
Genetic Variability and Recombination of the NSP2 Gene of PRRSV-2 Strains in China from 1996 to 2021
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. The Dataset
2.2. Phylogenetic Analysis
2.3. Alignment of NSP2 Nucleotide Sequences
2.4. Alignment of NSP2 Amino Acid Sequences
2.5. Recombination Analysis
3. Results
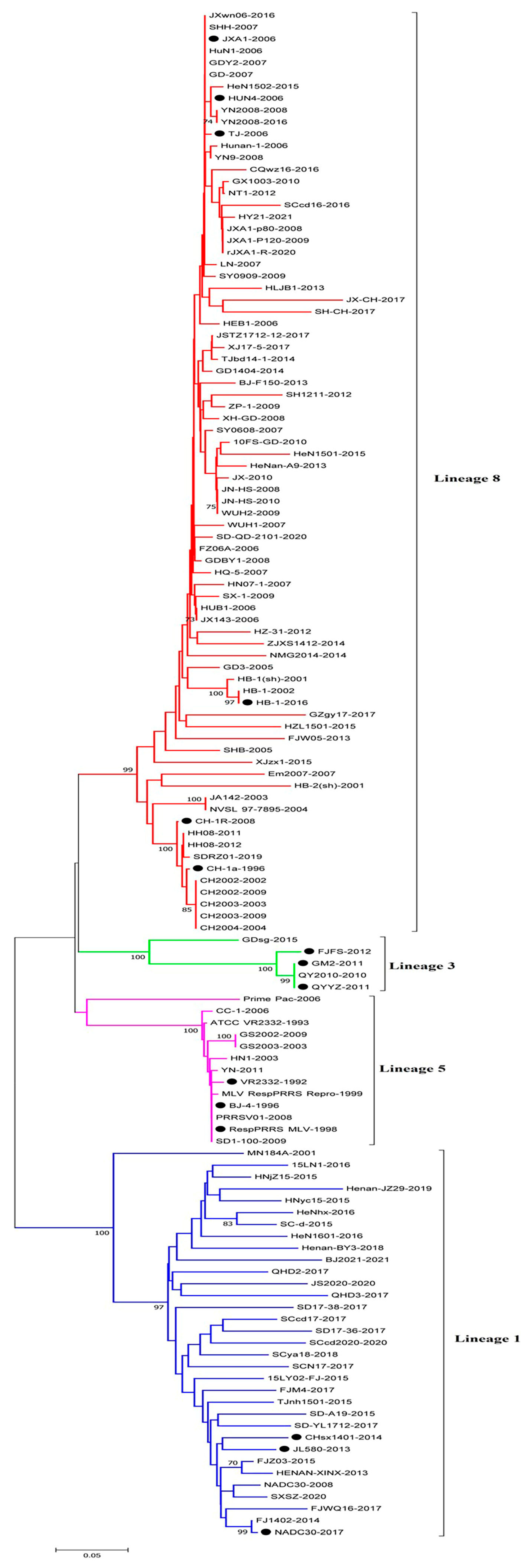
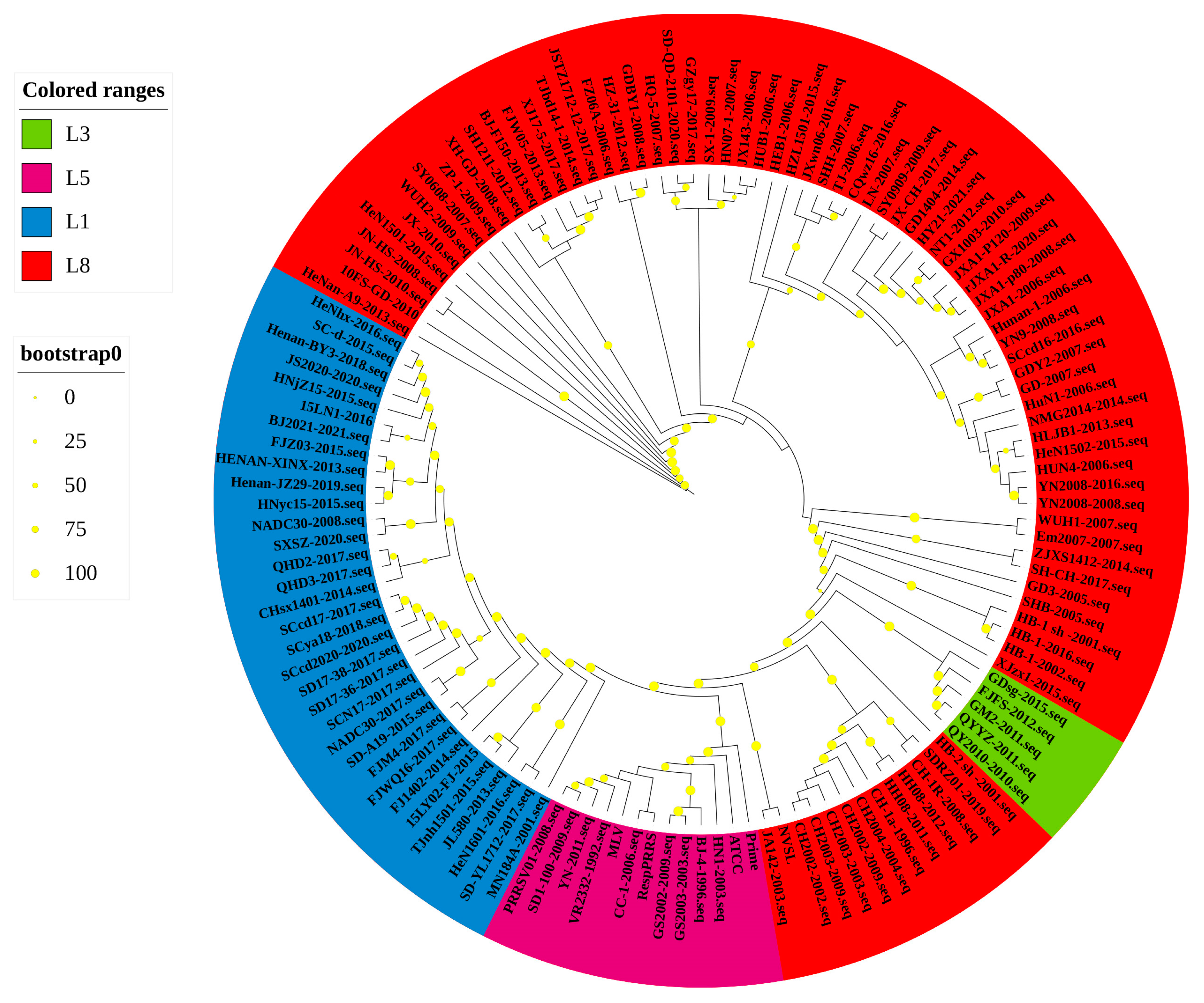
3.1. Phylogenetic Analysis
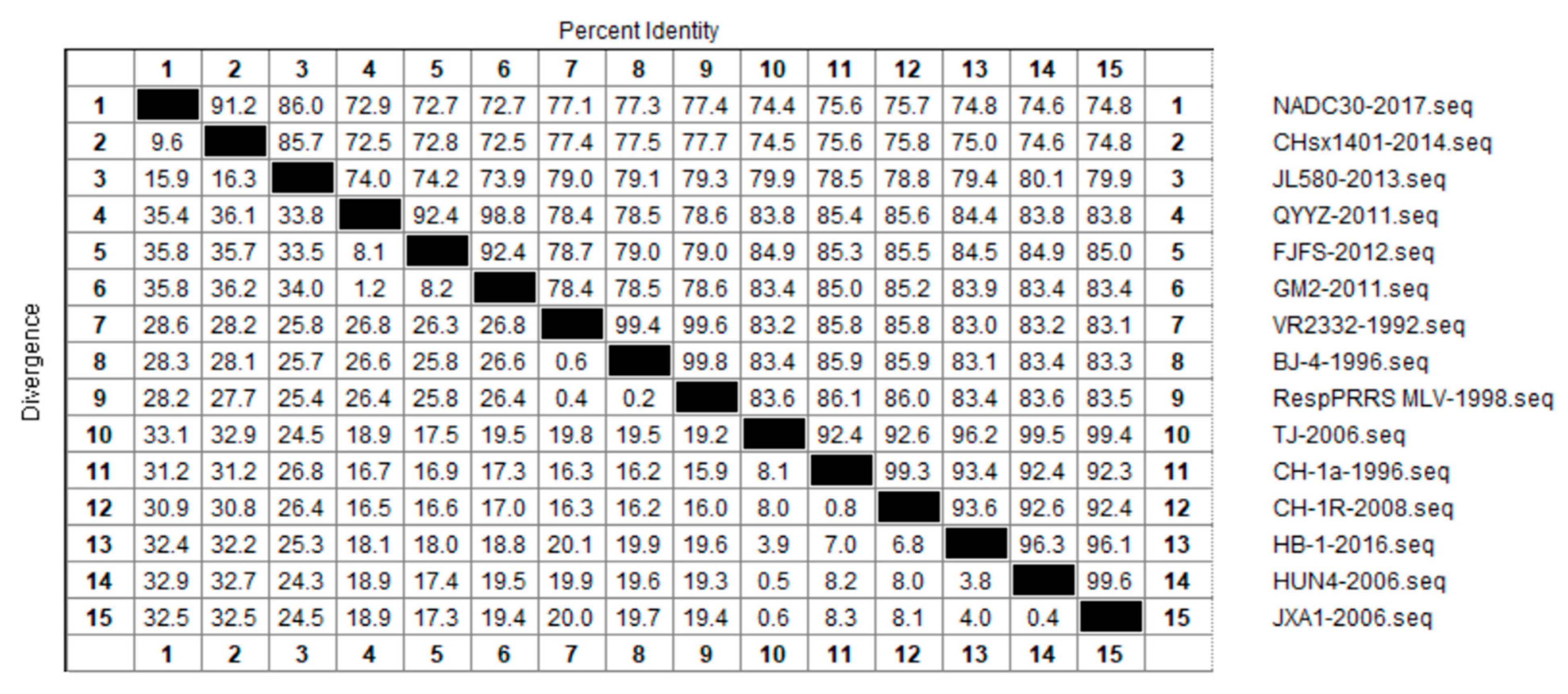
3.2. Nucleotide Similarity
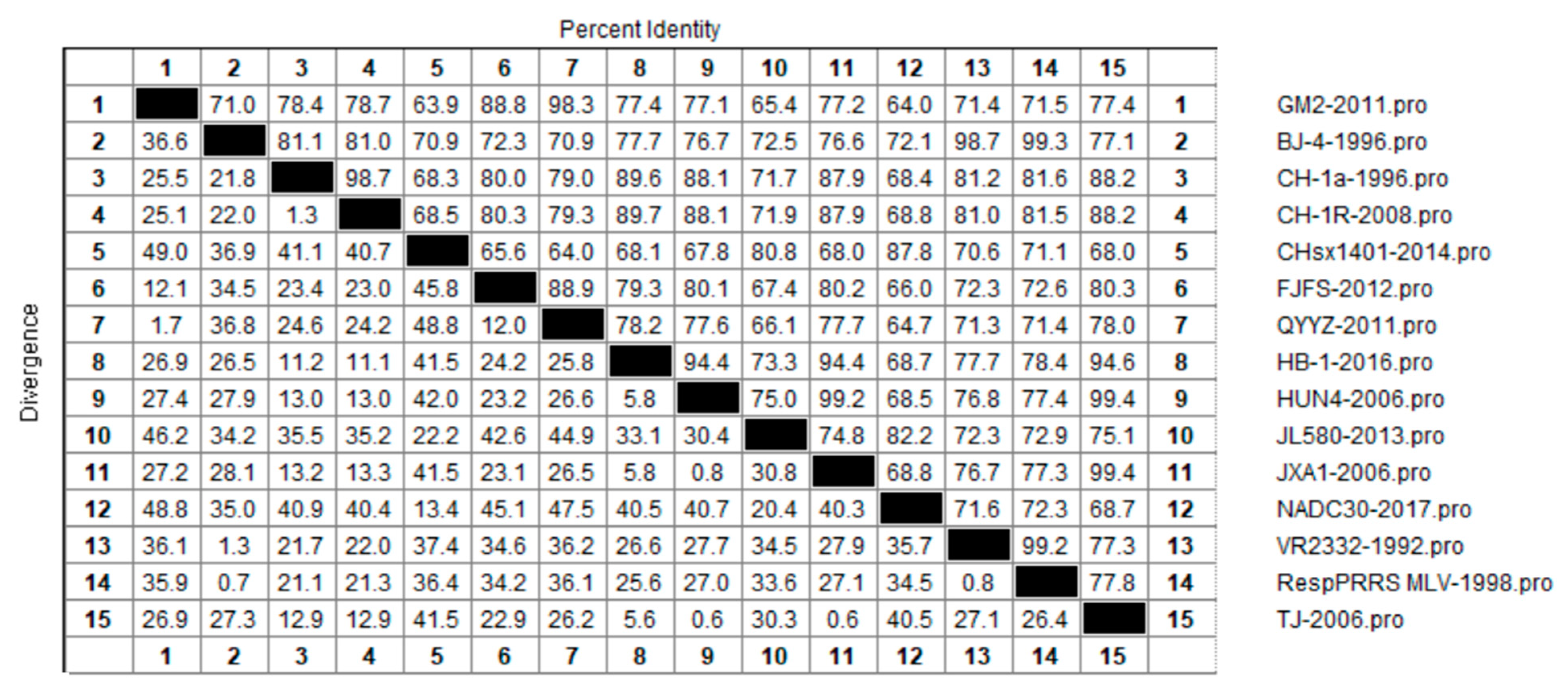
3.3. Amino Acid Sequence Similarity
3.4. Amino Acid Sequence Alignment
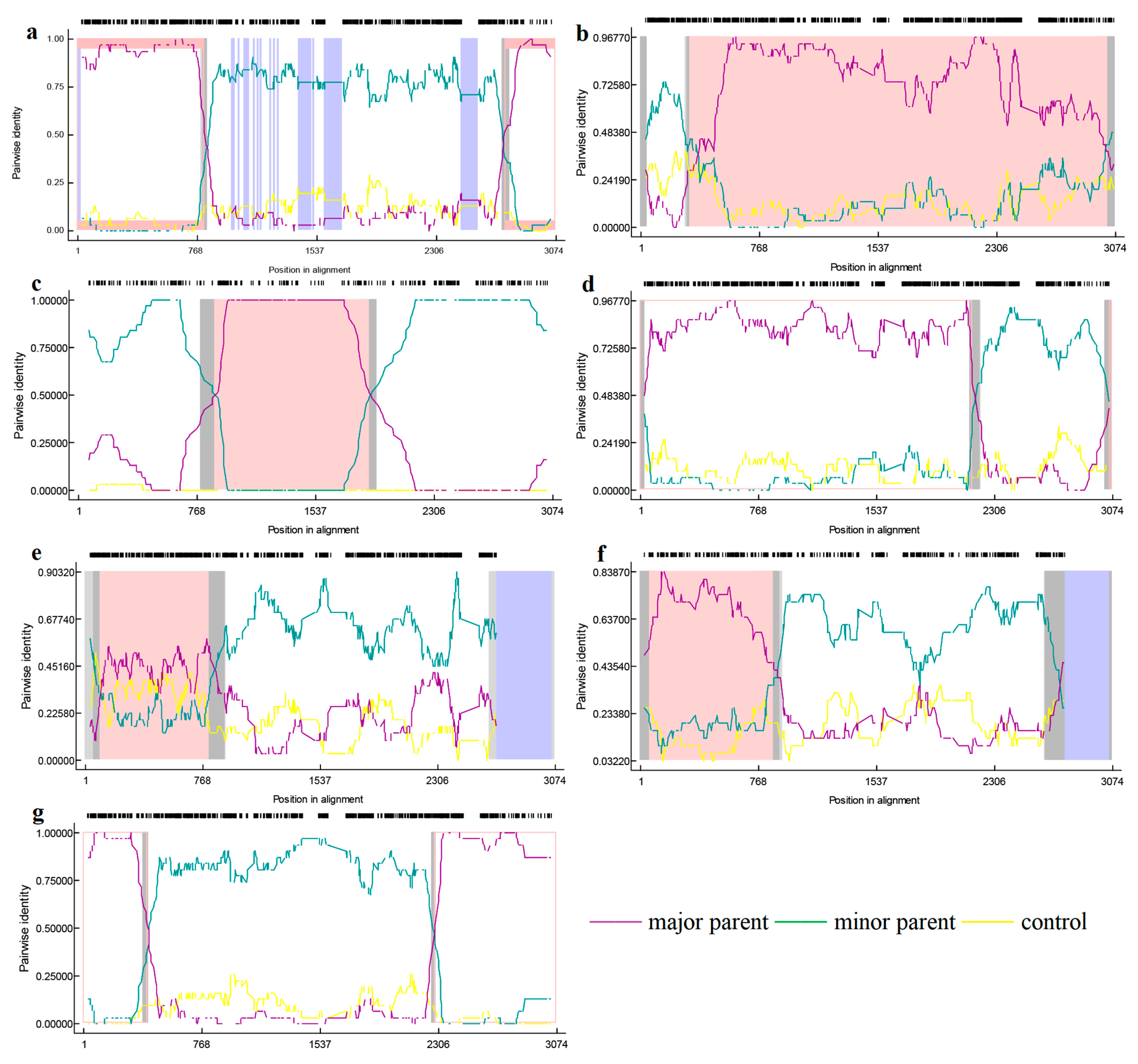
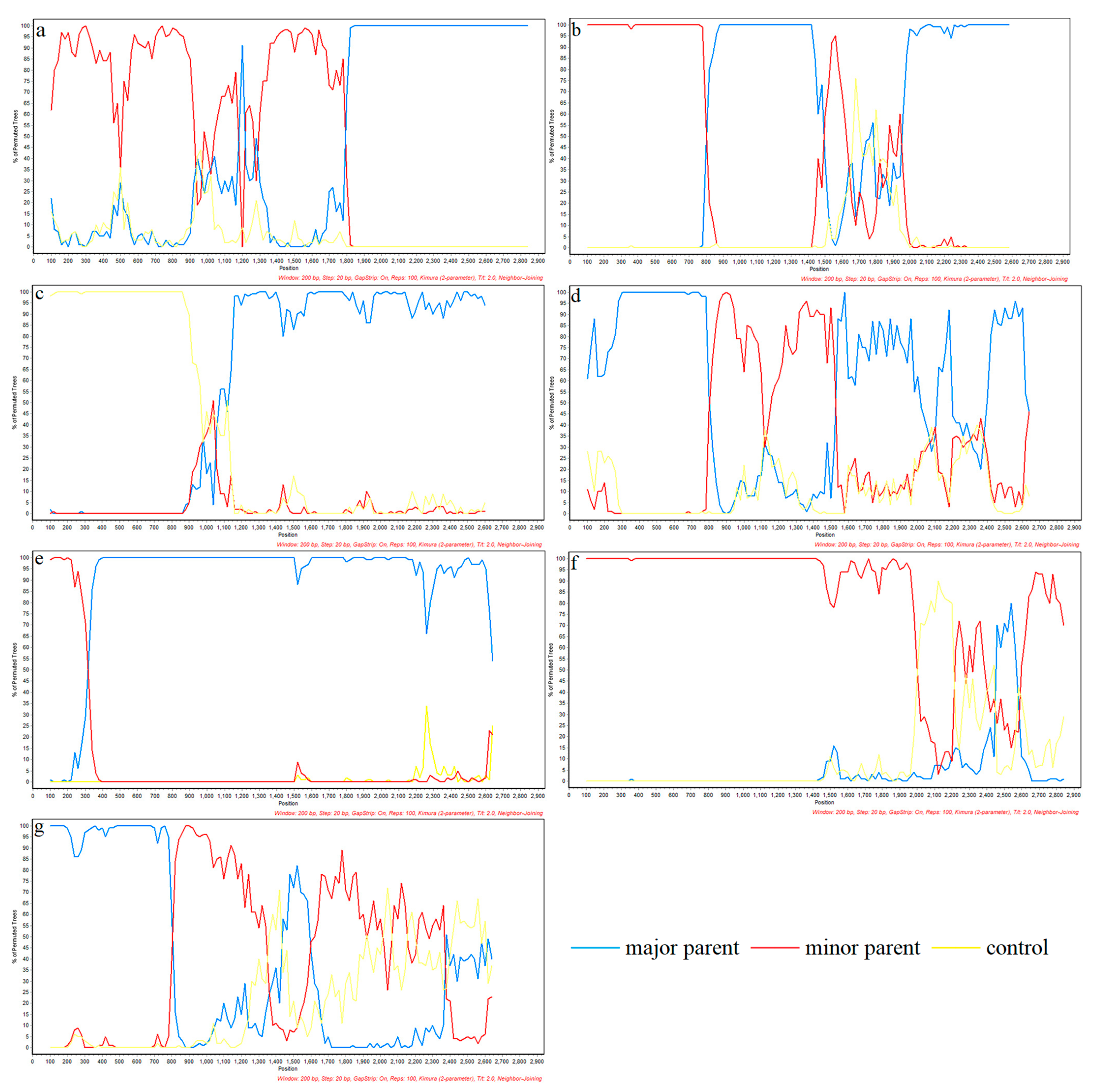
3.5. Recombinant Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Benfield, D.A.; Nelson, E.; Collins, J.E.; Harris, L.; Goyal, S.M.; Robison, D.; Christianson, W.T.; Morrison, R.B.; Gorcyca, D.; Chladek, D. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Investig. 1992, 4, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Snijder, E.J.; Meulenberg, J.J. The molecular biology of arteriviruses. J. Gen. Virol. 1998, 79, 961–979. [Google Scholar] [CrossRef] [PubMed]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef]
- Zhou, Z.; Ni, J.; Cao, Z.; Han, X.; Xia, Y.; Zi, Z.; Ning, K.; Liu, Q.; Cai, L.; Qiu, P.; et al. The epidemic status and genetic diversity of 14 highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) isolates from China in 2009. Vet. Microbiol. 2011, 150, 257–269. [Google Scholar] [CrossRef]
- Tian, K. NADC30-Like Porcine Reproductive and Respiratory Syndrome in China. Open Virol. J. 2017, 11, 59–65. [Google Scholar] [CrossRef]
- Li, X.; Wu, J.; Tan, F.; Li, Y.; Ji, G.; Zhuang, J.; Zhai, X.; Tian, K. Genome characterization of two NADC30-like porcine reproductive and respiratory syndrome viruses in China. Springerplus 2016, 5, 1677. [Google Scholar] [CrossRef]
- Fang, Y.; Treffers, E.E.; Li, Y.; Tas, A.; Sun, Z.; van der Meer, Y.; de Ru, A.H.; van Veelen, P.A.; Atkins, J.F.; Snijder, E.J.; et al. Efficient-2 frameshifting by mammalian ribosomes to synthesize an additional arterivirus protein. Proc. Natl. Acad. Sci. USA 2012, 109, 2920–2928. [Google Scholar] [CrossRef]
- Allende, R.; Lewis, T.L.; Lu, Z.; Rock, D.L.; Kutish, G.F.; Ali, A.; Doster, A.R.; Osorio, F.A. North American and European porcine reproductive and respiratory syndrome viruses differ in non-structural protein coding regions. J. Gen. Virol. 1999, 80, 307–315. [Google Scholar] [CrossRef]
- Johnson, C.R.; Griggs, T.F.; Gnanandarajah, J.; Murtaugh, M.P. Novel structural protein in porcine reproductive and respiratory syndrome virus encoded by an alternative ORF5 present in all arteriviruses. J. Gen. Virol. 2011, 92, 1107–1116. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, S.; Zhang, J.; Zeng, J.; Guo, X.; Ge, X.; Zhang, D.; Yang, H. Molecular variation analysis of porcine reproductive and respiratory syndrome virus in China. Virus Res. 2009, 145, 97–105. [Google Scholar] [CrossRef]
- Han, J.; Wang, Y.; Faaberg, K.S. Complete genome analysis of RFLP 184 isolates of porcine reproductive and respiratory syndrome virus. Virus Res. 2006, 122, 175–182. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, G.; Yu, L.; Li, L.; Zhang, Y.; Zhou, Y.; Tong, W.; Liu, C.; Gao, F.; Tong, G. Genetic Diversity of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) From 1996 to 2017 in China. Front. Microbiol. 2020, 11, 618. [Google Scholar] [CrossRef]
- Shi, M.; Lam, T.T.; Hon, C.C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.; Li, J.; Wong, L.T.; Yip, C.W.; Jiang, J.W.; et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, X.X.; Li, X.; Qiao, S.; Deng, R.; Zhang, G. Prevalence and genetic characteristics of porcine reproductive and respiratory syndrome virus in central China during 2016–2017: NADC30-like PRRSVs are predominant. Microb. Pathog. 2019, 135, 103657. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, B.; Xu, L.; Jin, H.; Ge, X.; Guo, X.; Han, J.; Yang, H. Efficacy evaluation of three modified-live virus vaccines against a strain of porcine reproductive and respiratory syndrome virus NADC30-like. Vet. Microbiol. 2017, 207, 108–116. [Google Scholar] [CrossRef]
- Deng, Y.; Pan, Y.; Wang, D.; Zhou, Q.; Bi, Y.; Chen, F.; Song, Y. Complete genome sequence of porcine reproductive and respiratory syndrome virus strain QY2010 reveals a novel subgroup emerging in China. J. Virol. 2012, 86, 7719–7720. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, X.; Zhai, J.; Li, B.; Wei, C.; Dai, A.; Yang, X.; Luo, M. Genetic diversity and evolutionary characteristics of type 2 porcine reproductive and respiratory syndrome virus in southeastern China from 2009 to 2014. Arch. Virol. 2017, 162, 2603–2615. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, X.X.; Li, R.; Qiao, S.; Zhang, G. The prevalent status and genetic diversity of porcine reproductive and respiratory syndrome virus in China: A molecular epidemiological perspective. Virol. J. 2018, 15, 2. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, Z.; Xu, T.; Zhou, Y.; Li, J.; Deng, H.; Li, F.; Xu, L.; Sun, X.; Zhu, L. Molecular Characterization of the Nsp2 and ORF5s of PRRSV Strains in Sichuan China during 2012–2020. Animals 2022, 12, 3309. [Google Scholar] [CrossRef]
- Yu, F.; Yan, Y.; Shi, M.; Liu, H.Z.; Zhang, H.L.; Yang, Y.B.; Huang, X.Y.; Gauger, P.C.; Zhang, J.; Zhang, Y.H.; et al. Phylogenetics, Genomic Recombination, and NSP2 Polymorphic Patterns of Porcine Reproductive and Respiratory Syndrome Virus in China and the United States in 2014–2018. J. Virol. 2020, 94, e01813-19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Kang, R.; Ji, G.; Tian, Y.; Ge, M.; Xie, B.; Yang, X.; Wang, H. Molecular characterization and recombination analysis of porcine reproductive and respiratory syndrome virus emerged in southwestern China during 2012–2016. Virus Genes 2018, 54, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Yin, B.; Qi, S.; Sha, W.; Qin, H.; Liu, L.; Yun, J.; Zhu, J.; Li, G.; Sun, D. Molecular Characterization of the Nsp2 and ORF5 (ORF5a) Genes of PRRSV Strains in Nine Provinces of China During 2016–2018. Front. Vet. Sci. 2021, 8, 605832. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Gao, J.C.; Xiong, J.Y.; Guo, J.C.; Yang, Y.B.; Jiang, C.G.; Tang, Y.D.; Tian, Z.J.; Cai, X.H.; Tong, G.Z.; et al. Two Residues in NSP9 Contribute to the Enhanced Replication and Pathogenicity of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2018, 92, e02209-17. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Ye, M.; Li, S.; Huang, Y.; Zhou, R.; Yu, X.; Tian, K.; Zhu, J. Emergence of a novel highly pathogenic recombinant virus from three lineages of porcine reproductive and respiratory syndrome virus 2 in China 2017. Transbound. Emerg. Dis. 2018, 65, 1775–1785. [Google Scholar] [CrossRef]
- Bian, T.; Sun, Y.; Hao, M.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. A recombinant type 2 porcine reproductive and respiratory syndrome virus between NADC30-like and a MLV-like: Genetic characterization and pathogenicity for piglets. Infect. Genet. Evol. 2017, 54, 279–286. [Google Scholar] [CrossRef]
- Shi, M.; Holmes, E.C.; Brar, M.S.; Leung, F.C. Recombination is associated with an outbreak of novel highly pathogenic porcine reproductive and respiratory syndrome viruses in China. J. Virol. 2013, 87, 10904–10907. [Google Scholar] [CrossRef]
- Zhao, H.; Han, Q.; Zhang, L.; Zhang, Z.; Wu, Y.; Shen, H.; Jiang, P. Emergence of mosaic recombinant strains potentially associated with vaccine JXA1-R and predominant circulating strains of porcine reproductive and respiratory syndrome virus in different provinces of China. Virol. J. 2017, 14, 67. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, X.; Tian, Y.; Yin, S.; Geng, G.; Ge, X.; Guo, X.; Yang, H. Genetic diversity analysis of genotype 2 porcine reproductive and respiratory syndrome viruses emerging in recent years in China. Biomed. Res. Int. 2014, 2014, 748068. [Google Scholar] [CrossRef]
- Zhao, K.; Ye, C.; Chang, X.B.; Jiang, C.G.; Wang, S.J.; Cai, X.H.; Tong, G.Z.; Tian, Z.J.; Shi, M.; An, T.Q. Importation and Recombination Are Responsible for the Latest Emergence of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus in China. J. Virol. 2015, 89, 10712–10716. [Google Scholar] [CrossRef]
- Chen, P.; Tan, X.; Lao, M.; Wu, X.; Zhao, X.; Zhou, S.; Yu, J.; Zhu, J.; Yu, L.; Tong, W.; et al. The Novel PRRSV Strain HBap4-2018 with a Unique Recombinant Pattern Is Highly Pathogenic to Piglets. Virol. Sin. 2021, 36, 1611–1625. [Google Scholar] [CrossRef]
- Li, Y.; Ji, G.; Wang, J.; Tan, F.; Zhuang, J.; Li, X.; Tian, K. Complete Genome Sequence of an NADC30-Like Porcine Reproductive and Respiratory Syndrome Virus Characterized by Recombination with Other Strains. Genome Announc. 2016, 4, e00330-16. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Aera | Sequence | Strain | Genbank Accession Number | Based on the ORF5 Lineage |
|---|---|---|---|---|---|
| 1996 | China | NSP2 | CH-1a | AY032626 | 8 |
| 1996 | China | NSP2 | BJ-4 | AF331831 | 5 |
| 2001 | China | NSP2 | HB-1(sh) | AY150312 | 8 |
| 2001 | China | NSP2 | HB-2(sh) | AY262352 | 8 |
| 2002 | China | NSP2 | CH2002 | EU880438.2 | 8 |
| 2002 | China | NSP2 | HB-1 | HQ233605 | 8 |
| 2003 | China | NSP2 | HN1 | AY457635.1 | 5 |
| 2003 | China | NSP2 | CH2003 | EU880440.2 | 8 |
| 2003 | China | NSP2 | GS2003 | EU880442 | 5 |
| 2004 | China | NSP2 | NVSL 97-7895 | AY545985 | 8 |
| 2004 | China | NSP2 | CH2004 | EU880439.2 | 8 |
| 2005 | China | NSP2 | GD3 | GU269541 | 8 |
| 2005 | China | NSP2 | SHB | EU864232.1 | 8 |
| 2006 | China | NSP2 | Ingelvac ATP | DQ988080 | 8 |
| 2006 | China | NSP2 | Prime Pac | DQ779791 | 8 |
| 2006 | China | NSP2 | HEB1 | EF112447 | 8 |
| 2006 | China | NSP2 | CC-1 | EF153486 | 8 |
| 2006 | China | NSP2 | FZ06A | MF370557 | 8 |
| 2006 | China | NSP2 | JX143 | EU708726 | 8 |
| 2006 | China | NSP2 | TJ | EU860248 | 8 |
| 2006 | China | NSP2 | HUB1 | EF075945 | 8 |
| 2006 | China | NSP2 | Hunan-1 | EU200965.1 | 8 |
| 2006 | China | NSP2 | JXA1 | EF112445 | 8 |
| 2006 | China | NSP2 | HuN1 | EF198112.1 | 8 |
| 2006 | China | NSP2 | HUN4 | EF635006.1 | 8 |
| 2007 | China | NSP2 | Em2007 | EU262603 | 8 |
| 2007 | China | NSP2 | SHH | EU106888 | 8 |
| 2007 | China | NSP2 | LN | EU109502 | 8 |
| 2007 | China | NSP2 | HN07-1 | KX766378 | 8 |
| 2007 | China | NSP2 | SY0608 | EU144079 | 8 |
| 2007 | China | NSP2 | GD | EU825724.1 | 8 |
| 2007 | China | NSP2 | HQ-5 | EU255920.1 | 8 |
| 2007 | China | NSP2 | GDY2/2007 | EU200964.1 | 8 |
| 2007 | China | NSP2 | WUH1 | EU187484 | 8 |
| 2008 | China | NSP2 | XH-GD | EU624117 | 8 |
| 2008 | China | NSP2 | YN2008 | EU880435 | 8 |
| 2008 | China | NSP2 | JN-HS | HM016158 | 8 |
| 2008 | China | NSP2 | YN9 | GU232738 | 8 |
| 2008 | China | NSP2 | JXA1-p80 | FJ548853 | 8 |
| 2008 | China | NSP2 | GDBY1 | GQ374442 | 8 |
| 2008 | China | NSP2 | CH-1R | EU807840 | 8 |
| 2008 | China | NSP2 | PRRSV01 | FJ175687 | 5 |
| 2009 | China | NSP2 | SY0909 | HQ315837 | 8 |
| 2009 | China | NSP2 | ZP-1 | HM016159 | 8 |
| 2009 | China | NSP2 | SX-1 | GQ857656 | 8 |
| 2009 | China | NSP2 | WUH2 | EU678352 | 8 |
| 2009 | China | NSP2 | CH2003 | EU880440 | 8 |
| 2009 | China | NSP2 | JXA1-P120 | KC422727 | 8 |
| 2009 | China | NSP2 | SD1-100 | GQ914997 | 5 |
| 2009 | China | NSP2 | GS2002 | EU880441 | 5 |
| 2009 | China | NSP2 | CH2002 | EU880438 | 8 |
| 2010 | China | NSP2 | JN-HS | HM016158 | 8 |
| 2010 | China | NSP2 | 10FS-GD | JX192634 | 8 |
| 2010 | China | NSP2 | JX | JX317649 | 8 |
| 2010 | China | NSP2 | QY2010 | JQ743666 | 3 |
| 2010 | China | NSP2 | GX1003 | JX912249 | 8 |
| 2011 | China | NSP2 | YN | JX857698 | 5 |
| 2011 | China | NSP2 | GM2 | JN662424 | 3 |
| 2011 | China | NSP2 | QYYZ | JQ308798 | 3 |
| 2011 | China | NSP2 | HH08 | JX679179 | 8 |
| 2012 | China | NSP2 | FJFS | KP998476 | 3 |
| 2012 | China | NSP2 | HZ-31 | KC445138 | 5 |
| 2012 | China | NSP2 | SH1211 | KF678434 | 8 |
| 2012 | China | NSP2 | NT1 | KP179402 | 8 |
| 2013 | China | NSP2 | BJ-F150 | KP890342 | 8 |
| 2013 | China | NSP2 | HeNan-A9 | KJ546412 | 8 |
| 2013 | China | NSP2 | HENAN-XINX | KF611905 | 1 |
| 2013 | China | NSP2 | HLJB1 | KT351740 | 8 |
| 2013 | China | NSP2 | JL580 | KR706343.1 | 1 |
| 2013 | China | NSP2 | FJW05 | KP860911 | 1 |
| 2014 | China | NSP2 | NMG2014 | KM000066 | 8 |
| 2014 | China | NSP2 | GD1404 | MF669720 | 8 |
| 2014 | China | NSP2 | ZJXS1412 | MF669722 | 8 |
| 2014 | China | NSP2 | CHsx1401 | KP861625 | 1 |
| 2014 | China | NSP2 | TJbd14-1 | KP742986 | 8 |
| 2014 | China | NSP2 | FJ1402 | KX169191 | 1 |
| 2015 | China | NSP2 | XJzx1 | KX689233 | 8 |
| 2015 | China | NSP2 | HNyc15 | KT945018 | 3 |
| 2015 | China | NSP2 | SC-d | MF375261 | 8 |
| 2015 | China | NSP2 | HeN1501 | MF766472 | 8 |
| 2015 | China | NSP2 | HeN1502 | MF766473 | 8 |
| 2015 | China | NSP2 | SD-A19 | MF375260 | 8 |
| 2015 | China | NSP2 | HZL1501 | MF669721 | 8 |
| 2015 | China | NSP2 | TJnh1501 | KX510269 | 8 |
| 2015 | China | NSP2 | GDsg | KX621003 | 3 |
| 2015 | China | NSP2 | 15LY02-FJ | KU215417 | 8 |
| 2015 | China | NSP2 | HNjZ15 | KT945017 | 1 |
| 2015 | China | NSP2 | FJZ03 | KP860909 | 1 |
| 2016 | China | NSP2 | YN2008 | EU880435 | 8 |
| 2016 | China | NSP2 | HeNhx | KX766379 | 1 |
| 2016 | China | NSP2 | HeN1601 | MF766474 | 8 |
| 2016 | China | NSP2 | CQwz16 | KY620008.1 | 8 |
| 2016 | China | NSP2 | SCcd16 | KY620011.1 | 8 |
| 2016 | China | NSP2 | HB-1 | AH015834.2 | 8 |
| 2016 | China | NSP2 | JXwn06 | EF641008 | 8 |
| 2016 | China | NSP2 | 15LN1 | KX815423 | 8 |
| 2017 | China | NSP2 | SD-YL1712 | MT708500 | 8 |
| 2017 | China | NSP2 | SD17-36 | MH121061 | 8 |
| 2017 | China | NSP2 | SD17-38 | MH068878 | 8 |
| 2017 | China | NSP2 | QHD2 | MH167387 | 8 |
| 2017 | China | NSP2 | JSTZ1712-12 | MK906026 | 8 |
| 2017 | China | NSP2 | QHD3 | MH167388 | 8 |
| 2017 | China | NSP2 | XJ17-5 | MK759853 | 8 |
| 2017 | China | NSP2 | SCN17 | MH078490 | 8 |
| 2017 | China | NSP2 | SCcd17 | MG914067 | 8 |
| 2017 | China | NSP2 | GZgy17 | MK144542 | 8 |
| 2017 | China | NSP2 | NADC30 | MH500776.1 | 1 |
| 2017 | China | NSP2 | FJWQ16 | KX758249 | 8 |
| 2017 | China | NSP2 | FJM4 | KY412888 | 8 |
| 2017 | China | NSP2 | JX-CH | KY495780 | 8 |
| 2017 | China | NSP2 | SH-CH | KY495781 | 8 |
| 2018 | China | NSP2 | SCya18 | MK144543 | 8 |
| 2018 | China | NSP2 | Henan-BY3 | MN550954.1 | 8 |
| 2019 | China | NSP2 | Henan-JZ29 | MN550957.1 | 8 |
| 2019 | China | NSP2 | SDRZ01 | MZ322956 | 8 |
| 2020 | China | NSP2 | SD-QD-2101 | MZ172971 | 8 |
| 2020 | China | NSP2 | SXSZ-2020 | MW880772 | 8 |
| 2020 | China | NSP2 | JS2020 | MZ342900 | 8 |
| 2020 | China | NSP2 | rJXA1-R | MT163314 | 8 |
| 2020 | China | NSP2 | SCcd2020 | MW803134 | 8 |
| 2021 | China | NSP2 | BJ2021 | OK095299 | 8 |
| 2021 | China | NSP2 | HY21 | OL687155.1 | 8 |
| Year | Aera | Sequence | Strain | Genbank Accession Number | Based on the ORF5 Lineage |
|---|---|---|---|---|---|
| 1992 | USA | NSP2 | VR2332 | EF536003.1 | 5 |
| 1993 | USA | NSP2 | ATCC VR2332 | U87392.3 | 5 |
| 1996 | USA | NSP2 | SDSU73 | JN654458.1 | 8 |
| 2003 | USA | NSP2 | JA142 | AY424271.1 | 8 |
| 1998 | USA | NSP2 | RespPRRS MLV | AF066183.4 | 5 |
| 1999 | USA | NSP2 | Ingelvac ATP | DQ988080.1 | 8 |
| 1999 | USA | NSP2 | MLV RespPRRS/Repro | AF159149 | 8 |
| 2001 | USA | NSP2 | MN184A | DQ176019 | 1 |
| 2002 | USA | NSP2 | P129 | AF494042.1 | 8 |
| 2004 | USA | NSP2 | NVSL 97-7895 | AY545985.1 | 8 |
| 2006 | USA | NSP2 | Prime Pac | DQ779791.1 | 8 |
| 2008 | USA | NSP2 | NADC30 | JN654459 | 1 |
| 2008 | USA | NSP2 | NADC31 | JN660150 | 1 |
| Recombination Event | Recombinant Strains (Lineages) | Main Parental Strain (Lineages) | Minor Parental Strain (Lineages) | Recombinant Breakpoint | Recombination Analysis Method |
|---|---|---|---|---|---|
| 1 | CH-1R-2008 (8) | GD3-2005 (8) | CH-1a-1996 (8) | 63~2935 (1884~1892) | RDP (P = 2.349 × 10−149) GENECONV (P = 8.169 × 10−145) BootScan (P = 1.298 × 10−113) MaxChi (P = 1.507 × 10−46) Chimaera (P = 3.911 × 10−48) SiScan (P = 3.619 × 10−63) 3seq (P = 3.334 × 10−8) |
| 2 | SD-YL1712-2017 (8) | BJ2021-2021 (8) | JSTZ1712-12-2017 (8) | 7~2485 (784~823) | RDP(P = 1.915 × 10−64) GENECONV (P = 6.699 × 10−96) BootScan (P = 4.816 × 10−98) MaxChi (P = 5.325 × 10−27) Chimaera (P = 1.423 × 10−25) SiScan (P = 3.001 × 10−31) 3seq (P = 3.726 × 10−9) |
| 3 | MN184A-2001 (1) | NADC30-2008 (8) | QHD3-2017 (8) | 63~2519 (1027~1061) | RDP (P = 5.700 × 10−72) GENECONV (P = 1.524 × 10−45) BootScan (P = 8.197 × 10−71) MaxChi (P = 3.496 × 10−21) Chimaera (P = 6.284×10−22) SiScan (P = 1.075 × 10−29) 3seq (P = 5.226 × 10−55) |
| 4 | NADC30-2017 (8) | SD-A19-2015 (8) | FJ1402-2014 (1) | 772~889 (1414~1520) | RDP (1.308 × 10−31) GENECONV (P = 2.888 × 10−27) BootScan (P = 9.744 × 10−32) MaxChi (P = 4.598 × 10−16) Chimaera (P = 4.563 × 10−16) SiScan (P = 5.757 × 10−19) 3seq (P = 2.526 × 10−43) |
| 5 | SCya18-2018 (8) | SD17-36-2017 (8) | ZJXS1412-2014 (8) | 17~2539 (352~482) | RDP (P = 1.874 × 10−30) GENECONV (P = 6.122 × 10−8) BootScan (P = 1.347 × 10−29) MaxChi (P = 7.742 × 10−10) Chimaera (P = 7.922 × 10−12) SiScan (P = 1.804 × 10−10) 3seq (P = 1.276 × 10−10) |
| 6 | Ingelvac-ATP-1999 (8) | CC-1-2006 (8) | HB-2(sh)-2001 (8) | 33~2894 (907~1536) | RDP (P = 5.359 × 10−10) GENECONV(NS) BootScan (P = 7.970 × 10−12) MaxChi (P = 2.038 × 10−14) Chimaera (P = 2.298 × 10−10) SiScan (P = 5.881 × 10−6) 3seq (P = 1.257 × 10−8) |
| 7 | SCcd2020-2020 (8) | JS2020-2020 (8) | SCcd17-2017 (8) | 762~903 (2112~2255) | RDP (P = 2.347 × 10−4) GENECONV(NS) BootScan (P = 3.109 × 10−3) MaxChi (P = 1.818 × 10−7) Chimaera(NS) SiScan (P = 4.693 × 10−16) 3seq (P = 2.094 × 10−13) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Luo, Q.; Zheng, Y.; Sha, H.; Li, G.; Kong, W.; Huang, L.; Zhao, M. Genetic Variability and Recombination of the NSP2 Gene of PRRSV-2 Strains in China from 1996 to 2021. Vet. Sci. 2023, 10, 325. https://doi.org/10.3390/vetsci10050325
Zhang H, Luo Q, Zheng Y, Sha H, Li G, Kong W, Huang L, Zhao M. Genetic Variability and Recombination of the NSP2 Gene of PRRSV-2 Strains in China from 1996 to 2021. Veterinary Sciences. 2023; 10(5):325. https://doi.org/10.3390/vetsci10050325
Chicago/Turabian StyleZhang, Hang, Qin Luo, Yajie Zheng, Huiyang Sha, Gan Li, Weili Kong, Liangzong Huang, and Mengmeng Zhao. 2023. "Genetic Variability and Recombination of the NSP2 Gene of PRRSV-2 Strains in China from 1996 to 2021" Veterinary Sciences 10, no. 5: 325. https://doi.org/10.3390/vetsci10050325