
A Calcium- and GTP-Dependent Transglutaminase in Leishmania infantum

 , ,
, ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Parasites
2.3. Preparation of Lysates
2.4. In Vivo Detection of TGase Activity
2.5. In Vitro in-Gel Detection of TGase Activity
2.6. In Vitro Micro-Well Plate Detection of TGase Activity
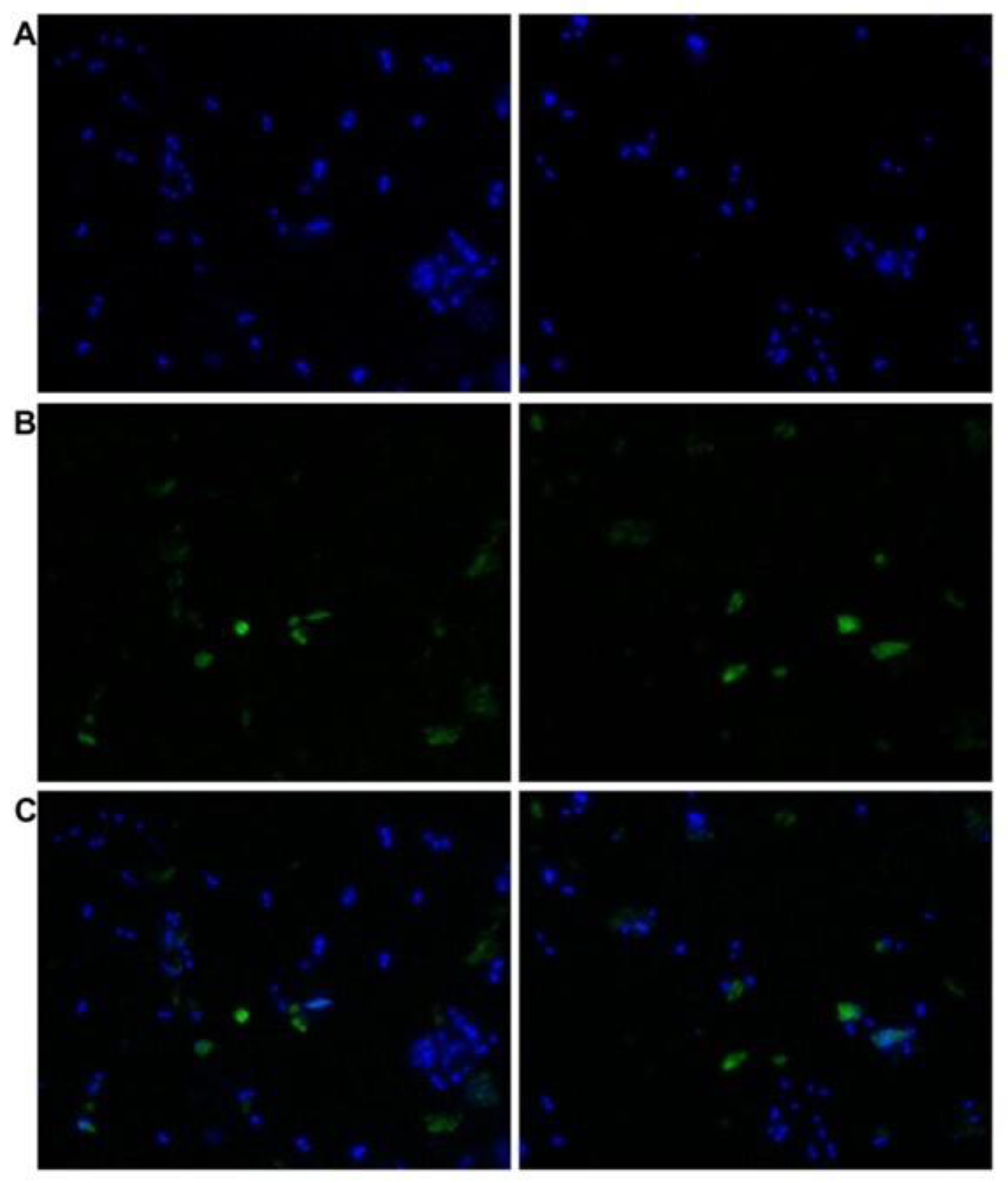
2.7. Immunocytochemical Staining
2.8. TGase Purification
2.9. Immunoblotting of Promastigote Lysates
2.10. Statistical Analyses
3. Results
3.1. Detection of TGase Activity and Effect of Ca2+ and GTP
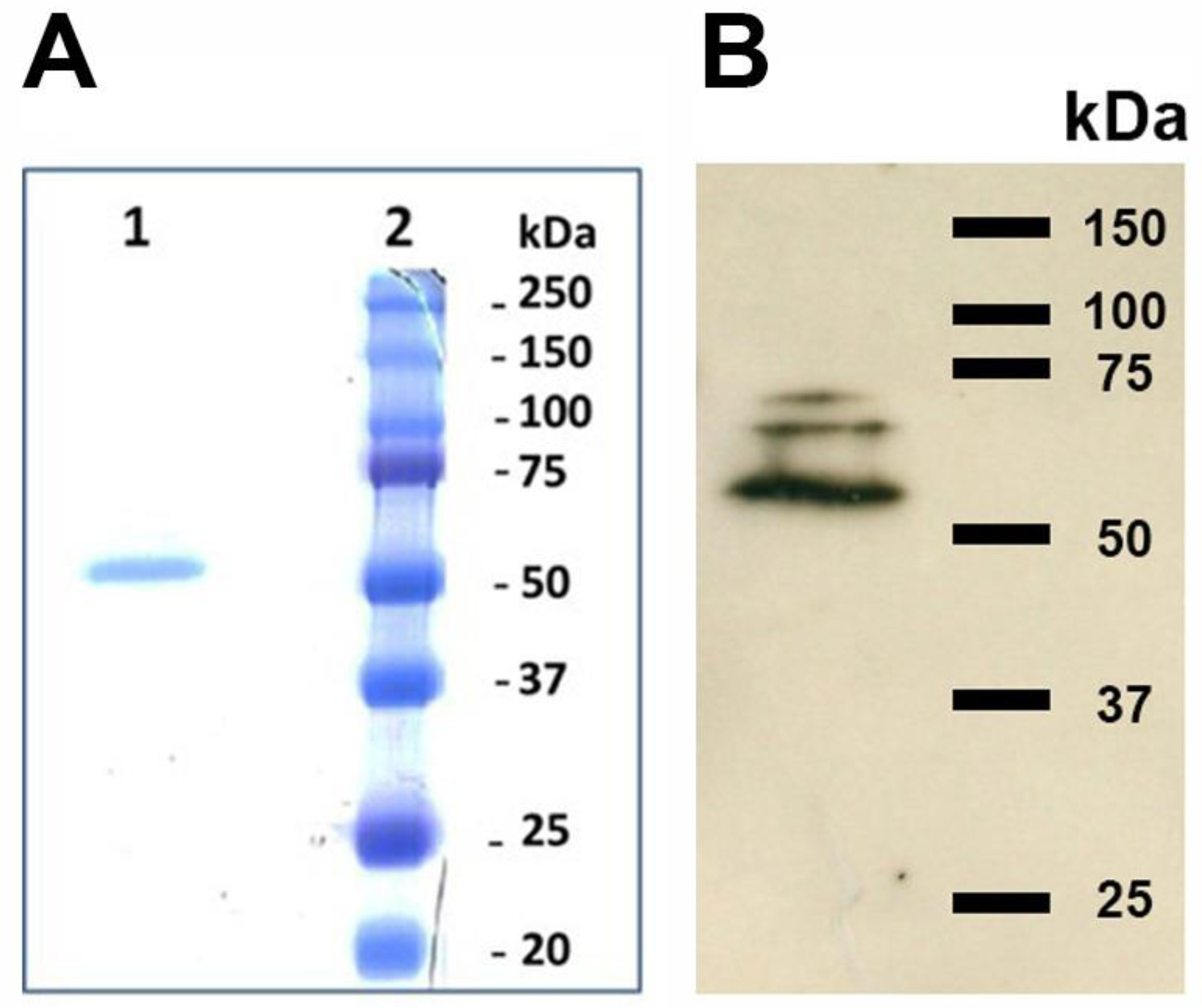
3.2. TGase Purification
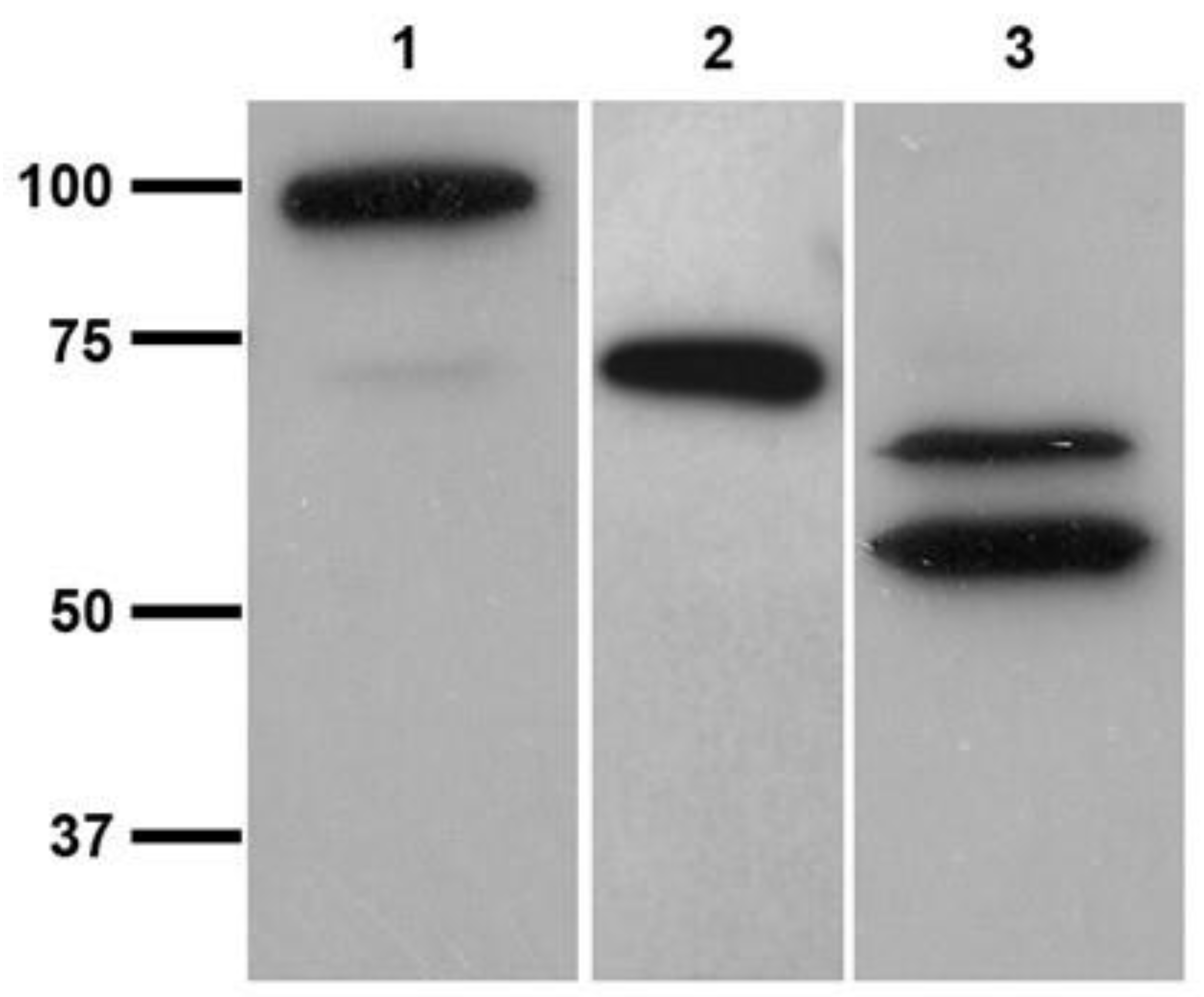
3.3. Cross-Reactivity of L. infantum Promastigotes with Antibodies against Human TGase 2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruschi, F.; Gradoni, L. The Leishmaniases: Old Neglected Tropical Diseases; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar]
- Knight, C.A.; Harris, D.R.; Alshammari, S.O.; Gugssa, A.; Young, T.; Lee, C.M. Recent epidemiological studies in the Middle East. Front. Microbiol. 2023, 13, 1052478. [Google Scholar] [CrossRef]
- Gossage, S.M.; Rogers, M.E.; Bates, P.A. Two separate growth phases during the development of Leishmania in sand flies: Implications for understanding the life cycle. Int. J. Parasitol. 2003, 33, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Yuste, M.; Martin-Sanchez, J.; Corpas-Lopez, V. Canine Leishmaniasis: Update on Epidemiology, Diagnosis, Treatment, and Prevention. Vet. Sci. 2022, 9, 387. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Das, P. Recent progress in drug targets and inhibitors towards combating leishmaniasis. Acta Trop. 2018, 181, 95–104. [Google Scholar] [CrossRef]
- Lorand, L.; Conrad, S.M. Transglutaminases. Mol. Cell. Biochem. 1984, 58, 9–35. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Romito, G.; Pepe, I.; De Vivo, G.; Merola, A.; Limatola, A.; Gentile, V. Transglutaminase-catalyzed reactions responsible for the pathogenesis of celiac disease and neurodegenerative diseases: From basic biochemistry to clinic. Curr. Med. Chem. 2006, 13, 1895–1902. [Google Scholar] [CrossRef]
- Eckert, R.L.; Kaartinen, M.T.; Nurminskaya, M.; Belkin, A.M.; Colak, G.; Johnson, G.V.W.; Mehta, K. Transglutaminase regulation of cell function. Physiol. Rev. 2014, 94, 383–417. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, G.; Suwa, M.; Ichikawa, Y.; Ohtsuka, T.; Kumagai, S.; Kikuchi, M.; Sato, Y.; Saito, Y. A novel function of tissue-type transglutaminase: Protein disulphide isomerase. Biochem. J. 2003, 373, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.U.; Mehta, K. Transglutaminases, thioredoxins and protein disulphide isomerase: Diverse enzymes with a common goal of cross-linking proteins in lower organisms. Indian J. Exp. Biol. 2004, 42, 235–243. [Google Scholar]
- Griffin, M.; Casadio, R.; Bergamini, C.M. Transglutaminases: Nature’s biological glues. Biochem. J. 2002, 368, 377–396. [Google Scholar] [CrossRef] [Green Version]
- Candi, E.; Paradisi, A.; Terrinoni, A.; Pietroni, V.; Oddi, S.; Cadot, B.; Jogini, V.; Meiyappan, M.; Clardy, J.; Finazzi-Agro, A.; et al. Transglutaminase 5 is regulated by guanine-adenine nucleotides. Biochem. J. 2004, 381, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Casadio, R.; Polverini, E.; Mariani, P.; Spinozzi, F.; Carsughi, F.; Fontana, A.; Polverino de Laureto, P.; Matteucci, G.; Bergamini, C.M. The structural basis for the regulation of tissue transglutaminase by calcium ions. Eur. J. Biochem. 1999, 262, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Cerione, R.A.; Clardy, J. Structural basis for the guanine nucleotide-binding activity of tissue transglutaminase and its regulation of transamidation activity. Proc. Natl. Acad. Sci. USA 2002, 99, 2743–2747. [Google Scholar] [CrossRef] [Green Version]
- Beninati, S.; Piacentini, M.; Bergamini, C.M. Transglutaminase 2, a double face enzyme. Amino Acids 2017, 49, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Adini, A.; Krugliak, M.; Ginsburg, H.; Li, L.; Lavie, L.; Warburg, A. Transglutaminase in Plasmodium parasites: Activity and putative role in oocysts and blood stages. Mol. Biochem. Parasitol. 2001, 117, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Duran, R.; Junqua, M.; Schmitter, J.M.; Gancet, C.; Goulas, P. Purification, characterisation, and gene cloning of transglutaminase from Streptoverticillium cinnamoneum CBS 683.68. Biochimie 1998, 80, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Strop, P. Versatility of Microbial Transglutaminase. Bioconjugate Chem. 2014, 25, 855–862. [Google Scholar] [CrossRef]
- Brobey, R.K.; Soong, L. Leishmania species: Evidence for transglutaminase activity and its role in parasite proliferation. Exp. Parasitol. 2006, 114, 94–102. [Google Scholar] [CrossRef]
- Atayde, V.D.; Hassani, K.; da Silva Lira Filho, A.; Raposo Borges, A.; Adhikari, A.; Martel, C.; Olivier, M. Leishmania exosomes and other virulence factors: Impact on innate immune response and macrophage functions. Cell. Immunol. 2016, 309, 7–18. [Google Scholar] [CrossRef]
- Gupta, A.K.; Das, S.; Kamran, M.; Ejazi, S.A.; Ali, N. The pathogenicity and virulence of Leishmania-interplay of virulence factors with host defenses. Virulence 2022, 13, 903–935. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.; Facchiano, A. Classification of microbial transglutaminases by evaluation of evolution trees, sequence motifs, secondary structure topology and conservation of potential catalytic residues. Biochem. Biophys. Res. Commun. 2019, 509, 506–513. [Google Scholar] [CrossRef] [PubMed]
- McCarthy-Burke, C.; Bates, P.A.; Dwyer, D.M. Leishmania donovani: Use of two different, commercially available, chemically defined media for the continuous in vitro cultivation of promastigotes. Exp. Parasitol. 1991, 73, 385–387. [Google Scholar] [CrossRef]
- Lajemi, M.; Demignot, S.; Borge, L.; Thenet-Gauci, S.; Adolphe, M. The use of Fluoresceincadaverine for detecting amine acceptor protein substrates accessible to active transglutaminase in living cells. Histochem. J. 1997, 29, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Lorand, L.; Parameswaran, K.N.; Velasco, P.T.; Hsu, L.K.; Siefring, G.E., Jr. New colored and fluorescent amine substrates for activated fibrin stabilizing factor (Factor XIIIa) and for transglutaminase. Anal. Biochem. 1983, 131, 419–425. [Google Scholar] [CrossRef]
- Lin, Y.; Means, G.E.; Feeney, R.E. The action of proteolytic enzymes on N,N-dimethyl proteins. Basis for a microassay for proteolytic enzymes. J. Biol. Chem. 1969, 244, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, H.F.; Conway, E.; Patterson, M.K., Jr.; Birckbichler, P.J.; Maxwell, M.D. Cellular transglutaminase has affinity for extracellular matrix. Vitro Cell. Dev. Biol. 1987, 23, 795–800. [Google Scholar] [CrossRef]
- Gambetti, S.; Dondi, A.; Cervellati, C.; Squerzanti, M.; Pansini, F.S.; Bergamini, C.M. Interaction with heparin protects tissue transglutaminase against inactivation by heating and by proteolysis. Biochimie 2005, 87, 551–555. [Google Scholar] [CrossRef]
- Wilhelm, B.; Meinhardt, A.; Seitz, J. Transglutaminases: Purification and activity assays. J. Chromatogr. B Biomed. Appl. 1996, 684, 163–177. [Google Scholar] [CrossRef]
- Nurminskaya, M.V.; Belkin, A.M. Cellular functions of tissue transglutaminase. Int. Rev. Cell. Mol. Biol. 2012, 294, 1–97. [Google Scholar]
- Parrotta, L.; Tanwar, U.K.; Aloisi, I.; Sobieszczuk-Nowicka, E.; Arasimowicz-Jelonek, M.; Del Duca, S. Plant Transglutaminases: New Insights in Biochemistry, Genetics, and Physiology. Cells 2022, 11, 1529. [Google Scholar] [CrossRef]
- Davids, B.J.; Mehta, K.; Fesus, L.; McCaffery, J.M.; Gillin, F.D. Dependence of Giardia lamblia encystation on novel transglutaminase activity. Mol. Biochem. Parasitol. 2004, 136, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Knodler, L.A.; Noiva, R.; Mehta, K.; McCaffery, J.M.; Aley, S.B.; Svard, S.G.; Nystul, T.G.; Reiner, D.S.; Silberman, J.D.; Gillin, F.D. Novel protein-disulfide isomerases from the early-diverging protist Giardia lamblia. J. Biol. Chem. 1999, 274, 29805–29811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, B.X.; Soong, L. Identification and enzymatic activities of four protein disulfide isomerase (PDI) isoforms of Leishmania amazonensis. Parasitol. Res. 2008, 102, 437–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.N.; Mehta, K. Purification and characterization of a novel transglutaminase from filarial nematode Brugia malayi. Eur. J. Biochem. 1994, 225, 625–634. [Google Scholar] [CrossRef]
- Jang, T.H.; Lee, D.S.; Choi, K.; Jeong, E.M.; Kim, I.G.; Kim, Y.W.; Chun, J.N.; Jeon, J.H.; Park, H.H. Crystal Structure of Transglutaminase 2 with GTP Complex and Amino Acid Sequence Evidence of Evolution of GTP Binding Site. PLoS ONE 2014, 9, e107005. [Google Scholar] [CrossRef] [Green Version]
- Kiraly, R.; Csosz, E.; Kurtan, T.; Antus, S.; Szigeti, K.; Simon-Vecsei, Z.; Korponay-Szabo, I.R.; Keresztessy, Z.; Fesus, L. Functional significance of five noncanonical Ca2+-binding sites of human transglutaminase 2 characterized by site-directed mutagenesis. FEBS J. 2009, 276, 7083–7096. [Google Scholar] [CrossRef] [Green Version]
- Antonyak, M.A.; Jansen, J.M.; Miller, A.M.; Ly, T.K.; Endo, M.; Cerione, R.A. Two isoforms of tissue transglutaminase mediate opposing cellular fates. Proc. Natl. Acad. Sci. USA 2006, 103, 18609–18614. [Google Scholar] [CrossRef] [Green Version]
- Fyfe, P.K.; Oza, S.L.; Fairlamb, A.H.; Hunter, W.N. Leishmania Trypanothione Synthetase-Amidase structure reveals a basis for regulation of conflicting synthetic and hydrolytic activities. J. Biol. Chem. 2008, 283, 17672–17680. [Google Scholar] [CrossRef] [Green Version]
- Anantharaman, V.; Aravind, L. Evolutionary history, structural features and biochemical diversity of the NlpC/P60 superfamily of enzymes. Genome Biol. 2003, 4, R11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Aravind, L. Novel transglutaminase-like peptidase and C2 domains elucidate the structure, biogenesis and evolution of the ciliary compartment. Cell Cycle 2012, 11, 3861–3875. [Google Scholar] [CrossRef] [Green Version]
- Atayde, V.D.; Aslan, H.; Townsend, S.; Hassani, K.; Kamhawi, S.; Olivier, M. Exosome secretion by the parasitic protozoan Leishmania within the sand fly midgut. Cell Rep. 2015, 13, 957–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denny, P.W.; Gokool, S.; Russell, D.G.; Field, M.C.; Smith, D.F. Acylation-dependent Protein Export in Leishmania. J. Biol. Chem. 2000, 275, 11017–11025. [Google Scholar] [CrossRef] [Green Version]
- Silverman, J.M.; Clos, J.; Camargo de’Oliveira, C.; Shirvani, O.; Fang, Y.; Wang, C.; Foster, L.J.; Reiner, N.E. An exosome-based secretion pathway is responsible for protein export from Leishmania and communication with macrophages. J. Immunol. 2010, 185, 5011–5022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Hidalgo, L.; Altuntas, S.; Rossin, F.; D’Eletto, M.; Marsella, C.; Farrace, M.G.; Falasca, L.; Antonioli, M.; Fimia, G.M.; Piacentini, M. Transglutaminase type 2-dependent selective recruitment of proteins into exosomes under stressful cellular conditions. Biochim. Biophys. Acta 2016, 1863, 2084–2092. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Hadano, J.; Kawasaki, D.; Dong, X.; Kawabata, S.I. Drosophila TG-A transglutaminase is secreted via an uncon ventional Golgi-independent mechanism involving exosomes and two types of fatty acylations. J. Biol. Chem. 2017, 292, 10723–10734. [Google Scholar] [CrossRef] [Green Version]
- Tatsukawa, H.; Hitomi, K. Role of Transglutaminase 2 in Cell Death, Survival, and Fibrosis. Cells 2021, 10, 1842. [Google Scholar] [CrossRef] [PubMed]
- Gannavaram, S.; Debrabant, A. Programmed cell death in Leishmania: Biochemical evidence and role in parasite infectivity. Front. Cell. Infect. Microbiol. 2012, 2, 95. [Google Scholar] [CrossRef] [Green Version]
- Mendes Wanderley, J.L.; Pinto da Silva, L.H.; Deolindo, P.; Soong, L.; Matos Borges, V.; Brito Prates, D.; Almeida de Souza, A.P.; Barral, A.; de Freitas Balanco, J.M.; Tanny Cunha do Nascimento, M.; et al. Cooper Cooperation between Apoptotic and Viable Metacyclics Enhances the Pathogenesis of Leishmaniasis. PLoS ONE 2009, 4, e5733. [Google Scholar]
- Medina-Colorado, A.A.; Osorio, E.Y.; Saldarriaga, O.A.; Travi, B.L.; Kong, F.; Spratt, H.; Soong, L.; Melby, P.C. Splenic CD4+ T Cells in Progressive Visceral Leishmaniasis show a mixed effector-regulatory phenotype and impair macrophage effector function through Inhibitory Receptor expression. PLoS ONE 2017, 12, e0169496. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almugadam, S.H.; Trentini, A.; Maritati, M.; Contini, C.; Manfrinato, M.C.; Cervellati, C.; Bellini, T.; Hanau, S. A Calcium- and GTP-Dependent Transglutaminase in Leishmania infantum. Vet. Sci. 2023, 10, 234. https://doi.org/10.3390/vetsci10030234
Almugadam SH, Trentini A, Maritati M, Contini C, Manfrinato MC, Cervellati C, Bellini T, Hanau S. A Calcium- and GTP-Dependent Transglutaminase in Leishmania infantum. Veterinary Sciences. 2023; 10(3):234. https://doi.org/10.3390/vetsci10030234
Chicago/Turabian StyleAlmugadam, Shawgi Hago, Alessandro Trentini, Martina Maritati, Carlo Contini, Maria Cristina Manfrinato, Carlo Cervellati, Tiziana Bellini, and Stefania Hanau. 2023. "A Calcium- and GTP-Dependent Transglutaminase in Leishmania infantum" Veterinary Sciences 10, no. 3: 234. https://doi.org/10.3390/vetsci10030234