiol. Resour. Announc. 2019, 8, e01247-18. [Google Scholar] [CrossRef]
- Guo, B.; Piñeyro, P.E.; Rademacher, C.J.; Zheng, Y.; Li, G.; Yuan, J.; Hoang, H.; Gauger, P.C.; Madson, D.M.; Schwartz, K.J.; et al. Novel Senecavirus A in Swine with Vesicular Disease, United States, July 2015. Emerg. Infect. Dis. 2016, 22, 1325–1327. [Google Scholar] [CrossRef]
- Joshi, L.; Mohr, K.A.; Clement, T.; Hain, K.S.; Myers, B.; Yaros, J.; Nelson, E.A.; Christopher-Hennings, J.; Gava, D.; Schaefer, R.; et al. Detection of the Emerging Picornavirus Senecavirus A in Pigs, Mice, and Houseflies. J. Clin. Microbiol. 2016, 54, 1536–1545. [Google Scholar] [CrossRef]
- Leme, R.A.; Miyabe, F.M.; Agnol, A.M.D.; Alfieri, A.F.; Alfieri, A. A new wave of Seneca Valley virus outbreaks in Brazil. Transbound. Emerg. Dis. 2019, 66, 1101–1104. [Google Scholar] [CrossRef]
- Montiel, N.; Buckley, A.; Guo, B.; Kulshreshtha, V.; VanGeelen, A.; Hoang, H.; Rademacher, C.; Yoon, K.-J.; Lager, K. Vesicular Disease in 9-Week-Old Pigs Experimentally Infected with Senecavirus A. Emerg. Infect. Dis. 2016, 22, 1246–1248. [Google Scholar] [CrossRef]
- Joshi, L.; Fernandes, M.H.V.; Clement, T.; Lawson, S.; Pillatzki, A.; Resende, T.P.; Vannucci, F.A.; Kutish, G.F.; Nelson, E.A.; Diel, D.G. Pathogenesis of Senecavirus A infection in finishing pigs. J. Gen. Virol. 2016, 97, 3267–3279. [Google Scholar] [CrossRef]
- Segalés, J.; de Barcellos, D.E.S.N.; Alfieri, A.; Burrough, E.; Marthaler, D. Senecavirus A: An Emerging Pathogen Causing Vesicular Disease and Mortality in Pigs? Vet. Pathol. 2016, 54, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Leme, R.; Oliveira, T.; Alfieri, A.; Headley, S. Pathological, Immunohistochemical and Molecular Findings Associated with Senecavirus A-Induced Lesions in Neonatal Piglets. J. Comp. Pathol. 2016, 155, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, T.E.S.; Michelazzo, M.M.Z.; Fernandes, T.; De Oliveira, A.G.; Leme, R.A.; Alfieri, A.F.; Alfieri, A.A.; Headley, S.A. Histopathological, immunohistochemical, and ultrastructural evidence of spontaneous Senecavirus A-induced lesions at the choroid plexus of newborn piglets. Sci. Rep. 2017, 7, 16555. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, Y.; Li, X.; Liang, L.; Cui, S. Pathogenicity Analysis of Weaned Piglets Challenged With Novel Emerging Senecavirus A in Fujian, China. Front. Vet. Sci. 2021, 8, 694110. [Google Scholar] [CrossRef]
- Gimenez-Lirola, L.G.; Rademacher, C.; Linhares, D.; Harmon, K.; Rotolo, M.; Sun, Y.; Baum, D.H.; Zimmerman, J.; Piñeyro, P. Serological and Molecular Detection of Senecavirus A Associated with an Outbreak of Swine Idiopathic Vesicular Disease and Neonatal Mortality. J. Clin. Microbiol. 2016, 54, 2082–2089. [Google Scholar] [CrossRef]
- Houston, E.; Temeeyasen, G.; Piñeyro, P.E. Comprehensive review on immunopathogenesis, diagnostic and epidemiology of Senecavirus A. Virus Res. 2020, 286, 198038. [Google Scholar] [CrossRef]
- Bracht, A.J.; O’Hearn, E.S.; Fabian, A.W.; Barrette, R.W.; Sayed, A. Real-Time Reverse Transcription PCR Assay for Detection of Senecavirus A in Swine Vesicular Diagnostic Specimens. PLoS ONE 2016, 11, e0146211. [Google Scholar] [CrossRef]
- Fowler, V.L.; Ransburgh, R.H.; Poulsen, E.G.; Wadsworth, J.; King, D.; Mioulet, V.; Knowles, N.J.; Williamson, S.; Liu, X.; Anderson, G.A.; et al. Development of a novel real-time RT-PCR assay to detect Seneca Valley virus-1 associated with emerging cases of vesicular disease in pigs. J. Virol. Methods 2017, 239, 34–37. [Google Scholar] [CrossRef]
- Agnol, A.M.D.; Otonel, R.A.A.; Leme, R.A.; Alfieri, A.; Alfieri, A. A TaqMan-based qRT-PCR assay for Senecavirus A detection in tissue samples of neonatal piglets. Mol. Cell. Probes 2017, 33, 28–31. [Google Scholar] [CrossRef]
- Feronato, C.; Leme, R.A.; Diniz, J.A.; Agnol, A.M.D.; Alfieri, A.F.; Alfieri, A.A. Development and evaluation of a nested-PCR assay for Senecavirus A diagnosis. Trop. Anim. Health Prod. 2017, 50, 337–344. [Google Scholar] [CrossRef]
- Armson, B.; Walsh, C.; Morant, N.; Fowler, V.L.; Knowles, N.J.; Clark, D. The development of two field-ready reverse transcription loop-mediated isothermal amplification assays for the rapid detection of Seneca Valley virus 1. Transbound. Emerg. Dis. 2018, 66, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Cong, F.; Liu, X.; Lian, Y.; Wu, M.; Xiao, L.; Yuan, W.; Huang, R.; Ma, J.; Guo, P.; et al. Development of a real time loop-mediated isothermal amplification method for detection of Senecavirus A. J. Virol. Methods 2018, 261, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro-De-Oliveira, T.F.; Fonseca-Júnior, A.A.; Camargos, M.F.; Laguardia-Nascimento, M.; Giannattasio-Ferraz, S.; Cottorello, A.C.P.; de Oliveira, A.M.; Góes-Neto, A.; Barbosa-Stancioli, E.F. Reverse transcriptase droplet digital PCR to identify the emerging vesicular virus Senecavirus A in biological samples. Transbound. Emerg. Dis. 2019, 66, 1360–1369. [Google Scholar] [CrossRef]
- Zhang, J.; Nfon, C.; Tsai, C.-F.; Lee, C.-H.; Fredericks, L.; Chen, Q.; Sinha, A.; Bade, S.; Harmon, K.; Piñeyro, P.; et al. Development and evaluation of a real-time RT-PCR and a field-deployable RT-insulated isothermal PCR for the detection of Seneca Valley virus. BMC Vet. Res. 2019, 15, 12917. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Y.; Lin, X.; Chen, Z.; Wu, S. Development of a novel reverse transcription droplet digital PCR assay for the sensitive detection of Senecavirus A. Transbound. Emerg. Dis. 2018, 66, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Resende, T.P.; Marthaler, D.G.; Vannucci, F.A. A novel RNA-based in situ hybridization to detect Seneca Valley virus in neonatal piglets and sows affected with vesicular disease. PLoS ONE 2017, 12, e0173190. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, C.M.T.; Akkutay-Yoldar, Z.; Stone, S.R.; Tousignant, S.J.; Vannucci, F.A.; Murtaugh, M.P. An indirect enzyme-linked immunosorbent assay for the identification of antibodies to Senecavirus A in swine. BMC Vet. Res. 2016, 13, 967. [Google Scholar] [CrossRef] [PubMed]
- Goolia, M.; Vannucci, F.; Yang, M.; Patnayak, D.; Babiuk, S.; Nfon, C.K. Validation of a competitive ELISA and a virus neutralization test for the detection and confirmation of antibodies to Senecavirus A in swine sera. J. Vet. Diagn. Investig. 2017, 29, 250–253. [Google Scholar] [CrossRef]
- Yang, M.; van Bruggen, R.; Xu, W. Generation and diagnostic application of monoclonal antibodies against Seneca Valley virus. J. Vet. Diagn. Investig. 2011, 24, 42–50. [Google Scholar] [CrossRef]
- Li, J.; Liang, W.; Xu, S.; Shi, J.; Zhou, X.; Liu, B.; Yu, L.; Xiong, J.; Si, G.; He, D. Rapid and sensitive detection of Senecavirus A by reverse transcription loop-mediated isothermal amplification combined with a lateral flow dipstick method. PLoS ONE 2019, 14, e0216245. [Google Scholar] [CrossRef]
- Wang, H.; Dong, J.; Zhang, T.; Wang, F.; Yang, R.; Zhang, Y.; Zhao, X. A novel rapid detection of Senecavirus A using recombinase polymerase amplification (RPA) coupled with lateral flow (LF) dipstrip. Anal. Biochem. 2022, 646, 114627. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Takahashi, M.; Nishizawa, T.; Tawara, A.; Fukai, K.; Muramatsu, U.; Naito, Y.; Yoshikawa, A. Genomic characterization of TT viruses (TTVs) in pigs, cats and dogs and their relatedness with species-specific TTVs in primates and tupaias. J. Gen. Virol. 2002, 83, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Manzin, A.; Mallus, F.; Macera, L.; Maggi, F.; Blois, S. Global impact of Torque teno virus infection in wild and domesticated animals. J. Infect. Dev. Ctries. 2015, 9, 562–570. [Google Scholar] [CrossRef] [PubMed]
- International Committee on Taxonomy of Viruses. Available online: https://talk.ictvonline.org/ictv-reports/ictv_9th_report/ssdna-viruses-2011/w/ssdna_viruses/139/anelloviridae (accessed on 28 June 2022).
- Varsani, A.; Opriessnig, T.; Celer, V.; Maggi, F.; Okamoto, H.; Blomström, A.-L.; Cadar, D.; Harrach, B.; Biagini, P.; Kraberger, S. Taxonomic update for mammalian anelloviruses (family Anelloviridae). Arch Virol. 2021, 166, 2943–2953. [Google Scholar] [CrossRef]
- Maggi, F.; Bendinelli, M. Immunobiology of the Torque Teno Viruses and Other Anelloviruses. Curr. Top Microbiol. 2009, 331, 65–90. [Google Scholar] [CrossRef]
- Li, G.; Zhang, W.; Wang, R.; Xing, G.; Wang, S.; Ji, X.; Wang, N.; Su, S.; Zhou, J. Genetic Analysis and Evolutionary Changes of the Torque teno sus Virus. Int. J. Mol. Sci. 2019, 20, 2881. [Google Scholar] [CrossRef]
- Kekarainen, T.; Segalés, J. Torque Teno Sus Virus in Pigs: An Emerging Pathogen? Transbound. Emerg. Dis. 2012, 59, 103–108. [Google Scholar] [CrossRef]
- Vargas-Ruiz, A.; Ramírez-Álvarez, H.; I Sánchez-Betancourt, J.; Quintero-Ramírez, V.; Rangel-Rodríguez, I.C.; A Vázquez-Perez, J.; A García-Camacho, L. Retrospective study of the relationship of Torque teno sus virus 1a and Torque teno sus virus 1b with porcine circovirus associated disease. Can. J. Vet. Res. Rev. Can. Rech. Vet. 2017, 81, 178–185. [Google Scholar]
- Kekarainen, T.; López-Soria, S.; Segalés, J. Detection of swine Torque teno virus genogroups 1 and 2 in boar sera and semen. Theriogenology 2007, 68, 966–971. [Google Scholar] [CrossRef]
- Cortey, M.; Pileri, E.; Segalés, J.; Kekarainen, T. Globalisation and global trade influence molecular viral population genetics of Torque Teno Sus Viruses 1 and 2 in pigs. Vet. Microbiol. 2012, 156, 81–87. [Google Scholar] [CrossRef]
- Cadar, D.; Kiss, T.; Ádám, D.; Cságola, A.; Novosel, D.; Tuboly, T. Phylogeny, spatio-temporal phylodynamics and evolutionary scenario of Torque teno sus virus 1 (TTSuV1) and 2 (TTSuV2) in wild boars: Fast dispersal and high genetic diversity. Vet. Microbiol. 2013, 166, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Subramanyam, V.; Hemadri, D.; Kashyap, S.P.; Hiremath, J.; Barman, N.N.; Ralte, E.L.; Patil, S.; Suresh, K.P.; Rahaman, H. Detection of torque teno sus virus infection in Indian pigs. Vet. World 2019, 12, 1467–1471. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J.; Kekarainen, T. Anelloviruses. In Zimmerman J.J, Karriker L.A, Ramirez A, Schwartz K.J, Stevenson G.W, Zhang J, editors, 11th ed.; John Wiley and Sons, Inc.: Boca Ration, NJ, USA, 2019; pp. 453–456. [Google Scholar]
- Leary, T.P.; Erker, J.C.; Chalmers, M.L.; Desai, S.M.; Mushahwar, I.K. Improved detection systems for TT virus reveal high prevalence in humans, non-human primates and farm animals. J. Gen. Virol. 1999, 80, 2115–2120. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J.; Martinezguino, L.; Cortey, M.; Navarro, N.; Huerta, E.; Sibila, M.; Pujols, J.; Kekarainen, T. Retrospective study on swine Torque teno virus genogroups 1 and 2 infection from 1985 to 2005 in Spain. Vet. Microbiol. 2009, 134, 199–207. [Google Scholar] [CrossRef] [PubMed]
- McKeown, N.; Fenaux, M.; Halbur, P.; Meng, X. Molecular characterization of porcine TT virus, an orphan virus, in pigs from six different countries. Vet. Microbiol. 2004, 104, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Martelli, F.; Caprioli, A.; Di Bartolo, I.; Cibin, V.; Pezzotti, G.; Ruggeri, F.M.; Ostanello, F. Detection of Swine Torque Teno Virus in Italian Pig Herds. J. Vet. Med. Ser. B 2006, 53, 234–238. [Google Scholar] [CrossRef]
- Taira, O.; Ogawa, H.; Nagao, A.; Tuchiya, K.; Nunoya, T.; Ueda, S. Prevalence of swine Torque teno virus genogroups 1 and 2 in Japanese swine with suspected post-weaning multisystemic wasting syndrome and porcine respiratory disease complex. Vet. Microbiol. 2009, 139, 347–350. [Google Scholar] [CrossRef]
- Gallei, A.; Pesch, S.; Esking, W.S.; Keller, C.; Ohlinger, V.F. Porcine Torque teno virus: Determination of viral genomic loads by genogroup-specific multiplex rt-PCR, detection of frequent multiple infections with genogroups 1 or 2, and establishment of viral full-length sequences. Vet. Microbiol. 2010, 143, 202–212. [Google Scholar] [CrossRef]
- Cornelissen-Keijsers, V.; Jiménez-Melsió, A.; Sonnemans, D.; Cortey, M.; Segalés, J.; Born, E.V.D.; Kekarainen, T. Discovery of a novel Torque teno sus virus species: Genetic characterization, epidemiological assessment and disease association. J. Gen. Virol. 2012, 93, 2682–2691. [Google Scholar] [CrossRef]
- Mei, M.; Zhu, L.; Xu, Z.; Zhao, L.; Zhou, Y.; Wu, Y.; Li, S.; Wei, H.; Guo, W. Molecular investigation of Torque teno sus virus in geographically distinct porcine breeding herds of Sichuan, China. Virol. J. 2013, 10, 161–169. [Google Scholar] [CrossRef]
- Rogers, A.J.; Huang, Y.-W.; Heffron, C.L.; Opriessnig, T.; Patterson, A.R.; Meng, X.-J. Prevalence of the Novel Torque Teno Sus Virus Species k2b from Pigs in the United States and Lack of Association with Post-Weaning Multisystemic Wasting Syndrome or Mulberry Heart Disease. Transbound. Emerg. Dis. 2016, 64, 1877–1883. [Google Scholar] [CrossRef] [PubMed]
- Polster, S.; Lechmann, J.; Lienhard, J.; Peltzer, D.; Prähauser, B.; Bachofen, C.; Seehusen, F. First Report of TTSuV1 in Domestic Swiss Pigs. Viruses 2022, 14, 870. [Google Scholar] [CrossRef]
- Martínez, L.; Kekarainen, T.; Sibila, M.; Ruiz-Fons, F.; Vidal, D.; Gortázar, C.; Segalés, J. Torque teno virus (TTV) is highly prevalent in the European wild boar (Sus scrofa). Vet. Microbiol. 2006, 118, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Righi, F.; Arnaboldi, S.; Filipello, V.; Ianiro, G.; Di Bartolo, I.; Calò, S.; Bellini, S.; Trogu, T.; Lelli, D.; Bianchi, A.; et al. Torque Teno Sus Virus (TTSuV) Prevalence in Wild Fauna of Northern Italy. Microorganisms 2022, 10, 242. [Google Scholar] [CrossRef] [PubMed]
- Nieto, D.; Aramouni, M.; Roma, L.G.; Segalés, J.; Kekarainen, T. Dynamics of Torque teno sus virus 1 (TTSuV1) and 2 (TTSuV2) DNA loads in serum of healthy and postweaning multisystemic wasting syndrome (PMWS) affected pigs. Vet. Microbiol. 2011, 152, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Aramouni, M.; Segalés, J.; Sibila, M.; Martin-Valls, G.; Nieto, D.; Kekarainen, T. Torque teno sus virus 1 and 2 viral loads in postweaning multisystemic wasting syndrome (PMWS) and porcine dermatitis and nephropathy syndrome (PDNS) affected pigs. Vet. Microbiol. 2011, 153, 377–381. [Google Scholar] [CrossRef]
- Blomström, A.-L.; Belák, S.; Fossum, C.; Fuxler, L.; Wallgren, P.; Berg, M. Studies of porcine circovirus type 2, porcine boca-like virus and torque teno virus indicate the presence of multiple viral infections in postweaning multisystemic wasting syndrome pigs. Virus Res. 2010, 152, 59–64. [Google Scholar] [CrossRef]
- Vlasakova, M.; Leskova, V.; Sliz, I.; Jackova, A.; Vilcek, S. The presence of six potentially pathogenic viruses in pigs suffering from post-weaning multisystemic wasting syndrome. BMC Vet. Res. 2014, 10, 221. [Google Scholar] [CrossRef]
- Blois, S.; Mallus, F.; Liciardi, M.; Pilo, C.; Camboni, T.; Macera, L.; Maggi, F.; Manzin, A. High Prevalence of Co-Infection with Multiple Torque Teno Sus Virus Species in Italian Pig Herds. PLoS ONE 2014, 9, e113720. [Google Scholar] [CrossRef]
- Kekarainen, T.; Sibila, M.; Segalés, J. Prevalence of swine Torque teno virus in post-weaning multisystemic wasting syndrome (PMWS)-affected and non-PMWS-affected pigs in Spain. J. Gen. Virol. 2006, 87, 833–837. [Google Scholar] [CrossRef]
- Ellis, J.A.; Allan, G.; Krakowka, S. Effect of coinfection with genogroup 1 porcine torque teno virus on porcine circovirus type 2–associated postweaning multisystemic wasting syndrome in gnotobiotic pigs. Am. J. Vet. Res. 2008, 69, 1608–1614. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.; Rakibuzzaman, A.G.M.; Ramamoorthy, S. Torque teno viruses in health and disease. Virus Res. 2020, 285, 198013. [Google Scholar] [CrossRef] [PubMed]
- Krakowka, S.; Ellis, J.A. Evaluation of the effects of porcine genogroup 1 torque teno virus in gnotobiotic swine. Am. J. Vet. Res. 2008, 69, 1623–1629. [Google Scholar] [CrossRef] [PubMed]
- Mei, M.; Zhu, L.; Wang, Y.; Xu, Z.; Zhao, L.; Peng, X.; Wu, Y.; Li, S.; Guo, W. Histopathological investigation in porcine infected with torque teno sus virus type 2 by inoculation. Virol. J. 2011, 8, 545. [Google Scholar] [CrossRef]
- Tshering, C.; Takagi, M.; Deguchi, E. Detection of Torque teno sus virus 1 and 2 in tissues from stillborn piglets delivered by sows via natural farrowing. J. Vet. Sci. 2012, 13, 425–427. [Google Scholar] [CrossRef]
- Teixeira, T.F.; Dezen, D.; Cibulski, S.P.; Varela, A.P.M.; Holz, C.L.; Franco, A.C.; Roehe, P.M. Torque Teno Sus Virus (TTSuV) in Cell Cultures and Trypsin. PLoS ONE 2011, 6, e17501. [Google Scholar] [CrossRef]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef]
- ICTV. Genus: Pestivirus. 2019. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/flaviviridae/361/genus-pestivirus (accessed on 19 November 2021).
- Postel, A.; Smith, D.B.; Becher, P. Proposed Update to the Taxonomy of Pestiviruses: Eight Additional Species within the Genus Pestivirus, Family Flaviviridae. Viruses 2021, 13, 1542. [Google Scholar] [CrossRef]
- Lamp, B.; Schwarz, L.; Högler, S.; Riedel, C.; Sinn, L.; Rebel-Bauder, B.; Weissenböck, H.; Ladinig, A.; Rümenapf, T. Novel Pestivirus Species in Pigs, Austria, 2015. Emerg. Infect. Dis. 2017, 23, 1176–1179. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Frost, M.J.; Finlaison, D.S.; King, K.R.; Ridpath, J.F.; Gu, X. Identification of a novel virus in pigs—Bungowannah virus: A possible new species of pestivirus. Virus Res. 2007, 129, 26–34. [Google Scholar] [CrossRef]
- Cagatay, G.N.; Antos, A.; Meyer, D.; Maistrelli, C.; Keuling, O.; Becher, P.; Postel, A. Frequent infection of wild boar with atypical porcine pestivirus (APPV). Transbound. Emerg. Dis. 2018, 65, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, C.; Stalder, H.; Sidler, X.; Renzullo, S.; Gurtner, C.; Grahofer, A.; Schweizer, M. Long-Term Circulation of Atypical Porcine Pestivirus (APPV) within Switzerland. Viruses 2019, 11, 653. [Google Scholar] [CrossRef] [PubMed]
- Kiesler, A.; Seitz, K.; Schwarz, L.; Buczolich, K.; Petznek, H.; Sassu, E.; Dürlinger, S.; Högler, S.; Klang, A.; Riedel, C.; et al. Clinical and Serological Evaluation of LINDA Virus Infections in Post-Weaning Piglets. Viruses 2019, 11, 975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiesler, A.; Plankensteiner, J.; Schwarz, L.; Riedel, C.; Seitz, K.; Mötz, M.; Ladinig, A.; Lamp, B.; Rümenapf, T. Prevalence of Linda Virus Neutralizing Antibodies in the Austrian Pig Population. Viruses 2021, 13, 1001. [Google Scholar] [CrossRef] [PubMed]
- Kiesler, A.; Schwarz, L.; Riedel, C.; Högler, S.; Brunthaler, R.; Dimmel, K.; Auer, A.; Zaruba, M.; Mötz, M.; Seitz, K.; et al. New Emergence of the Novel Pestivirus Linda Virus in a Pig Farm in Carinthia, Austria. Viruses 2022, 14, 326. [Google Scholar] [CrossRef] [PubMed]


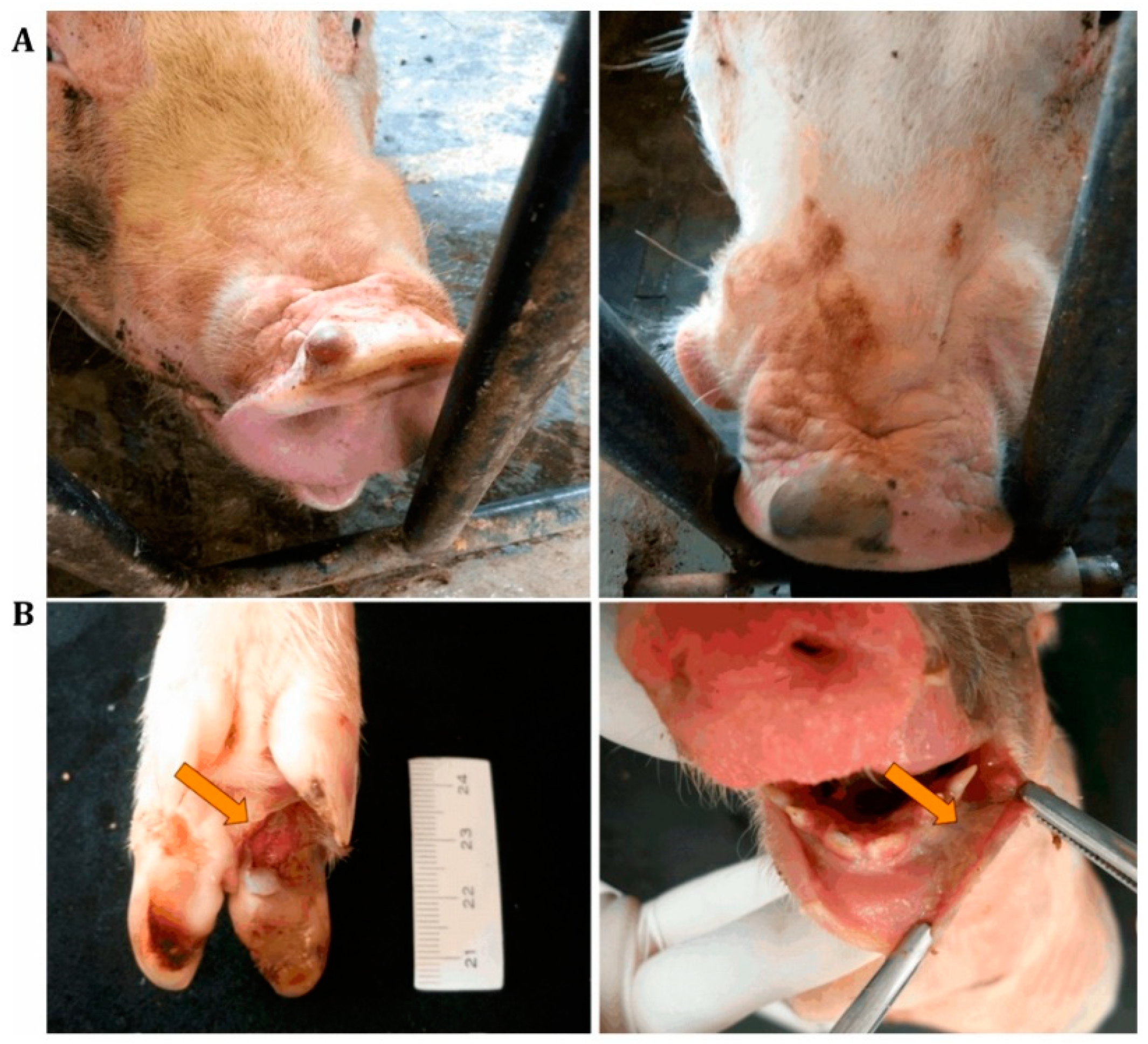{kind=link}
| Study | Gross Lesions | Histopathology | Diagnostic Method(s) |
|---|---|---|---|
| Plasma et al., 2008 [22] | Coalescing erosions or ruptured vesicle on the snout and coronary band. | NA * | PCR |
| Singh et al., 2012 [23] | Intact and ruptured vesicles and erosions in the oral cavity, around the nares and the coronary bands. Ulcers in fore and hind limbs. | Suppurative and ulcerative dermatitis | RT-PCR |
| Vannucci et al., 2015 [24] | Vesicles and coalescing erosions on the snouts and coronary bands of sows. Occasionally, vesicles and erosions on the coronary bands of piglets. | Hyperkeratosis, intra- and inter-cellular edema of keratinocytes and acantholytic degeneration of basal keratinocytes | Next-generation sequencing, RT-PCR |
| Baker et al., 2016 [27] | Nasal, coronary band, and hoof vesicular lesions in sows. No gross lesions in neonatal pigs. | NA | NA |
| Canning et al., 2016 [28] | Vesicular lesions on snouts and hoof lesions in sows. No gross lesions in neonatal pigs. | NA | RT-PCR |
| Wang et al., 2016 [29] | Ruptured vesicular lesions on the snout and coronary bands in sow | NA | RT-PCR |
| Wu et al., 2017 [31] * | Fluid-filled vesicles on the snout, interdigital cleft, coronary band. Petechial hemorrhages of kidney and interstitial pneumonia in piglets | Suppurative inflammation in dermis and epidermis, cell necrosis and damage of epithelial cells in hoof. Other lesions described are not strictly associated with SVA | RT-PCR |
| Montiel et al., 2016 [41] * | Intact or ruptured vesicular lesions on the coronary bands of toes and dewclaws or the interdigital space. Vesicular lesions and erosions on snouts. | NA | RT-PCR |
| Leme et al., 2016 [44] | Rib impressions on the pleural surface, pulmonary oedema, congestion of meningeal vessels, petechial hemorrhages of the kidney, diphtheritic glossitis, ulcerative lesions at the coronary band, multifocal cutaneous crusts | Interstitial pneumonia, lymphoid depletion, lymphocytic myocarditis, ballooning degeneration of transitional epithelium of the urinary bladder and ureters, lymphoplasmacytic encephalitis, vacuolation and atrophy of intestinal villi. | RT-PCR, IHC |
| Olivera et al., 2017 [45] | Erosive lesion at the coronary band, mesocolonic edema, multifocal ulceration of the skin | Ballooning degeneration of transitional epithelium, nonsuppurative meningoencephalitis, plexus choroiditis, atrophic enteritis | RT-PCR, IHC |
| Study | Gross Lesions | Histopathology | Diagnostic Method(s) |
|---|---|---|---|
| Krakowka and Ellis, 2008 * [97] | Interstitial pneumonia, mild thymic atrophy, edema in the ventral region of the neck and thoracic mediastinum | Interstitial pneumonia, membranous glomerulonephropathy, moderate lymphohistiocytic infiltrate in the liver. | PCR, nPCR, ISH |
| Mei et al., 2011 [98] * | Limited and still unclear | Hyperemia and congestion in the myocardium and endocardium, interstitial pneumonia, membranous glomerular nephropathy, and lymphocytic inflammation in the liver. | nPCR, ELISA |
| Polster et al., 2022 [86] | NA | Non-suppurative encephalitis/meningoencephalitis/plexus choroiditis. | RT-PCR, ISH, IHC |
| Study | Gross Lesions | Histopathology | Diagnostic Method(s) |
|---|---|---|---|
| Lamp et al., 2017 [104] | No specific gross lesions | Hypomyelination in the white matter of the spinal cord | RT-PCR, IHC |
| Kiesler et al., 2022 [110] | No specific gross lesions | Brain and spinal cord perivascular lymphoplasmacellular infiltrations | RT-PCR, IHC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hawko, S.; Burrai, G.P.; Polinas, M.; Angioi, P.P.; Dei Giudici, S.; Oggiano, A.; Alberti, A.; Hosri, C.; Antuofermo, E. A Review on Pathological and Diagnostic Aspects of Emerging Viruses—Senecavirus A, Torque teno sus virus and Linda Virus—In Swine. Vet. Sci. 2022, 9, 495. https://doi.org/10.3390/vetsci9090495
Hawko S, Burrai GP, Polinas M, Angioi PP, Dei Giudici S, Oggiano A, Alberti A, Hosri C, Antuofermo E. A Review on Pathological and Diagnostic Aspects of Emerging Viruses—Senecavirus A, Torque teno sus virus and Linda Virus—In Swine. Veterinary Sciences. 2022; 9(9):495. https://doi.org/10.3390/vetsci9090495
Chicago/Turabian StyleHawko, Salwa, Giovanni P. Burrai, Marta Polinas, Pier Paolo Angioi, Silvia Dei Giudici, Annalisa Oggiano, Alberto Alberti, Chadi Hosri, and Elisabetta Antuofermo. 2022. "A Review on Pathological and Diagnostic Aspects of Emerging Viruses—Senecavirus A, Torque teno sus virus and Linda Virus—In Swine" Veterinary Sciences 9, no. 9: 495. https://doi.org/10.3390/vetsci9090495