
Use of Phages to Treat Antimicrobial-Resistant Salmonella Infections in Poultry

Abstract
:Simple Summary
Abstract
1. Introduction
2. Salmonellosis and Its Association with Poultry
3. Antibiotic-Resistant Salmonella in Poultry
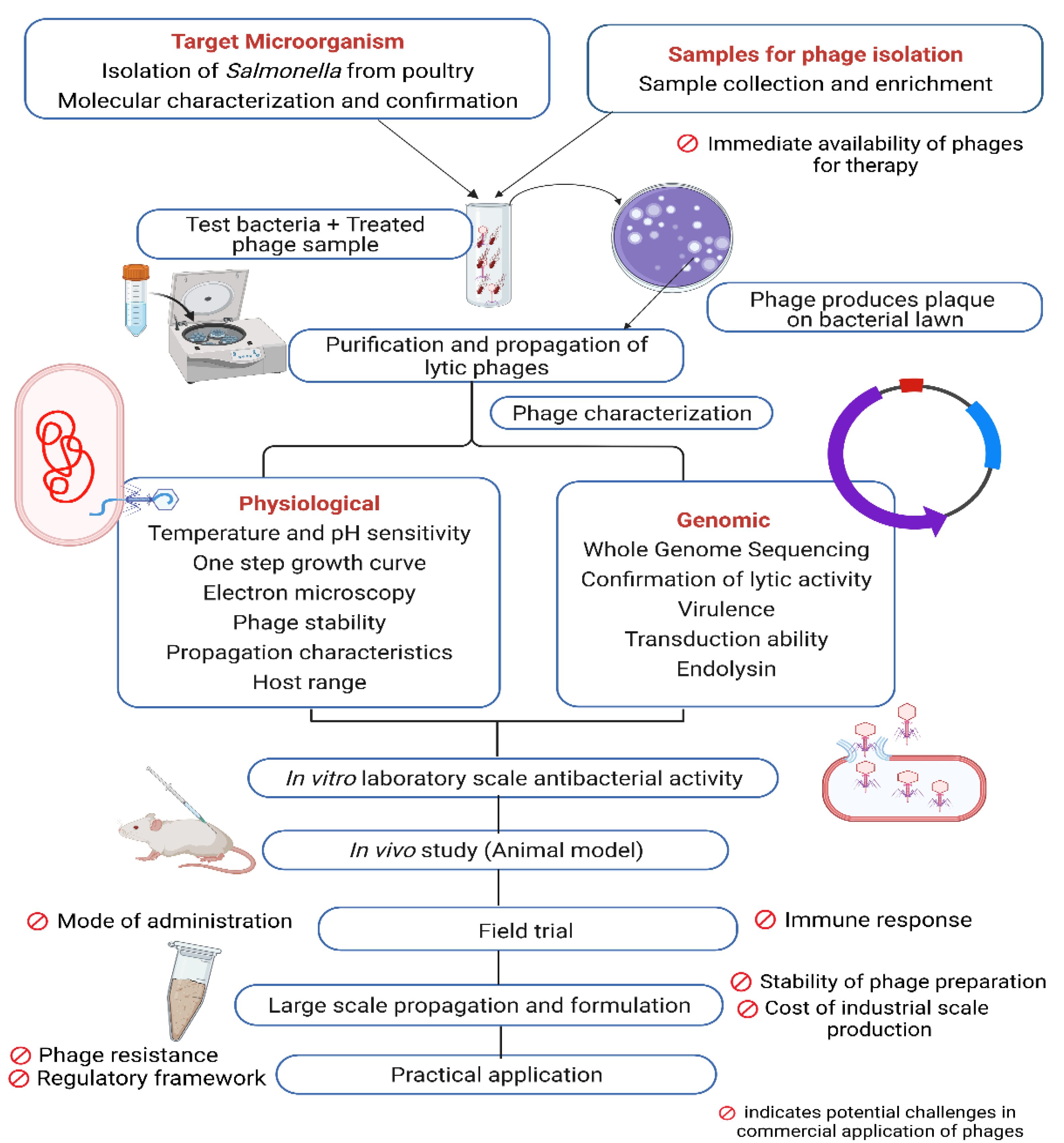
4. Bacteriophages an Alternative to Antibiotics in Controlling Pathogens
5. Application of Phages for Controlling Salmonella Infections in Poultry and Poultry Products
6. Challenges of Using Phages in Poultry and Probable Solutions
7. Future Prospect of Bacteriophage
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kurtz, J.R.; Goggins, J.A.; McLachlan, J.B. Salmonella infection: Interplay between the bacteria and host immune system. Immunol. Lett. 2017, 190, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Andino, A.; Hanning, I. Salmonella enterica: Survival, colonization, and virulence differences among serovars. Sci. World J. 2015, 2015, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef] [PubMed]
- Eng, S.-K.; Pusparajah, P.; Ab Mutalib, N.-S.; Ser, H.-L.; Chan, K.-G.; Lee, L.-H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef]
- Pui, C.F.; Wong, W.C.; Chai, L.C.; Tunung, R.; Jeyaletchumi, P.; Hidayah, N.; Ubong, A.; Farinazleen, M.G.; Cheah, Y.K.; Son, R. Salmonella: A foodborne pathogen. Int. Food Res. J. 2011, 18, 465–473. [Google Scholar]
- Angulo, F.J.; Mølbak, K. Human health consequences of antimicrobial drug-resistant salmonella and other foodborne pathogens. Clin. Infect. Dis. 2005, 41, 1613–1620. [Google Scholar] [CrossRef]
- Ehuwa, O.; Jaiswal, A.K.; Jaiswal, S. Salmonella, food safety and food handling practices. Foods 2021, 10, 907. [Google Scholar] [CrossRef]
- Hsi, D.J.; Ebel, E.D.; Williams, M.S.; Golden, N.J.; Schlosser, W.D. Comparing foodborne illness risks among meat commodities in the United States. Food Control 2015, 54, 353–359. [Google Scholar] [CrossRef]
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef]
- Medina, M.; Legido-Quigley, H.; Hsu, L.Y. Antimicrobial resistance in one health. In Global Health Security; Springer: Cham, Germany, 2020; pp. 209–229. [Google Scholar]
- Wallinga, D.; Smit, L.A.M.; Davis, M.F.; Casey, J.A.; Nachman, K.E. A Review of the Effectiveness of Current US Policies on Antimicrobial Use in Meat and Poultry Production. Curr. Environ. Health Rep. 2022, 9, 339–354. [Google Scholar] [CrossRef]
- Agyare, C.; Boamah, V.E.; Zumbi, C.N.; Osei, F.B. Antibiotic use in poultry production and its effects on bacterial resistance. In Antimicrobial Resistance—A Global Threat; IntechOpen: London, UK, 2018; pp. 1–20. [Google Scholar]
- Islam, S.; Urmi, U.L.; Rana, M.; Sultana, F.; Jahan, N.; Hossain, B.; Iqbal, S.; Hossain, M.; Mosaddek, A.S.; Nahar, S. High abundance of the colistin resistance gene mcr-1 in chicken gut-bacteria in Bangladesh. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kasman, L.M.; Porter, L.D. Bacteriophages. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Mahony, J.; McAuliffe, O.; Ross, R.P.; Van Sinderen, D. Bacteriophages as biocontrol agents of food pathogens. Curr. Opin. Biotechnol. 2011, 22, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Atterbury, R.J.; Gigante, A.M.; Rubio Lozano, M.D.L.S.; Méndez Medina, R.D.; Robinson, G.; Alloush, H.; Barrow, P.A.; Allen, V.M. Reduction of Salmonella contamination on the surface of chicken skin using bacteriophage. Virol. J. 2020, 17, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Virk, S.M.; Shi, J.; Zhou, Y.; Willias, S.P.; Morsy, M.K.; Abdelnabby, H.E.; Liu, J.; Wang, X.; Li, J. Isolation, characterization, and application of bacteriophage LPSE1 against Salmonella enterica in ready to eat (RTE) foods. Front. Microbiol. 2018, 9, 1046. [Google Scholar] [CrossRef]
- Hungaro, H.M.; Mendonça, R.C.S.; Gouvêa, D.M.; Vanetti, M.C.D.; de Oliveira Pinto, C.L. Use of bacteriophages to reduce Salmonella in chicken skin in comparison with chemical agents. Food Res. Int. 2013, 52, 75–81. [Google Scholar] [CrossRef]
- Sharma, M.; Patel, J.R.; Conway, W.S.; Ferguson, S.; Sulakvelidze, A. Effectiveness of bacteriophages in reducing Escherichia coli O157: H7 on fresh-cut cantaloupes and lettuce. J. Food Prot. 2009, 72, 1481–1485. [Google Scholar] [CrossRef]
- Atterbury, R.J.; Van Bergen, M.A.P.; Ortiz, F.; Lovell, M.A.; Harris, J.A.; De Boer, A.; Wagenaar, J.A.; Allen, V.M.; Barrow, P.A. Bacteriophage therapy to reduce Salmonella colonization of broiler chickens. Appl. Environ. Microbiol. 2007, 73, 4543–4549. [Google Scholar] [CrossRef]
- Li, J.; Zhao, F.; Zhan, W.; Li, Z.; Zou, L.; Zhao, Q. Challenges for the application of bacteriophages as effective antibacterial agents in the food industry. J. Sci. Food Agric. 2022, 102, 461–471. [Google Scholar] [CrossRef]
- Chen, H.-M.; Wang, Y.; Su, L.-H.; Chiu, C.-H. Nontyphoid Salmonella infection: Microbiology, clinical features, and antimicrobial therapy. Pediatr. Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef]
- Vought, K.J.; Tatini, S.R. Salmonella enteritidis contamination of ice cream associated with a 1994 multistate outbreak. J. Food Prot. 1998, 61, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Wong, D.M.A.L.F.; Patrick, M.E.; Binsztein, N.; Cieslik, A.; Chalermchaikit, T.; Aidara-Kane, A.; Ellis, A.; Angulo, F.J.; Wegener, H.C. Web-based surveillance and global Salmonella distribution, 2000–2002. Emerg. Infect. Dis. 2006, 12, 381. [Google Scholar] [CrossRef] [PubMed]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease ‘Burden of Illness’ Studies. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Authority, E.F.S. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. 2017, 15, 5077. [Google Scholar]
- United States Department of Health and Human services, CDC. Antibiotic Resistance Threats in the United States, 2019; Department of Health and Human Services: Atlanta, GA, USA, 2019. [Google Scholar]
- Mead, G.; Lammerding, A.M.; Cox, N.; Doyle, M.P.; Humbert, F.; Kulikovskiy, A.; Panin, A.; do Nascimento, V.P.; Wierup, M.; The Salmonella On Raw Poultry Writing Committee. Scientific and technical factors affecting the setting of Salmonella criteria for raw poultry: A global perspective. J. Food Prot. 2010, 73, 1566–1590. [Google Scholar] [CrossRef] [PubMed]
- Hugas, M.; Beloeil, P.A. Controlling Salmonella along the food chain in the European Union-progress over the last ten years. Eurosurveillance 2014, 19, 20804. [Google Scholar] [CrossRef] [PubMed]
- Guard-Petter, J. The chicken, the egg and Salmonella enteritidis. Environ. Microbiol. 2001, 3, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration, H.H.S. Prevention of Salmonella enteritidis in shell eggs during production, storage, and transportation. Final rule. Fed. Regist. 2009, 74, 33029–33101. [Google Scholar]
- De Reu, K.; Grijspeerdt, K.; Messens, W.; Heyndrickx, M.; Uyttendaele, M.; Debevere, J.; Herman, L. Eggshell factors influencing eggshell penetration and whole egg contamination by different bacteria, including Salmonella enteritidis. Int. J. Food Microbiol. 2006, 112, 253–260. [Google Scholar] [CrossRef]
- Messens, W.; Grijspeerdt, K.; Herman, L. Eggshell penetration by Salmonella: A review. World’s Poult. Sci. J. 2005, 61, 71–86. [Google Scholar] [CrossRef]
- Gantois, I.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F.; Gast, R.; Humphrey, T.J.; Van Immerseel, F. Mechanisms of egg contamination by Salmonella Enteritidis. FEMS Microbiol. Rev. 2009, 33, 718–738. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Yadav, A.S.; Singh, S.M.; Bharti, P. Prevalence of Salmonella in chicken eggs collected from poultry farms and marketing channels and their antimicrobial resistance. Food Res. Int. 2010, 43, 2027–2030. [Google Scholar] [CrossRef]
- Karim, M.R.; Giasuddin, M.; Samad, M.A.; Mahmud, M.S.; Islam, M.R.; Rahman, M.H.; Yousuf, M.A. Prevalence of Salmonella spp. in poultry and poultry products in Dhaka, Bangladesh. Int. J. Anim. Biol 2017, 3, 18–22. [Google Scholar]
- Mahmud, M.S.; Kabir, M.L.; Alam, S.M.S.; Ali, M.M.; Towhid, S.T. Prevalence of Salmonella spp. in poultry eggs from different retail markets at Savar area, Bangladesh. Am. J. Food Sci. Health 2015, 1, 27–31. [Google Scholar]
- Taddese, D.; Tolosa, T.; Deresa, B.; Lakow, M.; Olani, A.; Shumi, E. Antibiograms and risk factors of Salmonella isolates from laying hens and eggs in Jimma Town, South Western Ethiopia. BMC Res. Notes 2019, 12, 1–7. [Google Scholar] [CrossRef]
- Kemal, J.; Sibhat, B.; Menkir, S.; Beyene, D. Prevalence, assessment, and antimicrobial resistance patterns of Salmonella from raw chicken eggs in Haramaya, Ethiopia. J. Infect. Dev. Ctries. 2016, 10, 1230–1235. [Google Scholar] [CrossRef]
- Li, W.; Li, H.; Zheng, S.; Wang, Z.; Sheng, H.; Shi, C.; Shi, X.; Niu, Q.; Yang, B. Prevalence, serotype, antibiotic susceptibility, and genotype of Salmonella in eggs from poultry farms and marketplaces in Yangling, Shaanxi province, China. Front. Microbiol. 2020, 11, 1482. [Google Scholar] [CrossRef]
- Xie, T.; Wu, G.; He, X.; Lai, Z.; Zhang, H.; Zhao, J. Antimicrobial resistance and genetic diversity of Salmonella enterica from eggs. Food Sci. Nutr. 2019, 7, 2847–2853. [Google Scholar] [CrossRef]
- Hosseininezhad, B.; Berizi, E.; Nader, M.; Mazloomi, S.M.; Hosseinzadeh, S.; Ebrahimi, L.; Zare, M. Prevalence of Salmonella contamination in consumed eggs in Iran: A systematic review and meta-analysis study on published studies from 1996 to 2018. Vet. World 2020, 13, 2743–2751. [Google Scholar] [CrossRef]
- Shahzad, A.; Mahmood, M.S.; Hussain, I.; Siddique, F.; Abbas, R.Z. Prevalence of salmonella species in hen eggs and egg storing-trays collected from poultry farms and marketing outlets of Faisalabad, Pakistan. Pak. J. Agric. Sci. 2012, 49, 565–568. [Google Scholar]
- Al-Zenki, S.; Al-Nasser, A.; Al-Safar, A.; Alomirah, H.; Al-Haddad, A.; Hendriksen, R.S.; Aarestrup, F.M. Prevalence and antibiotic resistance of Salmonella isolated from a poultry farm and processing plant environment in the State of Kuwait. Foodborne Pathog. Dis. 2007, 4, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Zubair, A.I.; Al-Berfkani, M.I.; Issa, A.R. Prevalence of Salmonella species from poultry eggs of local stores in Duhok. Int. J. Res. Med. Sci. 2017, 5, 2468–2471. [Google Scholar] [CrossRef]
- Saitanu, K.; Jerngklinchan, J.; Koowatananukul, C. Incidence of salmonellae in duck eggs in Thailand. Southeast Asian J. Trop. Med. Public Health 1994, 25, 328. [Google Scholar] [PubMed]
- Musawa, A.I.; Bashiru, G.; Al-Rasheed, A.; Yakubu, Y.; Jibril, A.H.; Ballah, F.M.; Sidi, S.; Lawal, N.; Bala, J.A.; Odhah, M.N.; et al. Prevalence and antimicrobial susceptibility profiling of salmonella isolated from poultry products sold in sokoto metropolis, nigeria. J. Anim. Health Prod. 2021, 9, 148–155. [Google Scholar]
- Haubert, L.; Maia, D.S.V.; Rauber Würfel, S.D.F.; Vaniel, C.; da Silva, W.P. Virulence genes and sanitizers resistance in Salmonella isolates from eggs in southern Brazil. J. Food Sci. Technol. 2021, 59, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Sharuni, A.A.G.U.; Sumaiya, M.R.F.; Ekanayake, E.M.D.N.; Chandrasiri, N.S.; Arachchi, A.M.P.S. Contamination of chicken eggshell and egg contents with Salmonella species from selected farms in Kosgama, Colombo district. Sri Lankan J. Infect. Dis. 2021, 11, S2–S6. [Google Scholar] [CrossRef]
- Kapena, M.S.; Muma, J.B.; Mubita, C.M.; Munyeme, M. Antimicrobial resistance of Escherichia coli and Salmonella in raw retail table eggs in Lusaka, Zambia. Vet. World 2020, 13, 2528. [Google Scholar] [CrossRef]
- Betancor, L.; Pereira, M.; Martinez, A.; Giossa, G.; Fookes, M.; Flores, K.; Barrios, P.; Repiso, V.; Vignoli, R.; Cordeiro, N.; et al. Prevalence of Salmonella enterica in poultry and eggs in Uruguay during an epidemic due to Salmonella enterica serovar Enteritidis. J. Clin. Microbiol. 2010, 48, 2413–2423. [Google Scholar] [CrossRef]
- Sasaki, Y.; Tsujiyama, Y.; Asai, T.; Noda, Y.; Katayama, S.; Yamada, Y. Salmonella prevalence in commercial raw shell eggs in Japan: A survey. Epidemiol. Infect. 2011, 139, 1060–1064. [Google Scholar] [CrossRef]
- Lee, S.-K.; Chon, J.-W.; Song, K.-Y.; Hyeon, J.-Y.; Moon, J.-S.; Seo, K.-H. Prevalence, characterization, and antimicrobial susceptibility of Salmonella Gallinarum isolated from eggs produced in conventional or organic farms in South Korea. Poult. Sci. 2013, 92, 2789–2797. [Google Scholar] [CrossRef]
- Vergara, D.C.M.; Gutiérrez, V.E.R.; García, N.V. Prevalence and molecular identification of Salmonella spp. isolated from commercialized eggs at Ibague, Colombia. Rev. Salud Anim. 2016, 38, 164–172. [Google Scholar]
- Kouam, M.K.; Biekop, M.H.F.; Katte, B.; Teguia, A. Salmonella status of table eggs in commercial layer farms in Menoua Division, West region of Cameroon. Food Control 2018, 85, 345–349. [Google Scholar] [CrossRef]
- Murchie, L.; Whyte, P.; Xia, B.; Horrigan, S.; Kelly, L.; Madden, R.H. Prevalence of Salmonella in grade A whole shell eggs in the island of Ireland. J. Food Prot. 2007, 70, 1238–1240. [Google Scholar] [CrossRef]
- Liljebjelke, K.A.; Hofacre, C.L.; Liu, T.; White, D.G.; Ayers, S.; Young, S.; Maurer, J.J. Vertical and horizontal transmission of Salmonella within integrated broiler production system. Foodb. Pathog. Dis. 2005, 2, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Van de Giessen, A.W.; Bouwknegt, M.; Dam-Deisz, W.D.C.; van Pelt, W.; Wannet, W.J.B.; Visser, G. Surveillance of Salmonella spp. and Campylobacter spp. in poultry production flocks in The Netherlands. Epidemiol. Infect. 2006, 134, 1266–1275. [Google Scholar] [CrossRef]
- Gutierrez, M.; Fanning, J.; Murphy, A.; Murray, G.; Griffin, M.; Flack, A.; Leonard, N.; Egan, J. Salmonella in broiler flocks in the republic of Ireland. Foodborne Pathog. Dis. 2009, 6, 111–120. [Google Scholar] [CrossRef]
- Srinivasan, P.; Balasubramaniam, G.A.; Gopala, T.R.; Murthy, K.; Saravanan, S.; Balachandran, P. Prevalence and Pathology of Salmonellosis in Commercial Layer Chicken from Namakkal, India. Pak. Vet. J. 2014, 34, 3241–3328. [Google Scholar]
- Duc, V.M.; Nakamoto, Y.; Fujiwara, A.; Toyofuku, H.; Obi, T.; Chuma, T. Prevalence of Salmonella in broiler chickens in Kagoshima, Japan in 2009 to 2012 and the relationship between serovars changing and antimicrobial resistance. BMC Vet. Res. 2019, 15, 1–8. [Google Scholar] [CrossRef]
- Zhao, X.; Gao, Y.; Ye, C.; Yang, L.; Wang, T.; Chang, W. Prevalence and characteristics of Salmonella isolated from free-range chickens in Shandong Province, China. Biomed Res. Int. 2016, 2016, 1–6. [Google Scholar]
- Roe, M.T.; Pillai, S.D. Monitoring and identifying antibiotic resistance mechanisms in bacteria. Poult. Sci. 2003, 82, 622–626. [Google Scholar] [CrossRef]
- Dhanarani, T.S.; Shankar, C.; Park, J.; Dexilin, M.; Kumar, R.R.; Thamaraiselvi, K. Study on acquisition of bacterial antibiotic resistance determinants in poultry litter. Poult. Sci. 2009, 88, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Castro-Vargas, R.E.; Herrera-Sánchez, M.P.; Rodríguez-Hernández, R.; Rondón-Barragán, I.S. Antibiotic resistance in Salmonella spp. isolated from poultry: A global overview. Vet. World 2020, 13, 2070. [Google Scholar] [CrossRef] [PubMed]
- Harsha, H.T.; Reshmi, R.; Varghese, R.; Divya, P.S.; Rahiman, K.M.M.; Hatha, A.A.M. Prevalence and antibiotic resistance of Salmonella from the eggs of commercial samples. J. Microbiol. Infect. Dis. 2011, 1, 93–100. [Google Scholar] [CrossRef]
- Mahmud, T.; Hassan, M.M.; Alam, M.; Khan, M.M.; Bari, M.S.; Islam, A. Prevalence and multidrug-resistant pattern of Salmonella from the eggs and egg-storing trays of retail markets of Bangladesh. Int. J. One Health 2016, 2, 7–11. [Google Scholar] [CrossRef]
- Hoffmann, M.; Pettengill, J.B.; Gonzalez-Escalona, N.; Miller, J.; Ayers, S.L.; Zhao, S.; Allard, M.W.; McDermott, P.F.; Brown, E.W.; Monday, S.R. Comparative sequence analysis of multidrug-resistant IncA/C plasmids from Salmonella enterica. Front. Microbiol. 2017, 8, 1459. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.B.; Mahmud, M.; Akter, R.; Hasan, M.; Sobur, A.; Nazir, K.H.M.; Noreddin, A.; Rahman, T.; El Zowalaty, M.E.; Rahman, M. Molecular detection of multidrug resistant Salmonella species isolated from broiler farm in Bangladesh. Pathogens 2020, 9, 201. [Google Scholar] [CrossRef]
- Uddin, M.B.; Hossain, S.M.; Hasan, M.; Alam, M.N.; Debnath, M.; Begum, R.; Roy, S.; Harun-Al-Rashid, A.; Chowdhury, M.S.R.; Rahman, M.M.; et al. Multidrug antimicrobial resistance and molecular detection of MCR-1 gene in Salmonella species isolated from chicken. Animals 2021, 11, 206. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, X.; Jiang, Z.; Qi, Y.; Ed-Dra, A.; Yue, M. Epidemiological investigation and antimicrobial resistance profiles of Salmonella isolated from breeder chicken hatcheries in Henan, China. Front. Cell. Infect. Microbiol. 2020, 10, 497. [Google Scholar] [CrossRef]
- Elkenany, R.; Elsayed, M.M.; Zakaria, A.I.; El-Sayed, S.A.-E.-S.; Rizk, M.A. Antimicrobial resistance profiles and virulence genotyping of Salmonella enterica serovars recovered from broiler chickens and chicken carcasses in Egypt. BMC Vet. Res. 2019, 15, 1–9. [Google Scholar] [CrossRef]
- Ghaderi, R.; Moradi Bidhendi, S.; Khaki, P. Occurrence of multidrug-resistant Salmonella enterica serovar Enteritidis isolates from poultry in Iran. Arch. Razi Inst. 2016, 71, 43–49. [Google Scholar]
- Wajid, M.; Saleemi, M.K.; Sarwar, Y.; Ali, A. Detection and characterization of multidrug-resistant Salmonella enterica serovar Infantis as an emerging threat in poultry farms of Faisalabad, Pakistan. J. Appl. Microbiol. 2019, 127, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhao, H.; Sun, J.; Liu, Y.; Zhou, X.; Beier, R.C.; Wu, G.; Hou, X. Characterization of multidrug-resistant Salmonella enterica serovars Indiana and Enteritidis from chickens in Eastern China. PLoS ONE 2014, 9, e96050. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.; Kim, J.H.; Park, J.H.; Lee, Y.-J.; Lee, J.H. Molecular and virulence characteristics of multi-drug resistant Salmonella Enteritidis strains isolated from poultry. Vet. J. 2011, 189, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Yadav, A.S.; Tripathi, V.; Singh, R.P. Antimicrobial resistance profile of Salmonella present in poultry and poultry environment in north India. Food Control 2013, 33, 545–548. [Google Scholar] [CrossRef]
- Tirziu, E.; Lazăr, R.; Sala, C.; Nichita, I.; Morar, A.; Şereş, M.; Imre, K. Salmonella in raw chicken meat from the Romanian seaside: Frequency of isolation and antibiotic resistance. J. Food Prot. 2015, 78, 1003–1006. [Google Scholar] [CrossRef]
- Mokgophi, T.M.; Gcebe, N.; Fasina, F.; Adesiyun, A.A. Antimicrobial resistance profiles of Salmonella isolates on chickens processed and retailed at outlets of the informal market in Gauteng Province, South Africa. Pathogens 2021, 10, 273. [Google Scholar] [CrossRef]
- Ibrahim, S.; Wei Hoong, L.; Lai Siong, Y.; Mustapha, Z.; CW Zalati, C.W.; Aklilu, E.; Mohamad, M.; Kamaruzzaman, N.F. Prevalence of antimicrobial resistance (AMR) Salmonella spp. and Escherichia coli isolated from broilers in the East Coast of Peninsular Malaysia. Antibiotics 2021, 10, 579. [Google Scholar] [CrossRef]
- Vidayanti, I.N.; Sukon, P.; Khaengair, S.; Pulsrikarn, C.; Angkittitrakul, S. Prevalence and antimicrobial resistance of Salmonella spp. isolated from chicken meat in upper northeastern Thailand. Vet. Integr. Sci. 2021, 19, 121–131. [Google Scholar] [CrossRef]
- Zdragas, A.; Mazaraki, K.; Vafeas, G.; Giantzi, V.; Papadopoulos, T.; Ekateriniadou, L. Prevalence, seasonal occurrence and antimicrobial resistance of Salmonella in poultry retail products in Greece. Lett. Appl. Microbiol. 2012, 55, 308–313. [Google Scholar] [CrossRef]
- Ta, Y.T.; Nguyen, T.T.; To, P.B.; Pham, D.X.; Le, H.T.H.; Thi, G.N.; Alali, W.Q.; Walls, I.; Doyle, M.P. Quantification, serovars, and antibiotic resistance of Salmonella isolated from retail raw chicken meat in Vietnam. J. Food Prot. 2014, 77, 57–66. [Google Scholar] [CrossRef]
- Zwe, Y.H.; Tang, V.C.Y.; Aung, K.T.; Gutiérrez, R.A.; Ng, L.C.; Yuk, H.-G. Prevalence, sequence types, antibiotic resistance and, gyrA mutations of Salmonella isolated from retail fresh chicken meat in Singapore. Food Control 2018, 90, 233–240. [Google Scholar] [CrossRef]
- Donado-Godoy, P.; Clavijo, V.; León, M.; Arevalo, A.; Castellanos, R.; Bernal, J.; Tafur, M.A.; Ovalle, M.V.; Alali, W.Q.; Hume, M.; et al. Counts, serovars, and antimicrobial resistance phenotypes of Salmonella on raw chicken meat at retail in Colombia. J. Food Prot. 2014, 77, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Siriken, B.; Türk, H.; Yildirim, T.; Durupinar, B.; Erol, I. Prevalence and characterization of Salmonella isolated from chicken meat in Turkey. J. Food Sci. 2015, 80, M1044–M1050. [Google Scholar] [CrossRef]
- Moe, A.Z.; Paulsen, P.; Pichpol, D.; Fries, R.; Irsigler, H.; Baumann, M.P.O.; Oo, K.N. Prevalence and antimicrobial resistance of Salmonella isolates from chicken carcasses in retail markets in Yangon, Myanmar. J. Food Prot. 2017, 80, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Checkley, S.; Avery, B.; Chalmers, G.; Bohaychuk, V.; Gensler, G.; Reid-Smith, R.; Boerlin, P. Phenotypic and genetic characterization of antimicrobial resistance in Salmonella serovars isolated from retail meats in Alberta, Canada. Food Microbiol. 2012, 32, 110–117. [Google Scholar] [CrossRef]
- Procura, F.; Bueno, D.J.; Bruno, S.B.; Rogé, A.D. Prevalence, antimicrobial resistance profile and comparison of methods for the isolation of Salmonella in chicken liver from Argentina. Food Res. Int. 2019, 119, 541–546. [Google Scholar] [CrossRef]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G., Jr. Bacteriophage therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef]
- Hendrix, R.W.; Hatfull, G.F.; Ford, M.E.; Smith, M.C.M.; Burns, R.N. Evolutionary relationships among diverse bacteriophages and prophages: All the world’s a phage. Proc. Natl. Acad. Sci. USA 1999, 96, 2192–2197. [Google Scholar] [CrossRef]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage treatment of human infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef]
- Ackermann, H.-W. Phage classification and characterization. In Bacteriophages; Springer: Cham, Switzerland, 2009; pp. 127–140. [Google Scholar]
- Wittebole, X.; De Roock, S.; Opal, S.M. A historical overview of bacteriophage therapy as an alternative to antibiotics for the treatment of bacterial pathogens. Virulence 2014, 5, 226–235. [Google Scholar] [CrossRef]
- Azeredo, J.; Sutherland, I.W. The use of phages for the removal of infectious biofilms. Curr. Pharm. Biotechnol. 2008, 9, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Dvořáčková, M.; Růžička, F.; Benešík, M.; Pantůček, R.; Dvořáková-Heroldová, M. Antimicrobial effect of commercial phage preparation Stafal® on biofilm and planktonic forms of methicillin-resistant Staphylococcus aureus. Folia Microbiol. 2019, 64, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Pallavali, R.R.; Degati, V.L.; Lomada, D.; Reddy, M.C.; Durbaka, V.R.P. Isolation and in vitro evaluation of bacteriophages against MDR-bacterial isolates from septic wound infections. PLoS ONE 2017, 12, e0179245. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.-H.; Wang, J.-L.; Wen, F.-S.; Chang, K.-M.; Kuo, C.-F.; Lin, C.-H.; Luo, H.-R.; Hung, C.-H. Isolation and characterization of φkm18p, a novel lytic phage with therapeutic potential against extensively drug resistant Acinetobacter baumannii. PLoS ONE 2012, 7, e46537. [Google Scholar] [CrossRef] [PubMed]
- Wall, S.K.; Zhang, J.; Rostagno, M.H.; Ebner, P.D. Phage therapy to reduce preprocessing Salmonella infections in market-weight swine. Appl. Environ. Microbiol. 2010, 76, 48–53. [Google Scholar] [CrossRef]
- Ooi, M.L.; Drilling, A.J.; Morales, S.; Fong, S.; Moraitis, S.; Macias-Valle, L.; Vreugde, S.; Psaltis, A.J.; Wormald, P.-J. Safety and tolerability of bacteriophage therapy for chronic rhinosinusitis due to Staphylococcus aureus. JAMA Otolaryngol. Neck Surg. 2019, 145, 723–729. [Google Scholar] [CrossRef]
- Letkiewicz, S.; Międzybrodzki, R.; Fortuna, W.; Weber-Dąbrowska, B.; Górski, A. Eradication of Enterococcus faecalis by phage therapy in chronic bacterial prostatitis—Case report. Folia Microbiol. 2009, 54, 457–461. [Google Scholar] [CrossRef]
- Jennes, S.; Merabishvili, M.; Soentjens, P.; Pang, K.W.; Rose, T.; Keersebilck, E.; Soete, O.; François, P.-M.; Teodorescu, S.; Verween, G.; et al. Use of bacteriophages in the treatment of colistin-only-sensitive Pseudomonas aeruginosa septicaemia in a patient with acute kidney injury—A case report. Crit. Care 2017, 21, 1–3. [Google Scholar] [CrossRef]
- Fadlallah, A.; Chelala, E.; Legeais, J.-M. Corneal infection therapy with topical bacteriophage administration. Open Ophthalmol. J. 2015, 9, 167–168. [Google Scholar] [CrossRef]
- Markoishvili, K.; Tsitlanadze, G.; Katsarava, R.; Glenn, J.; Morris, M.D., Jr.; Sulakvelidze, A. A novel sustained-release matrix based on biodegradable poly (ester amide) s and impregnated with bacteriophages and an antibiotic shows promise in management of infected venous stasis ulcers and other poorly healing wounds. Int. J. Dermatol. 2002, 41, 453–458. [Google Scholar] [CrossRef]
- Perera, M.N.; Abuladze, T.; Li, M.; Woolston, J.; Sulakvelidze, A. Bacteriophage cocktail significantly reduces or eliminates Listeria monocytogenes contamination on lettuce, apples, cheese, smoked salmon and frozen foods. Food Microbiol. 2015, 52, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Costa, A.R.; Pinto, G.; Meneses, L.; Azeredo, J. Current challenges and future opportunities of phage therapy. FEMS Microbiol. Rev. 2020, 44, 684–700. [Google Scholar] [CrossRef] [PubMed]
- Wernicki, A.; Nowaczek, A.; Urban-Chmiel, R. Bacteriophage therapy to combat bacterial infections in poultry. Virol. J. 2017, 14, 1–13. [Google Scholar] [CrossRef]
- Henriques, A.; Sereno, R.; Almeida, A. Reducing Salmonella horizontal transmission during egg incubation by phage therapy. Foodborne Pathog. Dis. 2013, 10, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Sonalika, J.; Srujana, A.S.; Akhila, D.S.; Juliet, M.R.; Santhosh, K.S. Application of bacteriophages to control Salmonella Enteritidis in raw eggs. Iran. J. Vet. Res. 2020, 21, 221–225. [Google Scholar]
- Carvalho, C.M.; Santos, S.B.; Kropinski, A.M.; Ferreira, E.C.; Azeredo, J. Phages as therapeutic tools to control major foodborne pathogens: Campylobacter and Salmonella. In Bacteriophages; InTech: Rijeka, Croatia, 2012; pp. 179–214. [Google Scholar]
- Bardina, C.; Spricigo, D.A.; Cortés, P.; Llagostera, M. Significance of the bacteriophage treatment schedule in reducing Salmonella colonization of poultry. Appl. Environ. Microbiol. 2012, 78, 6600–6607. [Google Scholar] [CrossRef]
- Fiorentin, L.; Vieira, N.D.; Barioni, W., Jr. Oral treatment with bacteriophages reduces the concentration of Salmonella Enteritidis PT4 in caecal contents of broilers. Avian Pathol. 2005, 34, 258–263. [Google Scholar] [CrossRef]
- Bielke, L.R.; Higgins, S.E.; Donoghue, A.M.; Donoghue, D.J.; Hargis, B.M.; Tellez, G. Use of wide-host-range bacteriophages to reduce Salmonella on poultry products. Int. J. Poult. Sci. 2007, 6, 754–757. [Google Scholar]
- Nabil, N.M.; Tawakol, M.M.; Hassan, H.M. Assessing the impact of bacteriophages in the treatment of Salmonella in broiler chickens. Infect. Ecol. Epidemiol. 2018, 8, 1539056. [Google Scholar] [CrossRef]
- Higgins, S.E.; Higgins, J.P.; Bielke, L.R.; Hargis, B.M. Selection and application of bacteriophages for treating Salmonella enteritidis infections in poultry. Int. J. Poult. Sci. 2007, 6, 163–168. [Google Scholar] [CrossRef]
- Lim, T.-H.; Kim, M.-S.; Lee, D.-H.; Lee, Y.-N.; Park, J.-K.; Youn, H.-N.; Lee, H.-J.; Yang, S.-Y.; Cho, Y.-W.; Lee, J.-B.; et al. Use of bacteriophage for biological control of Salmonella Enteritidis infection in chicken. Res. Vet. Sci. 2012, 93, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Borie, C.; Sánchez, M.L.; Navarro, C.; Ramírez, S.; Morales, M.A.; Retamales, J.; Robeson, J. Aerosol spray treatment with bacteriophages and competitive exclusion reduces Salmonella enteritidis infection in chickens. Avian Dis. 2009, 53, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.-H.; Lee, D.-H.; Lee, Y.-N.; Park, J.-K.; Youn, H.-N.; Kim, M.-S.; Lee, H.-J.; Yang, S.-Y.; Cho, Y.-W.; Lee, J.-B.; et al. Efficacy of bacteriophage therapy on horizontal transmission of Salmonella Gallinarum on commercial layer chickens. Avian Dis. 2011, 55, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Kimminau, E.A.; Russo, K.N.; Karnezos, T.P.; Oh, H.G.; Lee, J.J.; Tate, C.C.; Baxter, J.A.; Berghaus, R.D.; Hofacre, C.L. Bacteriophage in-feed application: A novel approach to preventing Salmonella Enteritidis colonization in chicks fed experimentally contaminated feed. J. Appl. Poult. Res. 2020, 29, 930–936. [Google Scholar] [CrossRef]
- SKLAR, I.A.N.B.; JOERGER, R.D. Attempts to utilize bacteriophage to combat Salmonella Enterica serovar entemtidis infection in chickens. J. Food Saf. 2001, 21, 15–29. [Google Scholar] [CrossRef]
- Borie, C.; Albala, I.; Sánchez, P.; Sánchez, M.L.; Ramírez, S.; Navarro, C.; Morales, M.A.; Retamales, J.; Robeson, J. Bacteriophage treatment reduces Salmonella colonization of infected chickens. Avian Dis. 2008, 52, 64–67. [Google Scholar] [CrossRef]
- Wong, C.L.; Sieo, C.C.; Tan, W.S.; Abdullah, N.; Hair-Bejo, M.; Abu, J.; Ho, Y.W. Evaluation of a lytic bacteriophage, Φ st1, for biocontrol of Salmonella enterica serovar Typhimurium in chickens. Int. J. Food Microbiol. 2014, 172, 92–101. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, Y.; Li, W.; Zhu, W.; Wang, J.; Wang, X. Application of a novel lytic podoviridae phage Pu20 for biological control of drug-resistant Salmonella in liquid eggs. Pathogens 2021, 10, 34. [Google Scholar] [CrossRef]
- Yi, Y.; Abdelhamid, A.G.; Xu, Y.; Yousef, A.E. Characterization of broad-host lytic Salmonella phages isolated from livestock farms and application against Salmonella Enteritidis in liquid whole egg. LWT 2021, 144, 111269. [Google Scholar] [CrossRef]
- Spricigo, D.A.; Bardina, C.; Cortés, P.; Llagostera, M. Use of a bacteriophage cocktail to control Salmonella in food and the food industry. Int. J. Food Microbiol. 2013, 165, 169–174. [Google Scholar] [CrossRef]
- Duc, H.M.; Son, H.M.; Honjoh, K.; Miyamoto, T. Isolation and application of bacteriophages to reduce Salmonella contamination in raw chicken meat. LWT 2018, 91, 353–360. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, P.; Zhang, H.; Zhou, Y.; Zhang, L.; Wang, R. Bio-control of Salmonella Enteritidis in foods using bacteriophages. Viruses 2015, 7, 4836–4853. [Google Scholar] [CrossRef] [PubMed]
- Esmael, A.; Azab, E.; Gobouri, A.A.; Nasr-Eldin, M.A.; Moustafa, M.; Mohamed, S.A.; Badr, O.A.; Abdelatty, A.M. Isolation and characterization of two lytic bacteriophages infecting a multi-drug resistant Salmonella Typhimurium and their efficacy to combat salmonellosis in ready-to-use foods. Microorganisms 2021, 9, 423. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, H.J.; Jung, S.J.; Mizan, M.F.R.; Park, S.H.; Ha, S. Characterization of Salmonella spp.-specific bacteriophages and their biocontrol application in chicken breast meat. J. Food Sci. 2020, 85, 526–534. [Google Scholar] [CrossRef]
- Sukumaran, A.T.; Nannapaneni, R.; Kiess, A.; Sharma, C.S. Reduction of Salmonella on chicken meat and chicken skin by combined or sequential application of lytic bacteriophage with chemical antimicrobials. Int. J. Food Microbiol. 2015, 207, 8–15. [Google Scholar] [CrossRef]
- Shakeri, G.; Hammerl, J.A.; Jamshidi, A.; Ghazvini, K.; Rohde, M.; Szabo, I.; Kehrenberg, C.; Plötz, M.; Kittler, S. The Lytic Siphophage vB_StyS-LmqsSP1 Reduces the Number of Salmonella enterica Serovar Typhimurium Isolates on Chicken Skin. Appl. Environ. Microbiol. 2021, 87, e0142421. [Google Scholar] [CrossRef]
- Demirarslan, Ö.A.; Alasalvar, H.; Yildirim, Z. Biocontrol of Salmonella Enteritidis on chicken meat and skin using lytic SE-P3, P16, P37, and P47 bacteriophages. LWT 2021, 137, 110469. [Google Scholar] [CrossRef]
- Aguilera, M.; Martínez, S.; Tello, M.; Gallardo, M.J.; García, V. Use of Cocktail of Bacteriophage for Salmonella Typhimurium Control in Chicken Meat. Foods 2022, 11, 1164. [Google Scholar] [CrossRef]
- Kumar, A.; Malik, H.; Dubal, Z.B.; Jaiswal, R.K.; Kumar, S.; Kumar, B.; Agarwal, R.K. Isolation and characterization of Salmonella phages and phage cocktail mediated biocontrol of Salmonella enterica serovar Typhimurium in chicken meat. LWT 2022, 155, 112957. [Google Scholar]
- Rivera, D.; Moreno-Switt, A.I.; Denes, T.G.; Hudson, L.K.; Peters, T.L.; Samir, R.; Aziz, R.K.; Noben, J.-P.; Wagemans, J.; Dueñas, F. Novel Salmonella Phage, vB_Sen_STGO-35-1, Characterization and Evaluation in Chicken Meat. Microorganisms 2022, 10, 606. [Google Scholar] [CrossRef]
- Whichard, J.M.; Sriranganathan, N.; Pierson, F.W. Suppression of Salmonella growth by wild-type and large-plaque variants of bacteriophage Felix O1 in liquid culture and on chicken frankfurters. J. Food Prot. 2003, 66, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, Q.; Zhang, C.; Yang, J.; Lu, Z.; Lu, F.; Bie, X. Characterization of a broad host-spectrum virulent Salmonella bacteriophage fmb-p1 and its application on duck meat. Virus Res. 2017, 236, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, V.; Baquero, D.; Hernandez, S.; Farfan, J.C.; Arias, J.; Arévalo, A.; Donado-Godoy, P.; Vives-Flores, M. Phage cocktail SalmoFREE® reduces Salmonella on a commercial broiler farm. Poult. Sci. 2019, 98, 5054–5063. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, V.; Morales, L.T.; Flores, M.J.V.; Reyes, A. The gut microbiota of chickens in a commercial farm treated with a Salmonella phage cocktail. Sci. Rep. 2022, 12, 1–16. [Google Scholar]
- Thanki, A.M.; Hooton, S.; Gigante, A.M.; Atterbury, R.J.; Clokie, M.R.J. Potential Roles for Bacteriophages in Reducing Salmonella from Poultry and Swine; IntechOpen Limited: London, UK, 2021. [Google Scholar]
- Caflisch, K.M.; Suh, G.A.; Patel, R. Biological challenges of phage therapy and proposed solutions: A literature review. Expert Rev. Anti-Infect. Ther. 2019, 17, 1011–1041. [Google Scholar] [CrossRef]
- Sneppen, K.; Semsey, S.; Seshasayee, A.S.N.; Krishna, S. Restriction modification systems as engines of diversity. Front. Microbiol. 2015, 6, 528. [Google Scholar] [CrossRef]
- Moghadam, M.T.; Amirmozafari, N.; Shariati, A.; Hallajzadeh, M.; Mirkalantari, S.; Khoshbayan, A.; Jazi, F.M. How phages overcome the challenges of drug resistant bacteria in clinical infections. Infect. Drug Resist. 2020, 13, 45–61. [Google Scholar] [CrossRef]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging strategies to combat ESKAPE pathogens in the era of antimicrobial resistance: A review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef]
- Chan, B.K.; Abedon, S.T.; Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef]
- Haines, M.E.K.; Hodges, F.E.; Nale, J.Y.; Mahony, J.; Van Sinderen, D.; Kaczorowska, J.; Alrashid, B.; Akter, M.; Brown, N.; Sauvageau, D.; et al. Analysis of selection methods to develop novel phage therapy cocktails against antimicrobial resistant clinical isolates of bacteria. Front. Microbiol. 2021, 12, 564. [Google Scholar] [CrossRef]
- Ma, Y.; Pacan, J.C.; Wang, Q.; Xu, Y.; Huang, X.; Korenevsky, A.; Sabour, P.M. Microencapsulation of bacteriophage Felix O1 into chitosan-alginate microspheres for oral delivery. Appl. Environ. Microbiol. 2008, 74, 4799–4805. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Van Belleghem, J.D.; de Vries, C.R.; Burgener, E.; Chen, Q.; Manasherob, R.; Aronson, J.R.; Amanatullah, D.F.; Tamma, P.D.; Suh, G.A. The safety and toxicity of phage therapy: A review of animal and clinical studies. Viruses 2021, 13, 1268. [Google Scholar] [CrossRef] [PubMed]
- Fauconnier, A. Phage therapy regulation: From night to dawn. Viruses 2019, 11, 352. [Google Scholar] [CrossRef] [PubMed]
- Gigante, A.; Atterbury, R.J. Veterinary use of bacteriophage therapy in intensively-reared livestock. Virol. J. 2019, 16, 1–9. [Google Scholar] [CrossRef]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage applications for food production and processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef]
- Nagel, T.E.; Chan, B.K.; De Vos, D.; El-Shibiny, A.; Kang’Ethe, E.K.; Makumi, A.; Pirnay, J.-P. The developing world urgently needs phages to combat pathogenic bacteria. Front. Microbiol. 2016, 7, 882. [Google Scholar] [CrossRef]


{kind=link}
| Country | Prevalence | Sample | Year | References |
|---|---|---|---|---|
| India | 4.82% | Eggs | 2006–2007 | [36] |
| Bangladesh | 28% and 83%, 3% | Eggs and eggshell, egg content | 2014–2015 and 2011–2012 | [37,38] |
| Ethiopia | 2.4%, 4.8%, and 5.3% | Egg content, eggshell and egg from market | 2018 and 2012–2013 | [39,40] |
| China | 6.6%, 5.5% | Eggs from poultry, eggs from marketplace | 2013–2014 | [41] |
| Guangdong, China | 5.4% | Eggs | 2017–2018 | [42] |
| Iran | 13.61% | Eggshell | 1996–2018 | [43] |
| Pakistan | 29.36% and 10.31% | Eggshell and egg content | 2011–2012 | [44] |
| Kuwait | 10% | Eggs | 2004–2005 | [45] |
| Iraq | 4.85% | Eggshell | 2016 | [46] |
| Thailand | 12.4%, 11% | Eggshell, egg content | 1992 | [47] |
| Nigeria | 7.3% | Eggs | 2019 | [48] |
| Brazil | 1.25% | Eggshell, egg content | n/a | [49] |
| Sri Lanka | 6.7% | Eggs | n/a | [50] |
| Zambia | 2.31% | Eggshell | 2018 | [51] |
| Uruguay | 0.0049% | Egg content | 2010 | [52] |
| Japan | 0.25% | Eggshell | 2007–2008 | [53] |
| South Korea | 7.4% | Egg content | 2010–2012 | [54] |
| Colombia | 2.93% | Eggshell | 2014 | [55] |
| Cameroon | 88.6% | Eggs | 2016 | [56] |
| Ireland | 0.04% | Egg contents | 2005–2006 | [57] |
| Origin | Sample Type | Dominant Serotype | Phenotypic Resistance | Reference |
|---|---|---|---|---|
| Bangladesh | Cloacal swab, feed, litter | Salmonella Typhimurium | The percentage of resistance to tetracycline, chloramphenicol, ampicillin, and streptomycin were 97.14%, 94.28%, 82.85%, and 77.14%, respectively. | [70] |
| Bangladesh | Chicken samples (liver and intestine) | Salmonella spp. | High percentage of resistance were found against colistin (92.68%) and ciprofloxacin (73.17%), followed by tigecycline (62.20%), co-trimoxazole (60.98%). | [71] |
| Henan, China | Dead chicken | Salmonella Pullorum, Salmonella Enteritidis | 77%, 73%, 5.60% of isolates were resistant to ciprofloxacin, sulfisoxazole, and ampicillin, respectively; 69.64% were resistant to three or more antimicrobials. | [72] |
| Egypt | Broiler chicken | Salmonella Enteritidis, Salmonella Typhimurium | 76.7% isolates were multidrug resistant, resistant to sulfamethoxazole (100%), amoxicillin–clavulanic acid (68%), streptomycin (65%). | [73] |
| Iran | Fecal swab | Salmonella Enteritidis | Resistant to nitrofurantoin (90.2%), followed by nalidixic acid (67.2%), and cephalexin (37.7%). Multi-drug resistance characteristics were found in 57.4% isolates. | [74] |
| Pakistan | Poultry postmortem | Salmonella Infantis | Isolates showed maximum resistance against pefloxacin (94.4%), chloramphenicol (83.3%), and imipenem (77.7%). | [75] |
| Eastern region, China | Fecal swab | Salmonella Indiana, Salmonella Enteritidis | Isolates were resistant to sulfamethoxazole, ampicillin, tetracycline, doxycycline, and trimethoprim. | [76] |
| South Korea | Chicken meat, feces, and eggshells | Salmonella Enteritidis | All isolates were found to be resistant to at least 1 of 21 antibiotics, 65.2% were resistant to three or more antimicrobials, namely penicillins, sulfisoxazole, streptomycin, tetracyclines, quinolones. | [77] |
| India | Eggs, cloacal swabs, feces | Salmonella Typhimurium | All the isolates showed resistance to clindamycin, oxacillin, penicillin, and vancomycin at varying degree. | [78] |
| Romania | Chicken meat | Salmonella Infantis | 66.6% of isolates were resistant to tetracycline, followed by nalidixic acid and sulfamethoxazole (64.3%), ciprofloxacin (61.9%), streptomycin (59.5%). | [79] |
| South Africa | Chicken carcass swabs, cloacal swabs | Salmonella Bovismorbificans, Salmonella Hadar, Salmonella Dublin, Salmonella Enteritidis | The frequency of MDR among the Salmonella isolates was 81.8%, highest to erythromycin (94.9%) and spectinomycin (82.7%). | [80] |
| Malaysia | Cloaca swab | Salmonella spp. | Resistance to erythromycin (100%), chloramphenicol (76.2%), tetracycline (62%), ampicillin (47.7%), sulfamethoxazole/trimethoprim (42.9%). | [81] |
| Thailand | Chicken meat | Salmonella spp. | Nalidixic acid had the highest rate of resistance (31%), followed by ampicillin (24%), tetracycline (19%), and sulfamethoxazole trimethoprim (8%). | [82] |
| Greece | Chicken carcass and liver | Salmonella Hadar, Salmonella Enteritidis, | The percentage of resistance to streptomycin, tetracycline, nalidixic acid, ampicillin, and rifampicin were 64.5%, 56.2%, 39.5%, and 33.3%, respectively. | [83] |
| Vietnam | Chicken carcasses | Salmonella Albany, | 73.3% isolates were resistant to at least one antibiotic with highest resistance to tetracycline (59.1%) and ampicillin (41.6%); 17.7% multidrug-resistance was also observed. | [84] |
| Singapore | Chicken meat | Salmonella Saintpaul | 59.6% isolates were multidrug-resistant. Phenotypic resistance to ampicillin, tetracycline and chloramphenicol, sulfamethoxazole-trimethoprim and nalidixic acid were 78.8%, 61.5%, 55.8%, and 30.5%, respectively. | [85] |
| Colombia | Chicken carcasses | Salmonella Paratyphi B | The percentage of Salmonella isolates resistant to 1–5, 6–10, and 11–15 antimicrobial agents were 35.2%, 24.6%, and 33.9%, respectively. | [86] |
| Turkey | Chicken meat | Salmonella spp. | High degree of resistance (≥89.2) to vancomycin, tetracycline, streptomycin was observed. | [87] |
| Myanmar | Chicken meat | Salmonella Albany | 52.2% isolates were multidrug-resistant. High frequency of resistance to trimethoprim-sulfamethoxazole (70.3%), tetracycline (54.3%), streptomycin (49.3%), ampicillin (47.1%) was found. | [88] |
| Canada | Chicken meat | Salmonella Hadar | About 21% of chicken isolates were resistant to amoxicillin–clavulanic acid, ceftiofur, and ceftriaxone. | [89] |
| Argentina | Chicken liver | Salmonella Schwarzengrund | All isolates were found to be sensitive to all tested antibiotics except 100% resistant to erythromycin. | [90] |
| Experimental Model | Phage | Inoculation Dose | Phage Delivery Method | Outcomes (Compared with Control) | Reference |
|---|---|---|---|---|---|
| Broiler chicken | Three Salmonella phage | 109–11 PFU | Oral | Phage reduced cecal colonization of Salmonella Enteritidis and Salmonella Typhimurium by ≥4.2 log10 CFU and ≥2.9 log10 CFU, respectively, within 24 h. | [21] |
| Leghorn chicken specific-pathogen-free (SPF) | Three-phage cocktail | 1010 PFU | Oral | When the bacteriophage cocktail was given 1 day before or immediately after bacterial infection, and then again on different days following infection, there was a decrease in Salmonella concentration in the chicken cecum. | [112] |
| Broiler chicken | Three-phage cocktail |
1011 PFU | Oral | The colony-forming units of Salmonella Enteritidis PT4 per gram of cecal content were reduced by 3.5 orders of magnitude in the bacteriophage-treated group. | [113] |
| Chicken carcasses | Salmonella spp. phage | 10⁹ PFU/mL | Spraying | No Salmonella Enteritidis was detected in two trials and more than 70% reduction was achieved in the other two trials. | [114] |
| SPF chicks | Salmonella spp. phage | 1.18 × 1011 PFU–1.03 × 102 PFU | Oral | Cecal contents indicated a moderate decrease in Salmonella loads at 3 days post infection (dpi), with a greater reduction at 5 days post infection (dpi). All of the chicks were negative for Salmonella from 7 dpi through the completion of the trial at 15 dpi. | [115] |
| Broiler chicks | Mixture of bacteriophage | 2.5 × 109–7.5 × 109 PFU | Oral | Compared to untreated controls, Salmonella Enteritidis retrieved from cecal contents was reduced at 12 and 24 h following treatment. | [116] |
| One-day-old chicks | Bacteriophage ΦCJ07 | 105, 107 and 109 PFU | Oral | In challenged and contact chickens, all treatments reduced intestinal Salmonella colonization; after 3 weeks of treatment, no intestinal Salmonella was detected in 70% of contact hens treated with 109 PFU/g of bacteriophage. | [117] |
| Seven-day old chickens | Three different Salmonella-specific bacteriophages | 103 PFU | Spray | When competitive exclusion plus bacteriophage was used, the mean Salmonella Enteritidis cecal count decreased (1.6 × 102 CFU/g) compared to the control group (1.56 × 105 CFU/g). | [118] |
| Six-week-old chickens | Salmonella Gallinarum (SG)-specific bacteriophage | 106 PFU | Oral | In comparison to untreated contact hens, contact hens treated with the bacteriophage showed a considerable reduction in mortality. | [119] |
| Broiler chicks | Salmonella Enteritidis phage | 108 PFU | Oral | On day of trial 14, bacteriophage treatments significantly reduced the incidence of Salmonella Enteritidis in cloacal swabs. | [120] |
| Broiler chicks | P22hc-2, cPII and cI-7 and Felix 0 | 5 × 1011 PFU | Oral | In phage-treated hens, average cecal bacterial counts were 0.3–1.3 orders of magnitude lower than in untreated controls. | [121] |
| Ten-day old chickens | Three lytic phages | 103 PFU | Spray and Oral | Aerosol-spray of bacteriophages resulted in 72.7% decrease in the incidence of Salmonella Enteritidis infection. In addition, counts of Salmonella Enteritidis indicated that phage administration by coarse spray and drinking water decreased the bacteria′s colonization in the gut. | [122] |
| White Leghorn chicks | Φ st1 | 1012 PFU/mL | Intracloacal inoculation | Within 6 h of post-challenge, the Salmonella count had dropped to 2.9 log10 CFU/mL, and Salmonella Typhimurium was undetectable at and after 24 h. | [123] |
| Eggs | PSE5 | 4 × 107 PFU | Immersion | A reduction by 2 × 106 CFU/mL of Salmonella was achieved after phage treatment. | [110] |
| Liquid egg | Pu20 | 108 or 109 PFU/mL | Direct inoculation | At 4 °C and 25 °C for 24 h, the quantity of live bacteria in the treatment group reduced by up to 1.06 log10 CFU/mL and 1.12 log10 CFU/mL, respectively, and the highest antibacterial efficacy was 91.30% and 92.40%, respectively, when multiplicity of infection (MOI) = 1000. | [124] |
| Liquid whole egg | Two phages (OSY-STA and OSY-SHC) | n/a | Direct inoculation | 1.8 and >2.5 log CFU/mL reduction in Salmonella Typhimurium and Salmonella Enteritidis, respectively. | [125] |
| Chicken breasts and fresh eggs | UAB_Phi 20, UAB_Phi78, and UAB_Phi87 | 109 PFU/mL and 1 × 1010 PFU | Soaking in suspension and spraying | Salmonella reduction was >1 log10 CFU/g in chicken breasts. In fresh eggs, a reduction of 0.9 log10 CFU/cm2 in Salmonella was observed. | [126] |
| Raw chicken breast | Five Salmonella phages | 3 × 108 PFU | Suspension added on surface | The largest reductions in the number of Salmonella Enteritidis and Salmonella Typhimurium in phage-treated group were 3.06 and 2.21 log CFU/piece, respectively, when incubated at 25 °C. | [127] |
| Chicken breast | Two-phage cocktails | 4 × 109 PFU/mL | Added on surface | After 5 h, the Salmonella Enteritidis concentration on chicken breast was reduced by 2.5 log CFU/sample | [128] |
| Chicken breasts | SPHG1 and SPHG3 | 8.3 log10 PFU | Spotted | The phage cocktail was applied to chicken breasts at MOIs of 1000 or 100, and the viable count of Salmonella Typhimurium was significantly reduced. | [129] |
| Chicken breast meat | Four Salmonella phage | 108, 109, and 1010 PFU/mL | Directly added | When raw chicken breast samples were treated with a cocktail of all four bacteriophages at 4 °C for 7 days, viable cell counts of bacteria were considerably reduced. | [130] |
| Chicken breast fillets | Salmonella lytic bacteriophage preparation | 109 PFU/ml | Spraying | Salmonella reductions of 1.6–1.7 and 2.2–2.5 log CFU/cm2 were achieved with the use of chlorine and PAA followed by phage spray. | [131] |
| Chicken skin | Eϕ151, Tϕ7 phage suspension | 109 PFU | Spray | Salmonella reductions were 1.38 log10 MPN (Enteritidis) and 1.83 log10 MPN (Typhimurium) per skin area following phage treatment. | [17] |
| Chicken skin | vB_StyS-LmqsSP1 | 2.5 × 108 PFU/cm2 | Direct addition | Phage treatment of chicken skin resulted in about 2 log units reduction in Salmonella isolates from the first 3 h throughout a 1-week experiment at 4 ℃. | [132] |
| Raw chicken meat and chicken skin | SE-P3, P16, P37, and P47 | 10⁹ PFU | Direct inoculation | Throughout storage at 4 and 25 °C, phages reduced the number of viable Salmonella cells in samples containing 103 CFU/g to undetectable levels. | [133] |
| Chicken meat | Five bacteriophages | 109 PFU/mL | Direct inoculation | Compared to control, application of phage cocktail results in 1.4 logarithmic unit reduction at 10 ℃ at 48 h. | [134] |
| Chicken meat | Three lytic bacteriophages Ic_pst11, Is_pst22, and Is_pst24 | 108, 107, and 106 PFU/mL | Direct addition | At MOIs of 100, 1000, and 10,000, a substantial decline in the viable count of Salmonella Typhimurium was seen at 7 h after phage application with reductions of 1.17, 1.26, and 1.31 log10 CFU/g. | [135] |
| Chicken meat | STGO-35-1 | 4 × 106 PFU/mL | Direct addition | Phage treatment caused a significant 2.5 log10 reduction of Salmonella Enteritidis. | [136] |
| Chicken frankfurters | Felix O1 | 5.25 × 106 PFU | Direct addition of liquid | Suppression levels of 1.8 and 2.1 log units of Salmonella Typhimurium were achieved by two variants of phages. | [137] |
| Duck meat | fmb-p1 | 9.9 × 109 PFU | Direct inoculation | 4.52 log CFU/cm2 reduction in Salmonella Typhimurium counts in ready-to-eat duck meat was found. | [138] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.A.S.; Rahman, S.R. Use of Phages to Treat Antimicrobial-Resistant Salmonella Infections in Poultry. Vet. Sci. 2022, 9, 438. https://doi.org/10.3390/vetsci9080438
Khan MAS, Rahman SR. Use of Phages to Treat Antimicrobial-Resistant Salmonella Infections in Poultry. Veterinary Sciences. 2022; 9(8):438. https://doi.org/10.3390/vetsci9080438
Chicago/Turabian StyleKhan, Md Abu Sayem, and Sabita Rezwana Rahman. 2022. "Use of Phages to Treat Antimicrobial-Resistant Salmonella Infections in Poultry" Veterinary Sciences 9, no. 8: 438. https://doi.org/10.3390/vetsci9080438