

DNA Polymorphisms and mRNA Levels of Immune Biomarkers as Candidates for Inflammatory Postpartum Disorders Susceptibility in Italian Buffaloes
 ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Dairy Buffaloes and Research Samples
2.2. DNA Extraction and Amplification
2.3. Discovering Polymorphisms
2.4. Levels of Immune Gene Transcripts
2.5. Statistical Analysis
3. Results
3.1. Immune Gene Genetic Polymorphisms
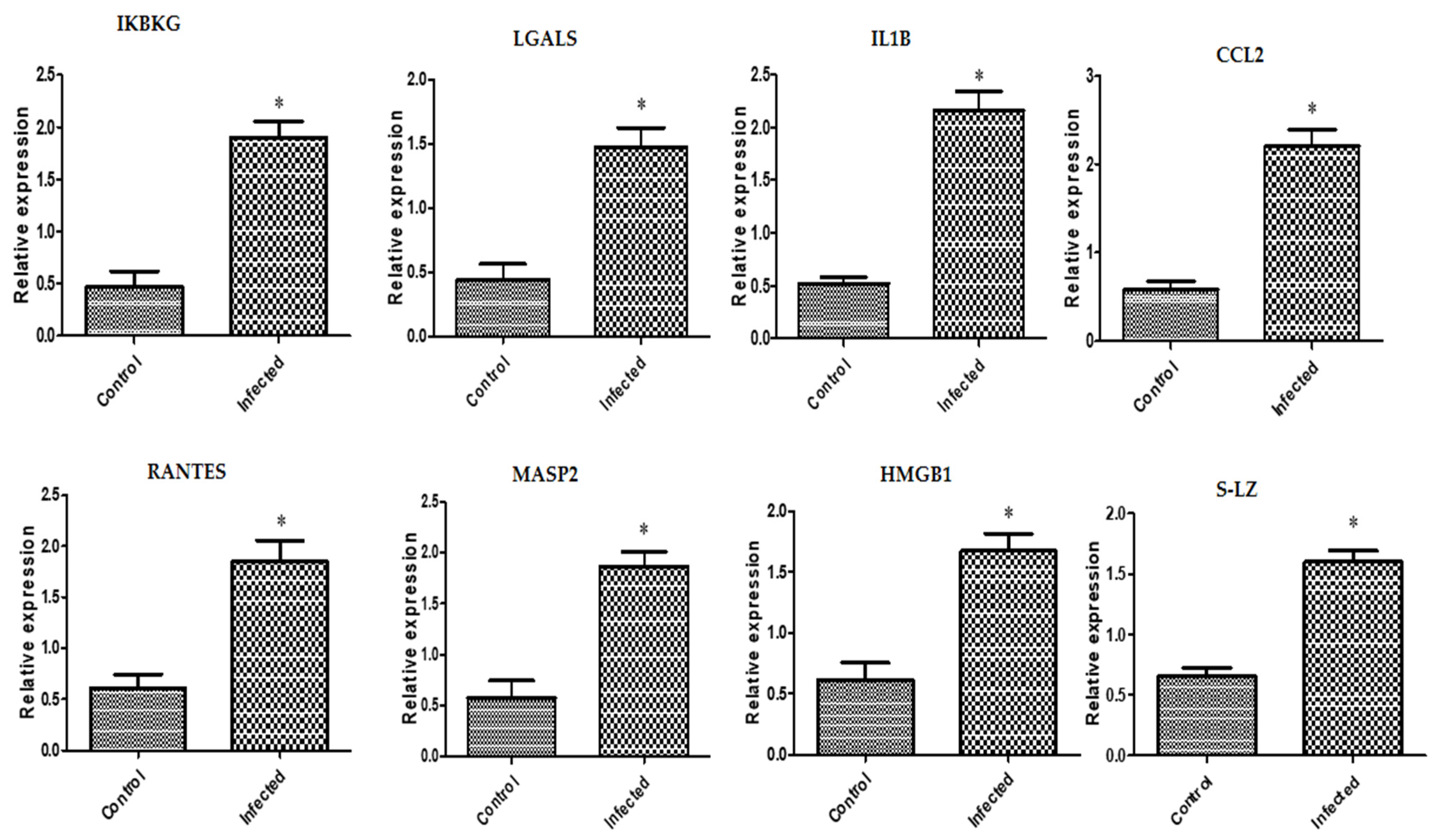
3.2. Immune Markers’ Transcriptional Levels Tendencies
4. Discussion
4.1. Association of Immune Genes Polymorphisms with Inflammatory Reproductive Disorders
4.2. Immune Markers’ Gene Expression Profile
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Wishy, A.B. The postpartum buffalo: I. Endocrinological changes and uterine involution. Anim. Reprod. Sci. 2007, 97, 201–215. [Google Scholar] [CrossRef]
- Patra, M.K.; Kumar, H.; Nandi, S. Neutrophil functions and cytokines expression profile in buffaloes with impending postpartum reproductive disorders. Asian Australas. J. Anim. Sci. 2013, 26, 1406–1415. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Bragaglio, A.; Braghieri, A.; Napolitano, F.; Domínguez-Oliva, A.; Mora-Medina, P.; Álvarez-Macías, A.; De Rosa, G.; Pacelli, C.; José, N.; et al. Dairy Buffalo Behavior: Calving, Imprinting and Allosuckling. Animals 2022, 12, 2899. [Google Scholar] [CrossRef]
- Elsayed, D.H.; El-Azzazi, F.E.; Mahmoud, Y.K.; Dessouki, S.M.; Ahmed, E.A. Subclinical endometritis and postpartum ovarian resumption in respect to TNF-α, IL-8 and CRP in Egyptian buffaloes. Anim. Reprod. 2020, 17, e20190027. [Google Scholar] [CrossRef]
- Ateya, A.; El-Sayed, A.; Mohamed, R. Gene expression and serum profile of antioxidant markers discriminate periparturient period time in dromedary camels. Mammal Res. 2021, 66, 603–613. [Google Scholar] [CrossRef]
- Riveros, J.L.; Urquieta, B.; Bonacic, C.; Hoffmann, B.; Bas, F.; Schuler, G. Endocrine changes during pregnancy, parturition and post-partum in guanacos (Lama guanicoe). Anim. Rep. Sci. 2009, 116, 318–325. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Azawi, O.I.; Omran, S.N.; Hadad, J.J. A study of endometritis causing repeat breeding of cycling Iraqi buffalo cows. Reprod. Domest. Anim. Zuchthyg. 2008, 43, 735–743. [Google Scholar] [CrossRef]
- Nehru, D.A.; Dhaliwal, G.S.; Jan, M.H.; Cheema, R.S.; Kumar, S. Clinical efficacy of intrauterine cephapirin benzathine administration on clearance of uterine bacteria and subclinical endometritis in postpartum buffaloes. Reproduction in domestic animals. Zuchthygiene 2019, 54, 317–324. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Williams, E.J.; Miller, A.N.; Nash, D.M.; Herath, S. Uterine diseases in cattle after parturition. Vet. J. 2008, 176, 115–121. [Google Scholar] [CrossRef]
- Del Vecchio, R.P.; Matsas, D.J.; Fortin, S.; Sponenberg, D.P.; Lewis, G.S. Spontaneous uterine infections are associated with elevated prostaglandin F2α metabolite concentrations in postpartum dairy cows. Theriogenology 1994, 41, 413–421. [Google Scholar] [CrossRef]
- Dohmen, M.J.; Joop, K.; Sturk, A.; Bols, P.E.; Lohuis, J.A. Relationship between intra-uterine bacterial contamination, endotoxin levels and the development of endometritis in postpartum cows with dystocia or retained placenta. Theriogenology 2000, 54, 1019–1032. [Google Scholar] [CrossRef]
- Donnez, J.; Cacciottola, L. Endometriosis: An Inflammatory Disease That Requires New Therapeutic Options. Int. J. Mol. Sci. 2022, 23, 1518. [Google Scholar] [CrossRef]
- Singh, H.; Brar, P.S.; Honparkhe, M.; Arora, A.K.; Dhindsa, S.S. Subclinical endometritis in estrual buffaloes: Diagnosis, prevalence and impact on reproductive performance. Trop. Anim. Health Prod. 2020, 52, 357–363. [Google Scholar] [CrossRef]
- Azawi, O.I.; Omran, S.N.; Hadad, J.J. Clinical, bacteriological, and histopathological study of toxic puerperal metritis in Iraqi buffalo. J. Dairy Sci. 2007, 90, 4654–4660. [Google Scholar] [CrossRef]
- Buzzaccarini, G.; Vitagliano, A.; Andrisani, A.; Santarsiero, C.M.; Cicinelli, R.; Nardelli, C.; Ambrosini, G.; Cicinelli, E. Chronic endometritis and altered embryo implantation: A unified pathophysiological theory from a literature systematic review. J. Assist. Reprod. Genet. 2020, 37, 2897–2911. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Roles of toll-like receptors in innate immune responses. Genes Cells 2001, 6, 733–742. [Google Scholar] [CrossRef]
- Saed, H.A.R.; Ibrahim, H.M.M.; El-Khodery, S.A.; Youssef, M.A. Relationship between expression pattern of vitamin D receptor, 1 alpha-hydroxylase enzyme, and chemokine RANTES genes and selected serum parameters during transition period in Holstein dairy cows. Vet. Rec. Open 2020, 7, e000339. [Google Scholar] [CrossRef]
- Prakash, O.; Kumar, A.; Sonwane, A.; Rathore, R.; Singh, R.V.; Chauhan, A.; Kumar, P.; Renjith, R.; Yadav, R.; Bhaladhare, A.; et al. Polymorphism of cytokine and innate immunity genes associated with bovine brucellosis in cattle. Mol. Biol. Rep. 2014, 41, 2815–2825. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, A.J.; Hayes, B.J.; Savin, K.; Bolormaa, S.; McPartlan, H.C.; Bowman, P.J.; Van der Jagt, C.; MacEachern, S.; Goddard, M.E. Validation of single nucleotide polymorphisms associated with milk production traits in dairy cattle. J. Dairy Sci. 2012, 95, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Nayeri, S.; Stothard, P. Tissues, Metabolic Pathways and Genes of Key Importance in Lactating Dairy Cattle. Springer Sci. Rev. 2016, 4, 49–77. [Google Scholar] [CrossRef]
- Sise, J.A.; Penty, J.M.; Montgomery, G.W.; Gootwine, E. The duplicated gene copy of the ovine growth hormone gene contains a PvuII polymorphism in the second intron. Anim. Genet. 1993, 24, 319–321. [Google Scholar] [CrossRef]
- Barbieri, M.E.; Manfredi, E.; Elsen, J.M.; Ricordeau, G.; Bouillon, J.; Grosclaude, F.; Mahé, M.F.; Bibé, B. Influence du locus de la caseine a(s1) sur les performances laitieres et les parametres genetiques des chevres de race Alpine. Genet. Sel. Evol. 1995, 27, 437–450. [Google Scholar] [CrossRef]
- Moioli, B.; D’Andrea, M.; Pilla, F. Candidate genes affecting sheep and goat milk quality. Small Rumin. Res. 2007, 68, 179–192. [Google Scholar] [CrossRef]
- Leymaster, K.; Freking, B. Genetic variations and associations for improving meat production and meat qualities in sheep and goats. In 6th Proceedings of the World Congress on Genetics Applied to Livestock Production; University of New England: Armidale, Australia, 1998; pp. 109–116. [Google Scholar]
- Diez-Tascón, C.; Keane, O.M.; Wilson, T.; Zadissa, A.; Hyndman, D.L.; Baird, D.B.; McEwan, J.C.; Crawford, A.M. Microarray analysis of selection lines from outbred populations to identify genes involved with nematode parasite resistance in sheep. Physiol. Genom. 2005, 21, 59–69. [Google Scholar] [CrossRef]
- Darwish, A.; Ebissy, E.; Ateya, A.; El-Sayed, A. Single nucleotide polymorphisms, gene expression and serum profile of immune and antioxidant markers associated with postpartum disorders susceptibility in Barki sheep. Anim. Biotechnol. 2021, 34, 327–339. [Google Scholar] [CrossRef]
- Fouda, M.; Hemeda, S.; El-Bayomi, K.; El-Araby, I.; Ateya, A.; Hendam, B. Genetic polymorphisms in FSHR/ ALUI and ESRa/BG1I loci and their association with repeat breeder incidence in buffalo. J. Hellenic Vet. Med. Soc. 2021, 72, 2869–2878. [Google Scholar]
- Al-Sharif, M.; Radwan, H.; Hendam, B.; Ateya, A. DNA polymorphisms of FGFBP1, leptin, κ-casein, and as1-casein genes and their association with reproductive performance in dromedary she-camels. Theriogenology 2022, 178, 18–29. [Google Scholar] [CrossRef]
- Boom, R.; Sol, C.J.; Salimans, M.M.; Jansen, C.L.; Wertheim-van Dillen, P.M.; Noordaa, J.V.D. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef]
- Boesenberg-Smith, K.A.; Pessarakli, M.M.; Wolk, D.M. Assessment of DNA Yield and Purity: An Overlooked Detail of PCR Troubleshooting. Clin. Microbiol. Newsl. 2012, 34, 1–6. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) Software Version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Doran, J.; Walters, C.; Kyle, V.; Wooding, P.; Hammett-Burke, R.; Colledge, W.H. Mfsd14a (Hiat1) gene disruption causes globozoospermia and infertility in male mice. Reproduction 2016, 152, 91–99. [Google Scholar] [CrossRef]
- Kasprzak-Filipek, K.; Wioletta Sawicka-Zugaj, W.S.; Litwinczuk, Z.; Chabuz, W.; Sveistiene, R.; Bulla, J. Assessment of the genetic structure of Central European cattle breeds based on functional gene polymorphism. Glob. Ecol. Conserv. 2018, 17, e00525. [Google Scholar] [CrossRef]
- Singh, U.; Deb, R.; Alyethodi, R.R.; Alex, R.; Kumar, S.; Chakraborty, S.; Dhama, K.; Sharma, A. Molecular markers and their applications in cattle genetic research: A review. Biomark. Genom. Med. 2014, 6, 49–58. [Google Scholar] [CrossRef]
- Mossallam, A.A.; El Nahas, S.M.; Osman, N.M.; Shahwan, E.-H.T.; Sabry, N.M. Sequence analysis of TLR4 gene in river buffalo (Egyptian breed) and SNPs association with Mastitis. Bull. Natl. Res. Cent. 2023, 47, 110. [Google Scholar] [CrossRef]
- Osman, M.N.; Abou Mossallam, A.A.; El Seedy, R.F.; Mahfouz, R.E. Single Nucleotide Polymorphisms in TLR4 Gene and Endometritis Resistance in River Buffalo (Bubalus bubalis). Jordan J. Biol. Sci. 2018, 11, 577–583. [Google Scholar]
- Abou Mossallam, A.A.; Osman, M.N.; Othman, O.E.; Mahfouz, R.E. Polymorphism evaluation of TLR2 gene associated with endometritis infection in buffalo reared in Egypt. Biotechnology 2022, 26, 45–55. [Google Scholar] [CrossRef]
- Al-Sharif, M.; Abdo, M.; Shabrawy, O.E.; El-Naga, E.M.A.; Fericean, L.; Banatean-Dunea, I.; Ateya, A. Investigating polymorphisms and expression profile of immune, antioxidant, and erythritol-related genes for limiting postparturient endometritis in Holstein cattle. Vet. Sci. 2023, 23, 370. [Google Scholar] [CrossRef]
- Ateya, A.I.; Ibrahim, S.S.; Al-Sharif, M.M. Single Nucleotide Polymorphisms, Gene Expression and Economic Evaluation of Parameters Associated with Mastitis Susceptibility in European Cattle Breeds. Vet. Sci. 2022, 14, 294. [Google Scholar] [CrossRef]
- Garcıa-Montoya, I.A.; Cendon, T.S.; Ar evalo-Gallegos, S.; Rascon-Cruz, Q. Lactoferrin a multiple bioactive protein: An overview. Biochim. Biophys. Acta 2012, 1820, 226–236. [Google Scholar] [CrossRef]
- Ateya, A.I.; El-Seady, Y.Y.; Atwa, S.M.; Merghani, B.H.; Sayed, N.A. Novel single nucleotide polymorphisms in lactoferrin gene and their association with mastitis susceptibility in Holstein cattle. Genetika 2016, 48, 199–210. [Google Scholar] [CrossRef]
- Guo, B.; Jiao, Y.; He, C.; Wei, L.; Chang, Z.; Yue, X.; Lan, X.; Chen, H.; Lei, C. A novel polymorphism of the lactoferrin gene and its association with milk composition and body traits in dairy goats. Genet. Mol. Res. 2010, 9, 2199–2206. [Google Scholar] [CrossRef]
- Swiderek, W.P.; Bhide, M.R.; Gruszczynska, J.; Soltis, K.; Witkowska, D.; Mikula, I. Toll-like receptor gene polymorphism and its relationship with somatic cell concentration and natural bacterial infections of the mammary gland in sheep. Folia Microbiol. 2006, 51, 647–652. [Google Scholar] [CrossRef]
- Sharma, B.S.; Leyva, I.; Schenkel, F.; Karrow, N.A. Association of toll-like receptor 4 polymorphisms with somatic cell score and lactation persistency in Holstein Bulls. J. Dairy Sci. 2006, 89, 3626–3635. [Google Scholar] [CrossRef]
- Wang, X.; Xu, S.; Gao, X.; Ren, H.; Chen, J. Genetic polymorphism of TLR4 gene and correlation with mastitis in cattle. J. Genet. Genom. 2007, 34, 406–412. [Google Scholar] [CrossRef]
- Ruiz-Rodriguez, C.T.; Brandt, J.R.; Oliverio, R.; Ishida, Y.; Guedj, N.; Garrett, E.F.; Bar-Gal, G.K.; Nikolaidis, N.; Cardoso, F.C.; Roca, A.L. Polymorphisms of the toll-like receptor 2 of goats (Capra hircus) may be associated with somatic cell count in milk. Anim. Biotechnol. 2017, 28, 112–119. [Google Scholar] [CrossRef]
- Barton, N.H. Mutation and the evolution of recombination. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 1281–1294. [Google Scholar] [CrossRef] [PubMed]
- Dakal, T.C.; Kala, D.; Dhiman, G.; Yadav, V.; Krokhotin, A.; Dokholyan, N.V. Predicting the functional consequences of non-synonymous single nucleotide polymorphisms in IL8 gene. Sci. Rep. 2017, 7, 6525. [Google Scholar] [CrossRef] [PubMed]
- Taha, A.D.; Mahfouz, R.E.; Bibars, A.M.; Hassan, A.N.; Othman, O.E. Cytokine genes expression in uteri of Bubalus bubalis associated with endometritis infection. Jordan J. Biol. Sci. 2021, 14, 245–251. [Google Scholar]
- Fagundes, N.S.; Rezende, A.L.; Alvarenga, P.B.; Magalhães, L.Q.; Santos, R.M.; Headley, S.A.; Silva, M.J.B.; Beletti, M.E.; Saut, J.P.E. Short communication: Proinflammatory gene expression relative to the collection technique of endometrial samples from cows with and without subclinical endometritis. J. Dairy Sci. 2019, 102, 5511–5517. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, L.; Sun, Y.; Huang, S.; Tang, J.; Yu, P.; Wang, G. Altered Molecular Expression of the TLR4/NF-κB Signaling Pathway in Mammary Tissue of Chinese Holstein Cattle with Mastitis. PLoS ONE 2015, 10, e0118458. [Google Scholar] [CrossRef]
- Raliou, M.; Dembélé, D.; Düvel, A.; Bolifraud, P.; Aubert, J.; Mary-Huard, T.; Rocha, D.; Piumi, F.; Mockly, S.; Heppelmann, M.; et al. Subclinical endometritis in dairy cattle is associated with distinct mRNA expression patterns in blood and endometrium. PLoS ONE 2019, 14, e0220244. [Google Scholar] [CrossRef]
- Yakan, A.; Ozkan, H.; Eraslan, A.; Unal, N.; OZbeyaz, C. Gene expression levels in some candidate genes for mastitis resistance, milk yield, and milk quality of goats reared under different feeding systems. Turk. J. Vet. Anim. Sci. 2018, 42, 18–28. [Google Scholar] [CrossRef]
- Pisoni, G.; Moroni, P.; Genini, S.; Stella, A.; Boettcher, P.; Cremonesi, P.; Scaccabarozzi, L.; Giuffra, E.; Castiglioni, B. Differentially expressed genes associated with Staphylococcus aureus mastitis in dairy goats. Vet. Immunol. Immunopathol. 2010, 135, 208–217. [Google Scholar] [CrossRef]
- Shao, C.; Wang, H.; Wang, X.; Jiang, S.; Sun, J.; Song, H.; Li, J. Characterization of inflammatory responses by cervical cytology, cytokine expression and ultrastructure changes in a goat subclinical endometritis model. J. Vet. Med. Sci. 2017, 79, 197–205. [Google Scholar] [CrossRef]
- Frans, G.; Bosch, J.v.d.W.T.; Moens, L.; Gijsbers, R.; Changi-Ashtiani, M.; Rokni-Zadeh, H.; Shahrooei, M.; Wuyts, G.; Meyts, I.; Bossuyt, X. Functional Evaluation of an IKBKG Variant Suspected to Cause Immunodeficiency Without Ectodermal Dysplasia. J. Clin. Immunol. 2017, 37, 801–810. [Google Scholar] [CrossRef]
- Dumic, J.; Dabelic, S.; Flögel, M. Galectin-3: An open-ended story. Biochim. Biophys. Acta 2006, 1760, 616–635. [Google Scholar] [CrossRef] [PubMed]
- Henderson, N.C.; Sethi, T. The regulation of inflammation by galectin-3. Immunol. Rev. 2009, 230, 160–171. [Google Scholar] [CrossRef]
- Salim, T.; Sershen, C.L.; May, E.E. Investigating the role of TNF-α and IFN-γ activation on the dynamics of iNOS gene expression in LPS stimulated macrophages. PLoS ONE 2016, 11, e0153289. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, A.I. Chemokine receptors: Multifaceted thera-peutic targets. Nat. Rev. Immunol. 2002, 2, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Olson, T.S.; Ley, K. Chemokines and chemokine receptors inleukocyte trafficking. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R7–R28. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Baca, I.; Schenkel, F.; Sharma, B.S.; Jansen, G.B.; Karrow, N.A. Identification of single nucleotide polymorphisms in the bovine CCL2, IL8, CCR2 and IL8RA genes and their association with health and production in Canadian Holsteins. Anim. Genet. 2007, 38, 198–202. [Google Scholar] [CrossRef]
- Oliva, A.; Kinter, A.L.; Vaccarezza, M.; Rubbert, A.; Catanzaro, A.; Moir, S.; Monaco, J.; Ehler, L.; Mizell, S.; Jackson, R.; et al. Natural killer cells from human immunodeficiency virus (HIV)-infected individuals are an important source of CC-chemokines and suppress HIV-1 entry and replication in vitro. J. Clin. Investig. 1998, 102, 223–231. [Google Scholar] [CrossRef]
- Marques, R.E.; Guabiraba, R.; Russo, R.C.; Teixeira, M.M. Targeting CCL5 in inflammation. Expert Opin. Ther. Targets 2013, 17, 1439–1460. [Google Scholar] [CrossRef]
- Palchevskiy, V.; Hashemi, N.; Weigt, S.S.; Xue, Y.Y.; Derhovanessian, A.; Keane, M.P.; Strieter, R.M.; Fishbein, M.C.; Deng, J.C.; LynchIII, J.P.; et al. Immune response CC chemokines CCL2 and CCL5 are associated with pulmonary sarcoidosis. Fibrogenesis Tissue Repair 2011, 4, 10. [Google Scholar] [CrossRef]
- Kasanmoentalib, E.S.; Valls Seron, M.; Ferwerda, B.; Tanck, M.W.; Zwinderman, A.H.; Baas, F.; van der Ende, A.; Schwaeble, W.J.; Brouwer, M.C.; van de Beek, D. Mannose-binding lectin-associated serine protease 2 (MASP-2) contributes to poor disease outcome in humans and mice with pneumococcal meningitis. J. Neuroinflamm. 2017, 14, 2. [Google Scholar] [CrossRef]
- Bao, F.; Fu, X.; Yu, G.; Wang, Z.; Liu, H.; Zhang, F. Mannose-binding lectin and mannose-binding lectin-associated serine protease-2 genotypes and serum levels in patients with Sporotrichosis. Am. J. Trop. Med. Hyg. 2019, 101, 1322–1324. [Google Scholar] [CrossRef] [PubMed]
- Tulio, S.; Faucz, F.R.; Werneck, R.I.; Olandoski, M.; Alexandre, R.B.; Boldt, A.B.; Pedroso, M.L.; de Messias-Reason, I.J. MASP2 gene polymorphism is associated with susceptibility to hepatitis C virus infection. Hum. Immunol. 2011, 72, 912–915. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Bai, J.Y.; Li, L.; Huang, S.; Li, C.M.; Wang, G.L. Genetic polymorphisms of the BMAP-28 and MASP-2 genes and their correlation with the somatic cell score in Chinese Holstein cattle. Genet. Mol. Res. 2015, 14, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Magna, M.; Pisetsky, D.S. The role of HMGB1 in the pathogenesis of inflammatory and autoimmune diseases. Mol. Med. 2014, 20, 138–146. [Google Scholar] [CrossRef]
- Andersson, U.; Harris, H.E. The role of HMGB1 in the pathogenesis of rheumatic disease. Biochim. Biophys. Acta 2010, 1799, 141–148. [Google Scholar] [CrossRef]
- Salehin, M.; Ghosh, A.K.; Mallick, P.K.; Bhattacharya, T.K. Molecular characterization, polymorphism and association study of lysozyme gene with milk production and somatic cell trait in Bos indicus × Bos taurus cattle. J. Anim. 2009, 3, 623–631. [Google Scholar] [CrossRef]
- Der Günne, H.S.A.; Tenhagen, B.A.; Kutzer, P.; Forderung, D.; Heuwieser, W. Do lactoferrin, lysozyme and the lactoperoxidase–thiocyanate–hydrogen peroxide system cause negative microbiological results in mastitis secretions. Dtsch. Tierarzt. Wochenschr. 2002, 109, 300–305. [Google Scholar]
- Rinaldi, M.; Moroni, P.; Paape, M.J.; Bannerman, D.D. Evaluation of assays for the measurement of bovine neutrophil reactive oxygen species. Vet. Immunol. Immunopathol. 2007, 115, 107–125. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Aitken, S.L. Impact of oxidative stress on the health and immune function of dairy cattle. Vet. Immunol. Immunopathol. 2009, 128, 104–109. [Google Scholar] [CrossRef]
- Duhig, K.; Chappell, L.C.; Shennan, A.H. Oxidative stress in pregnancy and reproduction. Obstet. Med. 2016, 9, 113–116. [Google Scholar] [CrossRef]
- Wang, Z.; Guan, D.; Huo, J.; Biswas, S.K.; Huang, Y.; Yang, Y.; Xu, S.; Lam, K.P. IL-10 enhances human natural killer cell effector functions via metabolic reprogramming regulated by mTORC1 signaling. Front. Immunol. 2021, 23, 619195. [Google Scholar] [CrossRef] [PubMed]
- Masella, R.; Di Benedetto, R.; Varì, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Investigated Gene | Sense | Antisense | Accession Number | Annealing Temperature (°C) | Size of PCR Product (bp) |
|---|---|---|---|---|---|
| IKBKG | 5′-CCGGCTTGGACAGCCTCTCTTG-3′ | 5′-TGCCCTGCTCTGAAGGCACATG-3′ | XM_044936650.2 | 58 | 412 |
| LGALS | 5′-CTCATTTCTCCTGCAGAGTCT-3′ | 5′-AACTTGAATTCGTATCCATCAG-3′ | XM_025282898.3 | 58 | 395 |
| IL1B | 5′-CCATGGCAACCGTACCTGAAC-3′ | 5′- ACTGACTGCACGGGTGCGTCAC-3′ | NM_001290898.1 | 58 | 359 |
| CCL2 | 5′-CTTGAATCCTCTCGCTGCAAC-3′ | 5′-AGATTCTTGGGTTGTGGAGTGA-3′ | HQ889748.1 | 60 | 347 |
| RANTES | 5′-CAGCCGGAGCTGCAGAGGATCA-3′ | 5′- CAAGCTGCTTAGGACAAGAGCG-3′ | XM_006056212.4 | 58 | 399 |
| MASP2 | 5′-AGAGCACAGACACAGAGCGGGC-3′ | 5′- AGATGCGGCCATTAGGTGGCGC-3′ | XR_006551148.1 | 60 | 513 |
| HMGB1 | 5′-CGCTGGCTGGAGAGTAATGTTA-3′ | 5′-CACCAATAGACAGGCCAGGATGT-3′ | XM_006078587.2 | 60 | 540 |
| S-LZ | 5′-TCTGGACATTTGACTTCTCAG-3′ | 5′-CTCAATGTAACTGCTGACGTC-3′ | EF535848.1 | 58 | 455 |
| Investigated Gene | Primer | Product Size (bp) | Annealing Temperature (°C) | GenBank Isolate | Origin |
|---|---|---|---|---|---|
| IKBKG | F5′-CTGGCAGGAGAAGCCATCAA-3 R5′-CAGCAGACAGGACACTAGCC-3′ | 101 | 58 | XM_044936650.2 | Current study |
| LGALS | F5′-GGCCCAAAGCTCATTTCTCC-3 R5′-GCACTCCCCAGGTTTGAGAT-3′ | 129 | 58 | XM_025282898.3 | Current study |
| IL1B | F5′-AGGTGGTGTCGGTCATTGTG-3′ R5′-AACTCGTCGGAGGACGTTTC-3′ | 142 | 60 | NM_001290898.1 | Current study |
| CCL2 | F5′-AAGCCTTGAGCACTCACTCC -3′ R5′-GCAGTTAGGGAAAGCCGGAA-3′ | 72 | 56 | HQ889748.1 | Current study |
| RANTES | F5′-CCCATATGCCTCAGACACCAC-3′ R5′-GGCGGTTCTTCCTGGTGATA-3′ | 140 | 60 | XM_006056212.4 | Current study |
| MASP2 | F5′-ACAGACAAGGCGGAATACGG-3′ R5′-GTGGTCTACAGCAGGCAAGT-3′ | 162 | 58 | XR_006551148.1 | Current study |
| HMGB1 | F5′-TGCCTCGCGGAGGAAAAATA-3′ R5′-GCAGACATGGTCTTCCACCT-3′ | 188 | 60 | XM_006078587.2 | Current study |
| S-LZ | F5′-CTGTAGCCTGTGCAAAGCAG -3′ R5′-CAGGGTGCAACCCTCAATGT-3′ | 113 | 60 | EF535848.1 | Current study |
| ß. actin | F5′-GATGATGATATTGCCGCGCTC-3′ R5′-AGGGTCAGGATGCCTCTCTT-3′ | 197 | 58 | NM_001290932.1 | Current study |
| Gene | SNPs | Healthy n = 60 | Reproductive Disorders n = 60 | Total n = 120 | Sort of Inherited Change | Amino Acid Order and Kind |
|---|---|---|---|---|---|---|
| IKBKG | C78G | 0/60 | 31/60 | 31/120 | Non-synonymous | 26 I to M |
| G328A | 27/60 | 0/60 | 27/120 | Non-synonymous | 110 G to S | |
| LGALS | T192C | 42/60 | 0/60 | 42/120 | Synonymous | 64 N |
| IL1B | C53T | 19/60 | 0/60 | 19/120 | Non-synonymous | 18 T to M |
| T125C | 47/60 | 0/60 | 47/120 | Non-synonymous | 42 V to A | |
| T205C | 0/60 | 23/60 | 23/120 | Non-synonymous | 69 L to S | |
| C311T | 49/60 | 0/60 | 49/120 | Non-synonymous | 104 S to L | |
| CCL2 | C201T | 0/60 | 53/60 | 53/120 | Synonymous | 67 P |
| RANTES | G96C | 0/60 | 29/60 | 29/120 | Synonymous | 32 T |
| A141G | 0/60 | 43/60 | 43/120 | Synonymous | 47 S | |
| C370T | 54/60 | 0/60 | 54/120 | Synonymous | 124 L | |
| MASP2 | T465A | 22/60 | 0/60 | 22/120 | Synonymous | 155 A |
| HMGB1 | C227T | 34/60 | 0/60 | 34/120 | Non-synonymous | 76 S to L |
| C275T | 0/60 | 29/60 | 29/120 | Non-synonymous | 92 P to L | |
| T407C | 56/60 | 0/60 | 56/120 | Non-synonymous | 136 F to S | |
| S-LZ | G33A | 39/60 | 0/60 | 39/120 | Synonymous | 11 R |
| A222G | 0/60 | 52/60 | 52/120 | Synonymous | 74 L | |
| G254A | 0/60 | 37/60 | 37/120 | Non-synonymous | 85 R to K |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ateya, A.; Safhi, F.A.; El-Emam, H.; Al-Ghadi, M.Q.; Abdo, M.; Fericean, L.; Olga, R.; Mihaela, O.; Hizam, M.M.; Mamdouh, M.; et al. DNA Polymorphisms and mRNA Levels of Immune Biomarkers as Candidates for Inflammatory Postpartum Disorders Susceptibility in Italian Buffaloes. Vet. Sci. 2023, 10, 573. https://doi.org/10.3390/vetsci10090573
Ateya A, Safhi FA, El-Emam H, Al-Ghadi MQ, Abdo M, Fericean L, Olga R, Mihaela O, Hizam MM, Mamdouh M, et al. DNA Polymorphisms and mRNA Levels of Immune Biomarkers as Candidates for Inflammatory Postpartum Disorders Susceptibility in Italian Buffaloes. Veterinary Sciences. 2023; 10(9):573. https://doi.org/10.3390/vetsci10090573
Chicago/Turabian StyleAteya, Ahmed, Fatmah A. Safhi, Huda El-Emam, Muath Q. Al-Ghadi, Mohamed Abdo, Liana Fericean, Rada Olga, Ostan Mihaela, Manar M. Hizam, Maha Mamdouh, and et al. 2023. "DNA Polymorphisms and mRNA Levels of Immune Biomarkers as Candidates for Inflammatory Postpartum Disorders Susceptibility in Italian Buffaloes" Veterinary Sciences 10, no. 9: 573. https://doi.org/10.3390/vetsci10090573