
Molecular Detection and Phylogenetic Characterization of Anaplasma spp. in Dogs from Hainan Province/Island, China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
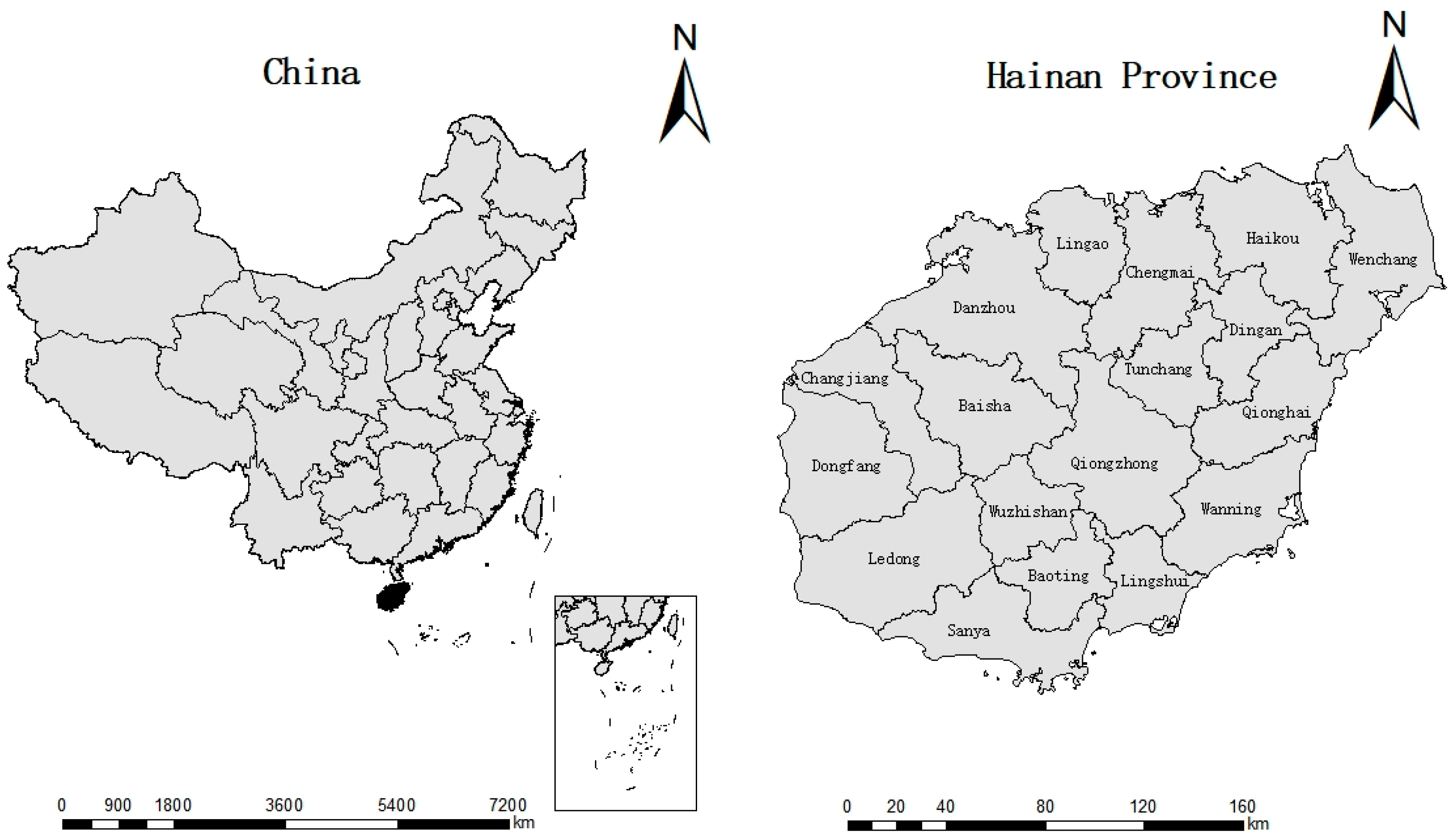
2.1. Study Site
2.2. Sampling
2.3. Nucleic Acid Extraction
2.4. PCR Amplification
2.5. Sequencing and Phylogenetic Analysis
2.6. Statistical Analysis
3. Results
3.1. Burden of Anaplasma spp. Infection in Hainan Province/Island
3.2. Analysis of Relevant Risk Factors
3.3. The Phylogenetic Analysis of the A. phagocytophilum and A. bovis 16S rRNA Gene
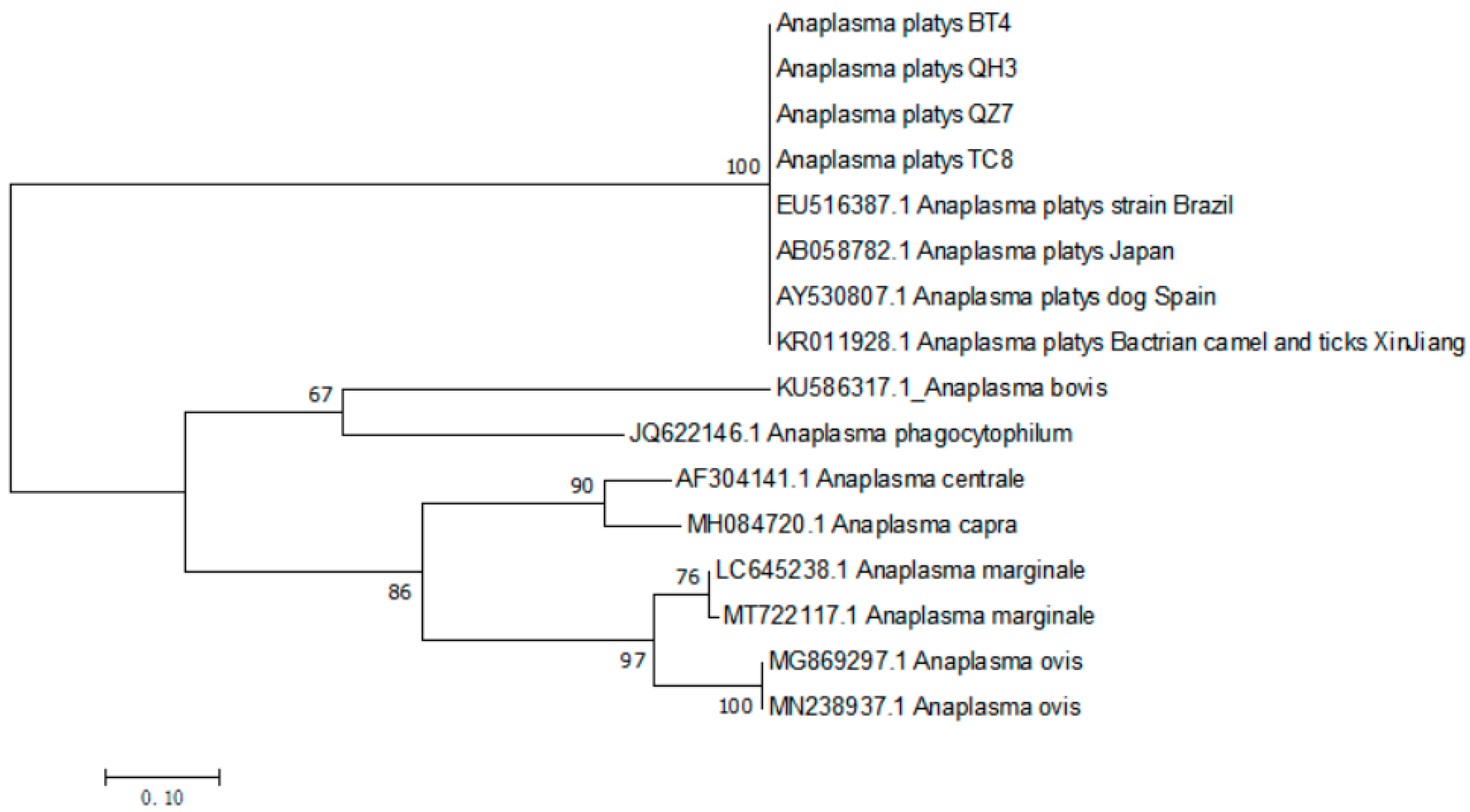
3.4. The Phylogenetic Analysis of A. platys gltA Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rar, V.; Tkachev, S.; Tikunova, N. Genetic diversity of Anaplasma bacteria: Twenty years later. Infect. Genet. Evol. 2021, 91, 104833. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.M.; Dumler, J.S.; Bakken, J.S.; Walker, D.H. Identification of a granulocytotropic Ehrlichia species as the etiologic agent of human disease. J. Clin. Microbiol. 1994, 32, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, Y.; Ni, D.; Li, Q.; Yu, Y.; Yu, X.J.; Wan, K.; Li, D.; Liang, G.; Jiang, X.; et al. Nosocomial transmission of human granulocytic anaplasmosis in China. JAMA 2008, 300, 2263–2270. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.; McBride, J.W. Tick-Borne Emerging Infections: Ehrlichiosis and Anaplasmosis. Clin. Lab. Med. 2017, 37, 317–340. [Google Scholar] [CrossRef]
- Demma, L.J.; Holman, R.C.; McQuiston, J.H.; Krebs, J.W.; Swerdlow, D.L. Epidemiology of human ehrlichiosis and anaplasmosis in the United States, 2001–2002. Am. J. Trop. Med. Hyg. 2005, 73, 400–409. [Google Scholar] [CrossRef]
- Walker, D.H.; Paddock, C.D.; Dumler, J.S. Emerging and re-emerging tick-transmitted rickettsial and ehrlichial infections. Med. Clin. N. Am. 2008, 92, 1345–1361. [Google Scholar] [CrossRef]
- Dong, J.; Olano, J.P.; McBride, J.W.; Walker, D.H. Emerging pathogens: Challenges and successes of molecular diagnostics. J. Mol. Diagn. 2008, 10, 185–197. [Google Scholar] [CrossRef]
- Harvey, J.W.; Simpson, C.F.; Gaskin, J.M. Cyclic thrombocytopenia induced by a Rickettsia-like agent in dogs. J. Infect. Dis. 1978, 137, 182–188. [Google Scholar] [CrossRef]
- Uilenberg, G. Other Ehrlichiosis of Ruminants. In Rickettsial and Chlamydial Diseases of Domestic Animals; Woldehiwet, Z., Ristic, M., Eds.; Oxford Pergamon Press: Oxford, UK, 1993. [Google Scholar]
- Rar, V.; Golovljova, I. Anaplasma, Ehrlichia, and “Candidatus Neoehrlichia” bacteria: Pathogenicity, biodiversity, and molecular genetic characteristics, a review. Infect. Genet. Evol. 2011, 11, 1842–1861. [Google Scholar] [CrossRef]
- Yang, B.; Sun, E.; Wen, Y.; Ye, C.; Liu, F.; Jiang, P.; Tao, X. Molecular evidence of coinfection of Anaplasma species in small ruminants from Anhui Province, China. Parasitol. Int. 2019, 71, 143–146. [Google Scholar] [CrossRef]
- Guo, W.P.; Tie, W.F.; Meng, S.; Li, D.; Wang, J.L.; Du, L.Y.; Xie, G.C. Extensive genetic diversity of Anaplasma bovis in ruminants in Xi’an, China. Ticks Tick Borne Dis. 2020, 11, 101477. [Google Scholar] [CrossRef] [PubMed]
- Sashika, M.; Abe, G.; Matsumoto, K.; Inokuma, H. Molecular survey of Anaplasma and Ehrlichia infections of feral raccoons (Procyon lotor) in Hokkaido, Japan. Vector Borne Zoonotic Dis. 2011, 11, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.J.; Park, J.; Lee, Y.S.; Chae, J.S.; Yu, D.H.; Park, B.K.; Kim, H.C.; Choi, K.S. Molecular identification of selected tick-borne pathogens in wild deer and raccoon dogs from the Republic of Korea. Veter. Parasitol. Reg. Stud. Rep. 2017, 7, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.G.; Chae, J.B.; Cho, Y.K.; Jo, Y.S.; Shin, N.S.; Lee, H.; Choi, K.S.; Yu, D.H.; Park, J.; Park, B.K.; et al. Molecular Detection of Anaplasma, Bartonella, and Borrelia theileri in Raccoon Dogs (Nyctereutes procyonoides) in Korea. Am. J. Trop. Med. Hyg. 2018, 98, 1061–1068. [Google Scholar] [CrossRef]
- DE Sousa, K.C.M.; Calchi, A.C.; Herrera, H.M.; Dumler, J.S.; Barros-Battesti, D.M.; Machado, R.Z.; André, M.R. Anaplasmataceae agents among wild mammals and ectoparasites in Brazil. Epidemiol. Infect. 2017, 145, 3424–3437. [Google Scholar] [CrossRef]
- Barlough, J.E.; Madigan, J.E.; DeRock, E.; Bigornia, L. Nested polymerase chain reaction for detection of Ehrlichia equi genomic DNA in horses and ticks (Ixodes pacifcus). Vet. Parasitol. 1996, 63, 319–329. [Google Scholar] [CrossRef]
- Kawahara, M.; Rikihisa, Y.; Lin, Q.; Isogai, E.; Tahara, K.; Itagaki, A.; Hiramitsu, Y.; Tajima, T. Novel genetic variants of Anaplasma phagocytophilum, Anaplasma bovis, Anaplasma centrale, and a novel Ehrlichia sp. in wild deer and ticks on two major islands in Japan. Appl. Environ. Microbiol. 2006, 72, 1102–1109. [Google Scholar] [CrossRef]
- Silva, C.B.D.; Santos, H.A.; Navarrete, M.G.; Ribeiro, C.C.D.U.; Gonzalez, B.C.; Zaldivar, M.F.; Pires, M.S.; Peckle, M.; Costa, R.L.D.; Vitari, G.L.V.; et al. Molecular detection and characterization of Anaplasma platys in dogs and ticks in Cuba. Ticks Tick Borne Dis. 2016, 7, 938–944. [Google Scholar] [CrossRef]
- Corales, J.M.; Viloria, V.V.; Venturina, V.M.; Mingala, C.N. The prevalence of Ehrlichia canis, Anaplasma platys and Babesia spp. in dogs in Nueva Ecija, Philippines based on multiplex polymerase chain reaction (mPCR) assay. Annals Parasitol. 2014, 60, 267–272. [Google Scholar]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of the genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unifification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and “HGE agent” as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar]
- Alhassan, A.; Hove, P.; Sharma, B.; Matthew-Belmar, V.; Karasek, I.; Lanza-Perea, M.; Werners, A.H.; Wilkerson, M.J.; Ganta, R.R. Molecular detection and characterization of Anaplasma platys and Ehrlichia canis in dogs from the Caribbean. Ticks Tick-Borne Dis. 2021, 12, 101727. [Google Scholar] [CrossRef] [PubMed]
- Soares, R.; Ramos, C.A.; Pedroso, T.; Babo-Terra, V.; Cleveland, H.; Araújo, F. Molecular survey of Anaplasma platys and Ehrlichia canis in dogs from Campo Grande, Mato Grosso do Sul, Brazil. An. Acad Bras Cienc. 2017, 89, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Almazán, C.; González-Álvarez, V.H.; Fernández de Mera, I.G.; Cabezas-Cruz, A.; Rodríguez-Martínez, R.; de la Fuente, J. Molecular identification and characterization of Anaplasma platys and Ehrlichia canis in dogs in Mexico. Ticks Tick Borne Dis. 2016, 7, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Diniz, P.P.; Beall, M.J.; Omark, K.; Chandrashekar, R.; Daniluk, D.A.; Cyr, K.E.; Koterski, J.F.; Robbins, R.G.; Lalo, P.G. High prevalence of tick-borne pathogens in dogs from an Indian reservation in northeastern Arizona. Vector Borne Zoon. Dis. 2010, 10, 117–123. [Google Scholar] [CrossRef]
- Cardoso, L.; Gilad, M.; E Cortes, H.C.; Nachum-Biala, Y.; Lopes, A.P.; Vila-Viçosa, M.J.; Simões, M.; Rodrigues, P.A.; Baneth, G. First report of Anaplasma platys infection in red foxes (Vulpes vulpes) and molecular detection of Ehrlichia canis and Leishmania infantum in foxes from Portugal. Parasit. Vectors. 2015, 8, 144. [Google Scholar] [CrossRef]
- Low, V.L.; Prakash, B.K.; Lim, Y.A.; Tan, T.K.; Vinnie-Siow, W.Y.; Sofian-Azirun, M.; AbuBakar, S. Detection of Anaplasmataceae agents and co-infection with other tick-borne protozoa in dogs and Rhipicephalus sanguineus sensu lato ticks. Exp. Appl. Acarol. 2018, 75, 429–435. [Google Scholar] [CrossRef]
- Manoj, R.R.S.; Iatta, R.; Latrofa, M.S.; Capozzi, L.; Raman, M.; Colella, V.; Otranto, D. Canine vector-borne pathogens from dogs and ticks from Tamil Nadu, India. Acta Trop. 2020, 203, 105308. [Google Scholar] [CrossRef]
- Li, H.-T.; Sun, L.-S.; Chen, Z.-M.; Hu, J.S.; Ye, C.D.; Jia, K.; Wang, H.; Yuan, L.G.; Zhang, G.H.; Li, S. Detection of Anaplasma platys in dogs using real-time loop-mediated isothermal amplification. Veter J. 2014, 199, 468–470. [Google Scholar] [CrossRef]
- Wang, S.; He, J.; Zhang, L. Serological investigation of vector-borne disease in dogs from rural areas of China. Asian Pac. J. Trop. Biomed. 2012, 2, 102–103. [Google Scholar] [CrossRef]
- Fukui, Y.; Inokuma, H. Subclinical Infections of Anaplasma phagocytophilum and Anaplasma bovis in Dogs from Ibaraki, Japan. Jpn. J. Infect. Dis. 2019, 72, 168–172. [Google Scholar] [CrossRef]
- Cui, Y.; Yan, Y.; Wang, X.; Cao, S.; Zhang, Y.; Jian, F.; Zhang, L.; Wang, R.; Shi, K.; Ning, C. First molecular evidence of mixed infections of Anaplasma species in dogs in Henan, China. Ticks Tick Borne Dis. 2017, 8, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, K.; Matsuyama, Y.; Matsuda, H.; Sakamoto, L.; Matsumoto, K.; Yokoyama, N.; Inokuma, H. Detection of Anaplasma bovis and Anaplasma phagocytophilum DNA from nymphs and larvae of Haemaphysalis megaspinosa in Hokkaido, Japan. Vet Parasitol. 2010, 168, 170–172. [Google Scholar] [CrossRef] [PubMed]
- Jilintai, S.N.; Hayakawa, D.; Suzuki, M.; Hata, H.; Kondo, S.; Matsumoto, K.; Yokoyama, N.; Inokuma, H. Molecular sursey for Anaplasma bovis and Anaplasma phagocytophilum infection in cattle in a pastureland where sika deer appear in Japan. Jpn. J. Infect Dis. 2009, 62, 73–75. [Google Scholar] [CrossRef]
- Arraga-Alvarado, C.M.; Qurollo, B.A.; Parra, O.C.; Berrueta, M.A.; Hegarty, B.C.; Breitschwerdt, E.B. Case report: Molecular evidence of Anaplasma platys infection in two women from Venezuela. Am. J. Trop. Med. Hyg. 2014, 91, 1161. [Google Scholar] [CrossRef] [PubMed]
- Kordick, S.K.; Breitschwerdt, E.B.; Hegarty, B.C.; Southwick, K.L.; Colitz, C.M.; Hancock, S.I.; Bradley, J.M.; Rumbough, R.; Mcpherson, J.T.; MacCormack, J.N. Coinfection with multiple tick-borne pathogens in a Walker hound kennel in North Carolina. J. Clin. Microbiol. 1999, 37, 2631–2638. [Google Scholar] [CrossRef]
- Yancey, C.B.; Diniz, P.P.V.P.; Breitschwerdt, E.B.; Hegarty, B.C.; Wiesen, C.; Qurollo, B.A. Doxycycline treatment efficacy in dogs with naturally occurring Anaplasma phagocytophilum infection. J. Small Anim. Pract. 2018, 59, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Ebani, V.V.; Bertelloni, F.; Torracca, B.; Cerri, D. Serological survey of Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, and Ehrlichia canis infections in rural and urban dogs in Central Italy. Ann. Agric. Environ. Med. 2014, 21, 671–675. [Google Scholar] [CrossRef]
- Elhamiani Khatat, S.; Khallaayoune, K.; Errafyk, N.; Van Gool, F.; Duchateau, L.; Daminet, S.; Kachani, M.; El Amri, H.; Azrib, R.; Sahibi, H. Detection of Anaplasma spp. and Ehrlichia spp. antibodies and Dirofifilaria immitis antigens in dogs from seven locations of Morocco. Vet. Parasitol. 2017, 239, 86–89. [Google Scholar] [CrossRef]
- Nyarko, E.; Grab, D.J.; Dumler, J.S. Anaplasma phagocytophilum-infected neutrophils enhance transmigration of Borrelia burgdorferi across the human blood brain barrier In Vitro. Int. J. Parasitol. 2006, 36, 601–605. [Google Scholar] [CrossRef]
- Nieto, N.C.; Foley, J.E. Meta-analysis of coinfection and coexposure with Borrelia burgdorferi and Anaplasma phagocytophilum in humans, domestic animals, wildlife, and Ixodes ricinus-complex ticks. Vector Borne Zoonotic Dis. 2009, 9, 93–102. [Google Scholar] [CrossRef]
- Holden, K.; Hodzic, E.; Feng, S.; Freet, K.J.; Lefebvre, R.B.; Barthold, S.W. Coinfection with Anaplasma phagocytophilum alters Borrelia burgdorferi population distribution in C3H/HeN mice. Infect. Immun. 2005, 73, 3440–3444. [Google Scholar] [CrossRef] [PubMed]
- Beall, M.J.; Chandrashekar, R.; Eberts, M.D.; Cyr, K.E.; Diniz, P.P.; Mainville, C.; Hegarty, B.C.; Crawford, J.M.; Breitschwerdt, E.B. Serological and molecular prevalence of Borrelia burgdorferi, Anaplasma phagocytophilum, and Ehrlichia species in dogs from Minnesota. Vector Borne Zoonotic Dis. 2008, 8, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Atif, F.A. Alpha proteobacteria of genus Anaplasma (Rickettsiales: Anaplasmataceae): Epidemiology and characteristics of Anaplasma species related to veterinary and public health importance. Parasitology 2016, 143, 659–685. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yan, M.; Liu, A.; Chen, T.; Luo, L.; Li, L.; Teng, Z.; Li, B.; Ji, Z.; Jian, M.; et al. The seroprevalence of Anaplasma phagocytophilum in global human populations: A systematic review and meta-analysis. Transbound. Emerg. Dis. 2020, 67, 2050–2064. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, Y.; Wang, W.; Guo, D.; Huang, S.; Jie, S. The clinical characteristics and outcomes of patients with human granulocytic anaplasmosis in China. Int. J. Infect. Dis. 2011, 15, e859–e866. [Google Scholar] [CrossRef] [PubMed]
- Niaz, S.; Ur Rahman, Z.; Ali, I.; Cossío-Bayúgar, R.; Amaro-Estrada, I.; Alanazi, A.D.; Khattak, I.; Zeb, J.; Nasreen, N.; Khan, A. Molecular prevalence, characterization and associated risk factors of Anaplasma spp. and Theileria spp. in small ruminants in Northern Pakistan. Parasite 2021, 28, 3. [Google Scholar] [CrossRef]
- Galay, R.L.; Manalo, A.A.L.; Dolores, S.L.D.; Aguilar, I.P.M.; Sandalo, K.A.C.; Cruz, K.B.; Divina, B.P.; Andoh, M.; Masatani, T.; Tanaka, T. Molecular detection of tick-borne pathogens in canine population and Rhipicephalus sanguineus (sensu lato) ticks from southern Metro Manila and Laguna, Philippines. Parasit Vectors 2018, 11, 643. [Google Scholar] [CrossRef]
- Chatanga, E.; Kainga, H.; Razemba, T.; Ssuna, R.; Swennen, L.; Hayashida, K.; Sugimoto, C.; Katakura, K.; Nonaka, N.; Nakao, R. Molecular detection and characterization of tick-borne hemoparasites and Anaplasmataceae in dogs in major cities of Malawi. Parasitol. Res. 2021, 120, 267–276. [Google Scholar] [CrossRef]
- QQiu, Y.; Kaneko, C.; Kajihara, M.; Ngonda, S.; Simulundu, E.; Muleya, W.; Thu, M.J.; Hang’ombe, M.B.; Katakura, K.; Takada, A.; et al. Tick-borne hemoparasites and Anaplasmataceae in domestic dogs in Zambia. Ticks Tick Borne Dis. 2018, 9, 988–995. [Google Scholar] [CrossRef]
- Konto, M.; Biu, A.A.; Ahmed, M.I.; Charles, S. Prevalence and seasonal abundance of ticks on dogs and the role of Rhipicephalus sanguineus in transmitting Babesia species in Maidugiri, northeastern Nigeria. Vet. World 2014, 7, 119–124. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Location | Number of Samples | Location | Number of Samples |
|---|---|---|---|
| Haikou | 636 | Lingshui | 4 |
| Qiongzhong | 19 | Baisha | 14 |
| Dongfang | 2 | Chenmai | 16 |
| Wenchang | 31 | Baoting | 24 |
| Changjiang | 15 | Wanning | 18 |
| Danzhou | 36 | Ledong | 16 |
| Dingan | 107 | Tunchang | 30 |
| Wuzhishan | 12 | Lingao | 5 |
| Qionghai | 34 | Sanya | 32 |
| Target Gene | Primer | Sequence 5′ to 3′ | Amplicon Size (bp) | References |
|---|---|---|---|---|
| 16S rRNA | EE-1 | TCCTGGCTCAGAACGAACGCTGGCGGC | 1433 | Barlough et al. [17] |
| EE-2 | AGTCACTGACCCAACCTTAAATGGCTG | |||
| 16S rRNA | SSAP2f | GCTGAATGTGGGGATAATTTAT | 641 | Kawahara et al. [18] |
| SSAP2r | ATGGCTGCTTCCTTTCGGTTA | |||
| 16S rRNA | AB1f | CTCGTAGCTTGCTATGAGAAC | 551 | |
| AB1r | TCTCCCGGACTCCAGTCTG | |||
| gltA | gltAf | GACCTACGATCCGGGATTCA | 580 | Silva et al. [19] |
| gltAr | CCGCACGGTCGCTGTT |
| Category | Primer | Amplification Conditions | |||||
|---|---|---|---|---|---|---|---|
| A. bovis | EE1 EE2 | 94 °C | 94 °C | 55 °C | 72 °C | 35 | 72 °C |
| 5 min | 30 s | 30 s | 30 s | 5 min | |||
| AB1f AB1r | 94 °C | 94 °C | 58 °C | 72 °C | 40 | 72 °C | |
| 5 min | 30 s | 30 s | 30 s | 10 min | |||
| phagocytophilum | EE1 EE2 | 94 °C | 94 °C | 55 °C | 72 °C | 35 | 72 °C |
| 5 min | 30 s | 30 s | 30 s | 5 min | |||
| SSAP2f SSAP2r | 94 °C | 94 °C | 58 °C | 72 °C | 40 | 72 °C | |
| 5 min | 30 s | 30 s | 30 s | 10 min | |||
| A. platys | gltAF gltAR | 94 °C | 94 °C | 60 °C | 72 °C | 35 | 72 °C |
| 3 min | 1 min | 1 min | 1 min | 5 min | |||
| First | Second | ||
|---|---|---|---|
| Components | Volume (μL) | Components | Volume (μL) |
| 2 × Taq Plus Master Mix II (Dye Plus) | 12.5 | 2 × Taq Plus Master Mix II (Dye Plus) | 12.5 |
| Forward Primer | 0.5 | Forward Primer | 0.7 |
| Reverse Primer | 0.5 | Reverse Primer | 0.7 |
| Template | 1 | Template | 1 |
| ddH2O | 10.5 | ddH2O | 10.1 |
| Total | 25 | Total | 25 |
| Components | Volume (μL) |
|---|---|
| 2 × Taq Plus Master Mix II (Dye Plus) | 12.5 |
| Forward Primer | 0.5 |
| Reverse Primer | 0.5 |
| Template | 1 |
| ddH2O | 10.5 |
| Total | 25 |
| Location | No. Tested | Positive | No. Infected/(%) |
|---|---|---|---|
| Haikou | 636 | 16 | 2.5 |
| Qiongzhong | 19 | 12 | 63.2 |
| Dongfang | 2 | 0 | 0 |
| Wenchang | 31 | 7 | 22.6 |
| Changjing | 15 | 1 | 6.7 |
| Danzhou | 36 | 7 | 19.4 |
| Dingan | 107 | 12 | 11.2 |
| Wuzhishan | 12 | 0 | 0 |
| Qionghai | 34 | 5 | 14.7 |
| Lingshui | 4 | 1 | 25 |
| Baisha | 14 | 1 | 7.1 |
| Chengmai | 16 | 2 | 12.5 |
| Baoting | 24 | 6 | 25 |
| Wanning | 18 | 1 | 5.5 |
| Ledong | 16 | 7 | 43.8 |
| Tunchang | 30 | 16 | 53.3 |
| Lingao | 5 | 4 | 80 |
| Sanya | 32 | 1 | 3.1 |
| Total | 1051 | 99 | 9.4 |
| Location | No. Tested | No. Infected/(%) | ||
|---|---|---|---|---|
| A. phagocytophilum (%) | A. bovis (%) | A. platys (%) | ||
| Haikou | 636 | 0.15(1/636) | 2.20(14/636) | 0.15(1/636) |
| Qiongzhong | 19 | 0(0/19) | 0(0/19) | 63.16(12/19) |
| Dongfang | 2 | 0(0/2) | 0(0/2) | 0(0/2) |
| Wenchang | 31 | 0(0/31) | 16.13(5/31) | 6.45(2/31) |
| Changjing | 15 | 0(0/15) | 0(0/15) | 6.67(1/15) |
| Danzhou | 36 | 0(0/36) | 0(0/36) | 19.44(7/36) |
| Dingan | 107 | 5.61(6/107) | 1.87(2/107) | 5.61(6/107) |
| Wuzhishan | 12 | 0(0/12) | 0(0/12) | 0(0/12) |
| Qionghai | 34 | 0(0/34) | 0(0/34) | 14.71(5/34) |
| Lingshui | 4 | 0(0/4) | 0(0/4) | 25.0(1/4) |
| Baisha | 14 | 0(0/14) | 0(0/14) | 7.14(1/14) |
| Chengmai | 16 | 0(0/16) | 5.26(1/16) | 6.25(1/16) |
| Baoting | 24 | 12.50(3/24) | 4.17(1/24) | 12.50(3/24) |
| Wanning | 18 | 5.55(1/18) | 0(0/18) | 0(0/18) |
| Ledong | 16 | 0(0/16) | 0(0/16) | 43.75(7/16) |
| Tunchang | 30 | 0(0/30) | 0(0/30) | 53.33(16/30) |
| Lingao | 5 | 0(0/5) | 80.0(4/5) | (0/5) |
| Sanya | 32 | 0(0/32) | 3.13(1/32) | 0(0/32) |
| Total | 1051 | 1.0(11/1051) | 2.7(28/1051) | 6.0(63/1051) |
| Factor | Parametrs | Number | Infection Rate (%) | Statistical Analysis |
|---|---|---|---|---|
| Gender | Male | 547 | 11.15(61/547) | X2 = 4.011, df = 1, p = 0.045 |
| Female | 504 | 7.54(38/504) | ||
| Age | <1 year | 357 | 12.61(45/357) | X2 = 6.430, df = 1, p = 0.011 * |
| >1 year | 694 | 7.78(54/694) | ||
| Pesticide | Yes | 412 | 6.55(27/412 | X2 = 4.939, df = 1, p = 0.026 * |
| No | 639 | 11.27(72/639) | ||
| Tick infection | Yes | 56 | 35.71(20/56) | X2 = 47.933, df = 1, p < 0.0001 * |
| No | 995 | 7.94(79/995) | ||
| Feeding mode | urban dogs | 217 | 5.07(11/217) | X2 = 30.305, df = 2, p < 0.0001 * |
| dogs in shelters | 292 | 3.76(11/292) | ||
| rural dogs | 542 | 14.21(77/542) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.; Zhou, S.; Upadhyay, A.; Zhao, J.; Liao, C.; Guan, Q.; Wang, J.; Han, Q. Molecular Detection and Phylogenetic Characterization of Anaplasma spp. in Dogs from Hainan Province/Island, China. Vet. Sci. 2023, 10, 339. https://doi.org/10.3390/vetsci10050339
Lin Y, Zhou S, Upadhyay A, Zhao J, Liao C, Guan Q, Wang J, Han Q. Molecular Detection and Phylogenetic Characterization of Anaplasma spp. in Dogs from Hainan Province/Island, China. Veterinary Sciences. 2023; 10(5):339. https://doi.org/10.3390/vetsci10050339
Chicago/Turabian StyleLin, Yang, Sa Zhou, Archana Upadhyay, Jianguo Zhao, Chenghong Liao, Qingfeng Guan, Jinhua Wang, and Qian Han. 2023. "Molecular Detection and Phylogenetic Characterization of Anaplasma spp. in Dogs from Hainan Province/Island, China" Veterinary Sciences 10, no. 5: 339. https://doi.org/10.3390/vetsci10050339